











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
The Antillean manatee (Trichechus manatus manatus), one of 2 subspecies of West Indian manatees, is listed as endangered in the International Union for the Conservation of Nature (IUCN) Red List of Threatened Species (Self-Sullivan and Mignucci-Giannoni 2008). Little has been published on the presence of Antillean manatees in the Usumacinta and Grijalva Rivers (Mexico). In his monograph on the mammals of Yucatán, México, Gaumer (1917) described these 2 rivers and surrounding wetlands as sites with frequent manatee sightings. In 1976, the United States Fish and Wildlife Service carried out interviews that confirmed the frequency of manatees along the Usumacinta River (Campbell and Gicca 1978). Some years later, using data from interviews, Colmenero-Rolón (1984) found that the Grijalva-Usumacinta Basin in the state of Tabasco was the most important manatee habitat area in Mexico. This could be true, but the region has not been systematically studied yet (Álvarez-Flores et al. 1988, UNEP 2010, Jiménez-Domínguez and Olivera-Gómez 2014). The Regional Management Plan for the West Indian Manatee (Trichechus manatus) reported a minimum population size of 1,000- 2,000 manatees, based on best available data (UNEP 2010), and the majority of these individuals might be found in the Grijalva-Usumacinta hydrological basin in Tabasco (SEMARNAT/CONANP 2010). According to the most recent IUCN Red List assessment, best available data for Mexico indicate an estimated minimum population size of 1,000 individuals and an unknown population trend due to data deficiencies (Deutsch et al. 2008).
Manatees frequently inhabit fluvial-lagoon systems throughout the world, such as those in Venezuela and Colombia (Castelblanco-Martínez et al. 2009, Rivas-Rodríguez et al. 2012, Jaramillo-Villa 2015), Brazil (Arraut et al. 2010), and Mexico (Colmenero-Rolón and Hoz-Zavala 1986, Rodas-Trejo et al. 2008, Jiménez-Domínguez and Olivera-Gómez 2014). Turbid waters in these ecosystems have historically curtailed manatee studies (González-Socoloske and Olivera-Gómez 2012). However, a technique was developed to detect manatees in dark, turbid waters using side-scan sonar (SSS), and in studies conducted in Florida and Mexico it yielded 81% and 93% accurate detection rates, respectively (González-Socoloske et al. 2009). With this technique, we can determine important population characteristics in turbid waters, such as group size and mother-calf pairs (based on the relative size of the individuals) (González-Socoloske and Olivera-Gómez 2012).
The Usumacinta River has many small rivers systems with interconnecting streams and lagoons where manatees have been frequently sighted. One of these systems, the San Pedro River system, is known for the high frequency of manatee sightings in it (Jiménez-Domínguez and Olivera-Gómez 2014). This study aimed to estimate the population density of manatees in the San Pedro River and associated streams using SSS, and to determine how this density varies with changes in water level.
Materials and methods
Study area
The San Pedro River system is located in the southern portion of the municipality of Balancán (18º10 -17º26 N, 90º59 -91º42 W) (Fig. 1a). It extends from the Reforma rapids and Reforma Waterfalls in the Limón River to the terminus of the San Pedro River, which empties into the Usumacinta River (Fig. 1a, b). The San Pedro River system is a calcareous subbasin that contains water with a weak load of suspended material (Castillo -Domínguez et al. 2011), a normal condition of the system down to the Limón River. The last section of the system extends ~6.5 km upstream from the San Pedro River mouth and includes the Susil and Balmoc streams (Fig. 1a). This section is within the alluvial terraces of the Usumacinta River (West et al. 1985) and is strongly affected by the heavy loads of suspended particles coming from this river. There are 2 lagoons in this section that were not included in the study. The Limón River, which is an upstream continuation of the San Pedro River, was partially included in some surveys (Fig 1b).
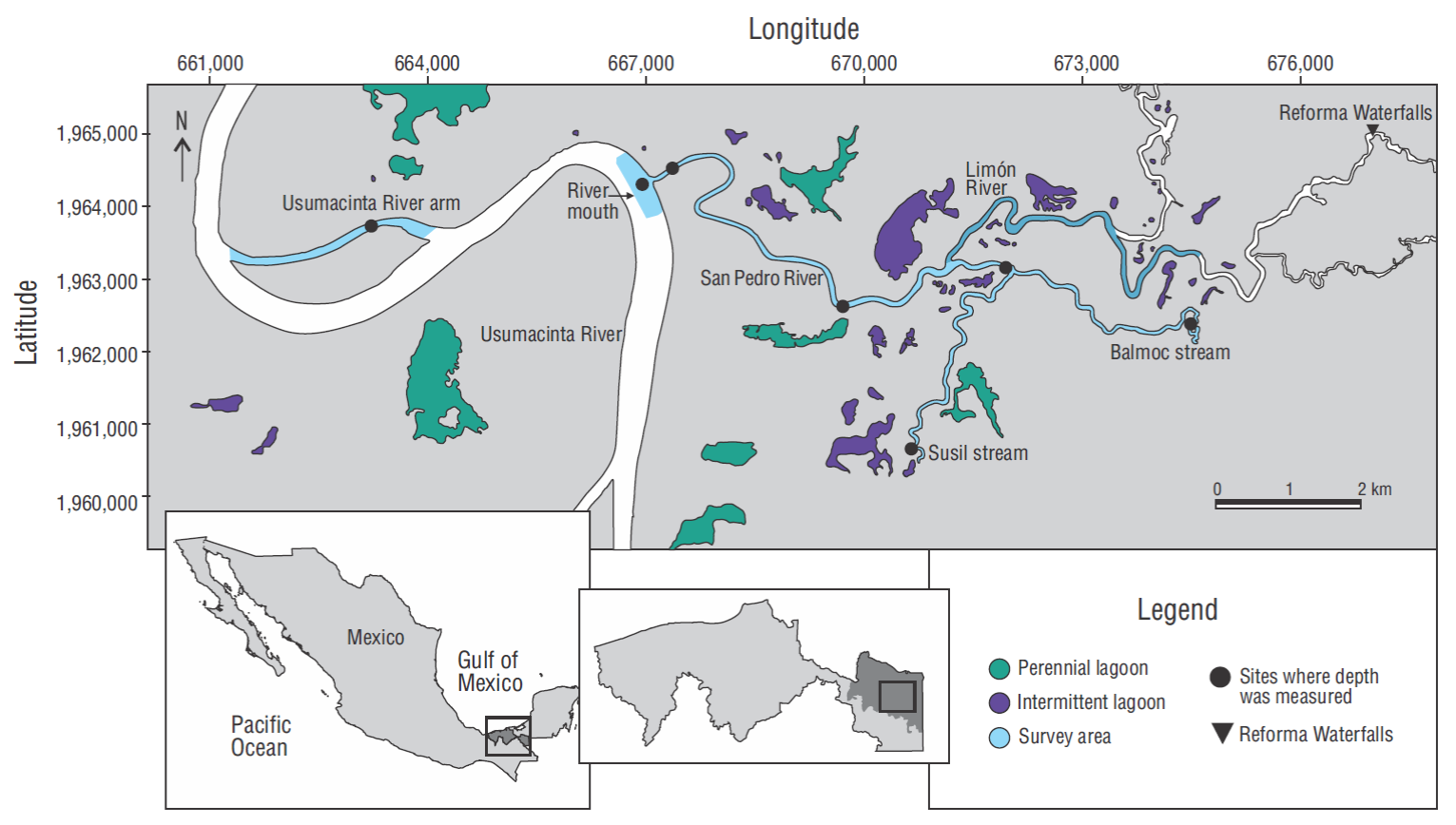
Figure 1 Map showing the southern portion of the Usumacinta River, and the San Pedro River system. The shaded river segments were surveyed using side-scan sonar. The darker shaded segment indicates the segment of the Limón River that was sampled during the last 4 sessions. Filled circles indicate the sites where depth was measured and the filled triangle indicates the location of Reforma Waterfalls.
The San Pedro River is fed by the Usumacinta River and is thus subject to major changes in water level. During the flood season (September-October), rivers overflow large areas, interconnecting all water bodies in surrounding wet-lands and grasslands (González -Socoloske 2013, Jiménez-Domínguez and Olivera-Gómez 2014). Conversely, during the dry season (March to June), rivers reach low water levels, exposing riverine walls 6 m high and/or blocking access to large bodies of water. When water level is rapidly increasing (July-August), conditions are locally known as “rezumo”, or rising level, and when the water level is falling, conditions are known as falling levels (November-February). This changing water level has a direct effect on food availability for manatees, since richness and abundance of plant species decrease with low water levels (González -Socoloske 2013).
Vegetation cover and composition was homogeneous in the survey areas along the San Pedro River, nearby streams, and the first 5 km of the Limón River, and it was mainly composed of Dalbergia glabra, Parmentiera parviflora, Lonchocarpus guatemalensis, and Mimosa pigra. There is a change in vegetation upstream of the Limón River, near the Reforma Waterfalls, where mangrove grows (Rhizophora mangle). The effects of the rising and falling water levels are observed in all water bodies.
Near the San Pedro River and the Susil and Balmoc streams there are permanent lagoons, and intermittent lagoons form during high water events. Lagoons near the Limón River are scarce because of the permeable, calcareous soil (Fig. 1a).
Fishing is one of the local economic activities that take place in the study area. To fish, villagers place large semipermanent nylon nets, which they call “tapes,” across the full width of a water course to trap fish as they swim by it. This fishing equipment is abandoned or removed during flood season, and it is the main cause of manatee deaths in some areas of this region (Romero-Berny et al. 2014). In the San Pedro River system, manatees are learning to lift the base of the tapes (Morales-Vela 2016). In the mid-1980s there were sporadic cases of manatee hunting in some areas of Tabasco (Colmenero-Rolón and Hoz-Zavala 1986); however, there were no confirmed reports of hunting in the past decade. Fishermen in the San Pedro River system stated that manatees are not hunted in the area, and there were no hunting reports during the study period. People that were interviewed in areas bordering rivers know that manatees are protected by federal laws. They also have some knowledge about this species, its ecological importance, and the areas it inhabits (Guzmán-Nieto 2008).
Sampling scheme
Surveys were conducted on a 5-m-long fiberglass boat powered by a 15-HP outboard engine, at a maximum speed of 10 km/h. We used a Humminbird 899ci HD SI Combo system (Johnson Outdoors, Eufaula, AL, USA; 455 kHz frequency, 180º water column coverage) to detect manatees and estimate manatee density (González-Socoloske et al. 2009). The SSS system was calibrated to cover 25 m on each side of the boat to maintain clear manatee distinction across a 50-m bandwidth (Fig. 2). The crew included a local guide with experience in manatee observations and detection of their traces; this person did visual scanning and controlled the boat. There was also an observer with experience in manatee detection who monitored the SSS and took notes. Both crew-members performed the same activity during the entire study.

Figure 2 Side-scan sonar image showing 3 adult manatees recorded at the San Pedro River; 2 manatees are close together.
All surveys took place from January to June 2016, over a course of 2 non-consecutive weeks of field work (i.e., 2 sessions) per month. Each session consisted of 3 consecutive sampling days, running one SSS boat survey per day, for a total of 6 surveys per month. Boat surveys were conducted from 08:00 to 11:00 local time, but sometimes this time period was extended to 13:00 because of the number of detected manatees and the additional time needed to collect all complementary information. In all, we implemented 36 sampling surveys using the SSS technique over 12 sessions.
To observe possible changes in manatee detection by SSS between hours, 2 additional partial survey rounds were conducted during the last 5 sessions (April-June), one from 13:00 to 15:00 and the other from 18:00 to 20:00. These 2 surveys covered the segment from the San Pedro River entrance to the Limón River confluence. This was also done to observe changes in manatee preferences for the central (from the transect line to 12 m on each side) or lateral (13 to 25 m from the transect line) parts of the river.
Water depth was recorded at 5 fixed points throughout the system and at 2 fixed points in the Usumacinta River to compare changes in water depth throughout the study period (Fig. 1). These points were chosen because they were easy to recognize in the field (i.e., there was something characteristic at each point, for example a tree trunk) and we had geographic coordinates for those points.
Coverage area
To obtain transect lengths, during a prospective visit UTM coordinates were obtained for several points in the study area using a GPS (Garmin eTrex 30). These geographic locations were used in Google Earth, and distance between geographic coordinates was measured with the Route tool. To calculate the square area, we used the layer from the National Institute of Statistics and Geography (INEGI, Mexico) Hydrological Network in ArcGIS 10.3 (.shp file; scale = 1:50,000).
The study area covered the first 6.5 km upstream of the San Pedro River (area = 0.45 km2), the Susil and Balmoc streams (3.5 and 4.0 km long, respectively; both comprising an area of 0.19 km2), the San Pedro River mouth (500 × 200 m; 0.17 km2), and an arm of shallow water (area 0.42 km2) that is 3.3 km away from the San Pedro River mouth (Fig. 1a). Within the San Pedro River system, there are 2 lagoons connected to the Susil and Balmoc streams (both lagoons go by the same name of the streams) that were not included in the study. The Limón River was surveyed along 5 km (0.44 km2) from its confluence with the San Pedro River and only during the May and June sessions (9-12) (Fig. 1b). We followed a zigzag pattern at the San Pedro River mouth and a linear pattern at the arm. The streams were not surveyed during sessions 8 (April), 9, and 10 (May), nor was the Usumacinta arm during sessions 8-12 (April-June) because low water levels hindered access. The total sampling area was 1.67 km2 (Fig. 1).
Positive confirmation of manatees
There are 2 errors associated with the observer using the SSS to detect manatees: false positive, when another object is mistaken for a manatee, and false negative, when a manatee is detected by the sonar unit but goes undetected by the observer (González-Socoloske y Olivera-Gómez 2012). These errors are called perception biases (Reynolds et al. 2012). Thus, to minimize both errors, when a manatee or a possible sighting was detected, an image was recorded by the sonar unit and we passed over the same location a couple of times as a preliminary procedure for confirmation. A positive record on the sonar unit display should have met the following criteria: shape, size, shadow projection, and acoustic reflection (Fig. 2) (González-Socoloske et al. 2009). Simultaneous visual scanning was done to directly find a manatee or to look for indirect evidence of its presence, such as bubbles on the water surface, suspended sediment in the water column, floating feces, or chewed plants. All recorded SSS images were examined using the above criteria. As a second measure to minimize human error, we made short stops to rest and eat. Also, an umbrella was used to protect the sonar display from the sun, improving detection and reducing glare.
Statistical analysis
Density was expressed as the number of detected manatees per square kilometer. To calculate manatee density at the San Pedro and Limón Rivers, the system was divided into 2 sections (Fig. 1b). The first section included the survey areas at the San Pedro River and its streams, for a total of 0.64 km2. The second section was 0.44 km2 and included the Limón River. Total area for the 2 sections was 1.08 km2. Sectioning of areas was done because the Limón River was surveyed during only 4 sessions and because its physical characteristics are different from those of the San Pedro River and its streams. The Usumacinta River arm and the San Pedro River mouth were not taken into account because manatees were not detected during the SSS surveys.
To identify differences in mean density among survey sessions, we used the Friedman test (α = 0.05) with an a posteriori mean range comparative analysis. This test works with related (dependent) samples and compares population means. The test allows for the analysis of repeated measures data if participants are assessed on 2 or more occasions or under 2 or more conditions. The null hypothesis indicates no differences between datasets and the alternative hypothesis indicates that there are differences between groups (Siegel and Castellan 1995). The test was chosen because of the dependency of the data, as sampling was done over the same area and observations possibly belonged to the same set of animals seen at different times.
Confidence intervals for manatee density in the San Pedro River system were calculated using the bootstrap method (1,000 samples) (Efron 1979, Good 2006), taking the population mean as a parameter. Estimates were computed using Infostat statistical software (Di Rienzo et al. 2016).
Total number of animals in the system
To determine how many manatees were in the system, 2 strata were defined: the San Pedro River (including Susil and Balmok streams) and the Limón River survey areas (during sessions 9-12). We estimated the number of manatees by using mean and maximum density (Table 1). As a possible alternative scenario, we extrapolated density at the Limón River sampled area to the non-sampled area (0.29 km2) upriver to the Reforma Waterfalls, which act as natural barriers for manatee dispersion (Fig. 1b).
Table 1 Mean manatee density (no. of manatees per square kilometer) in the San Pedro River system from January through June 2016. Values in parentheses indicate the estimated mean density for the Limón River.
| Month | Session | Density | Standard deviation |
| January | 1* | 3.7 | 1.8 |
| 2* | 9.9 | 4.8 | |
| February | 3† | 12.0 | 3.3 |
| 4*+ | 21.9 | 3.1 | |
| March | 5* | 26.6 | 1.6 |
| 6* | 22.5 | 7.4 | |
| April | 7* | 12.0 | 0.9 |
| 8 | 14.9 | 3.4 | |
| May | 9* | 25.4 (3.0) | 0.7 |
| 10* | 32.9 (5.3) | 1.3 | |
| June | 11* | 43.3 (0.8) | 3.8 |
| 12* | 35.0 (0.0) | 7.1 | |
| Overall | 21.7 | 3.2 |
Results
A total of 471 manatees were counted at the San Pedro River system, 3 of which were found near the mouth of the system. No manatees were observed at the Usumacinta River arm. Only 12 manatees were found in the Limón River sampled area. From the total count, 450 (95%) manatees were positively confirmed during the first sonar scan and 21 (5%) were confirmed by the second or third pass.
Calves represent ~ 10% (n = 46) of the total count. Four different calves were identified: one was found dead in February (during session 3) and the other 3 were seen in company of an adult female on the same day at different sites. One of these calves was repeatedly assisted by its mother for swimming, so it was considered a newborn. The dead calf was a 110-cm-long male. Its state of decomposition indicated it may have been dead for a couple of days, but it was not possible to determine the cause of death. The presence of female-calf pairs was regularly observed within the San Pedro River system throughout the study (Table 1).
Density
Manatee Density in the San Pedro River and the 2 streams increased from January to June, with an average density of 21. 7 manatees/km2 (Table 1). Minimum average density was 3.7 manatees/km2 in mid-January (Table 1, Fig. 3). By the end of January, density continuously increased until it reached 26.6 manatees/km2 in early March, after which it decreased rapidly until the first week of April (session 7), when water level dropped (Fig. 3). Density then increased at a constant speed (session 9), reaching an average maximum of 43.3 manatees/km2 in mid-June (session 11), when water level increased (Fig. 3). A global analysis of density variation between sessions showed significant differences (Friedman χ2 = 40.719, df = 11, P < 0.01). According to the a posterior mean range analysis, similarities were observed between sessions 1-3, 7-8, 4-6 and 9, and 10-12, with significant differences among these groups. The average estimated density for the Limón River sampled area (sessions 9-12) was 2.3 manatees/km2, with a maximum of 5.3 manatees/km2 during session 10 and a minimum of 0.0 manatees/km2 during session 12 (Table 1, Fig. 3).
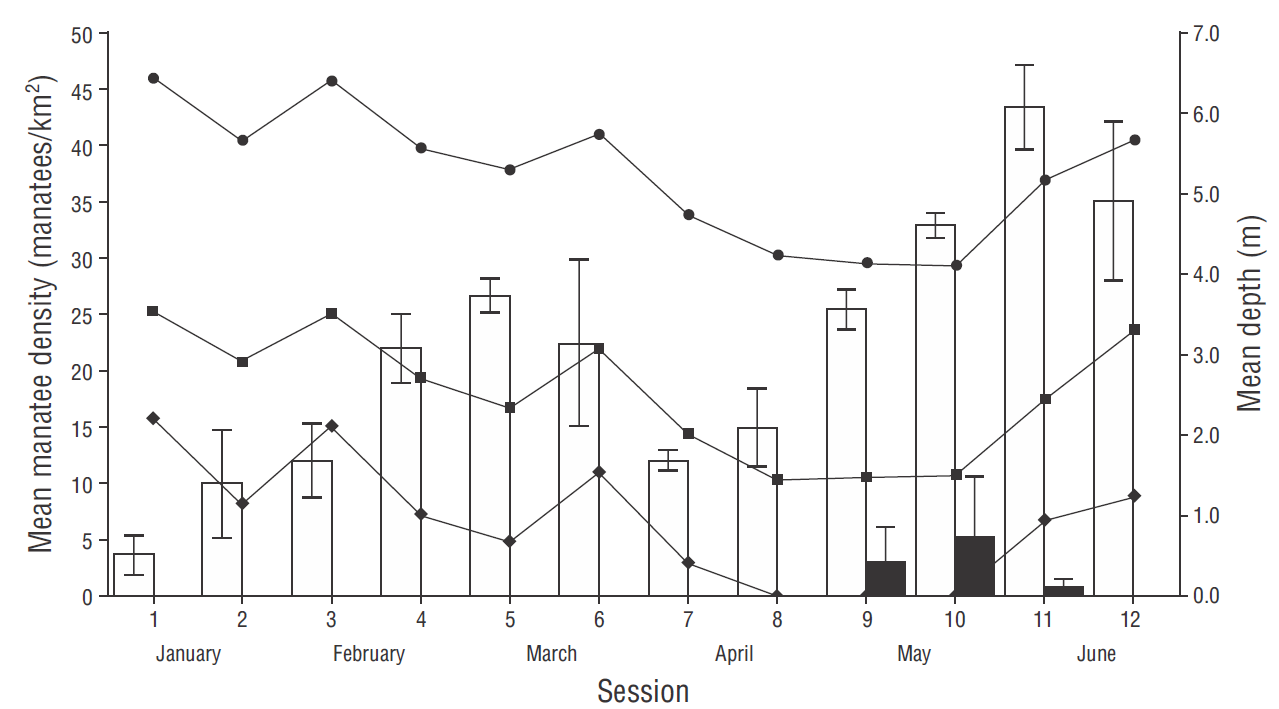
Figure 3 Graph showing the fluctuations of manatee density and mean water depth in the San Pedro River system from January to June 2016. Bars show mean manatee density in the San Pedro River and Balmoc and Susil streams (white), and in the Limón River (black); vertical lines on bars indicate standard deviation. Lines with symbols indicate mean depth at a fixed point in the San Pedro River (circles), at the entrance of the streams (squares), and in the Balmoc stream (diamonds).
Number of manatees in the system
The maximum number of manatees estimated for the San Pedro River system (1.08 km2) was 28 and occurred in June (session 11) (Table 2). This number is the sum of the 27.7 manatees estimated for the San Pedro River and the Susil and Balmoc streams (considering 43.3 manatees/km2 in 0.64 km2) and the 0.3 manatees estimated for the Limón River (considering 0.8 manatees in 0.44 km2, per data from session 11).
Table 2 Mean manatee density in the San Pedro River and the Susil and Balmoc streams, and in the sampled and unsampled segments of the Limón River. Maximum density corresponding to session 11 was used to obtain the number of manatees that may have been present in the San Pedro River system.
| Area (km2) | Mean density (manatees/km2) | Estimated N | Peak density for session 11 | Estimated N | |
| San Pedro River and streams | 0.64 | 21.7 | 13.8 | 43.3 | 27.7 |
| Limón River (sampled area) | 0.44 | 2.3 | 1.0 | 0.8 | 0.3 |
| Limón River (un-sampled area) | 0.29 | 2.3* | 0.7 | 0.8 | 0.2 |
*Using mean density for sampled area
When we extrapolated the density from the Limón River sampled area to the non-sampled area (total area = 0.73 km2), the estimated number of manatees in the Limón River was 5 using mean density and 1 using the density estimated from the session 11 data (Table 2). Under this scenario, the number of manatees in the system was 33 (27.7 plus 5) or 29 (27.7 plus 1), but since 2 lagoons in the system were not surveyed, the number of manatees could be somewhat larger. Under an optimal scenario, combining the peak densities from the San Pedro and Limón Rivers, the expected number of manatees could be nearly 59 animals, but this is a nonrealistic scenario.
Manatee availability between survey hours
The number of recorded manatees did not vary among the 3 different time intervals (Friedman χ2 = 0.5, d.f. = 2, P = 0.8145), indicating that the survey hours were unbiased. Manatees preferred the central part of the river in the morning (8:00-10:00) and afternoon (13:00-15:00) but tended to be near the river banks at twilight (18:00-20:00).
Discussion
Lack of manatee sightings at the Usumacinta River arm and the 3 manatees counted at the San Pedro River mouth indicate manatee preference for the San Pedro River system over the Usumacinta River. The San Pedro River system is an ideal site for females with calves because these individuals remained in that system throughout the study. We even observed helping behavior from a mother as she assisted her calf to reach the water surface and breathe; this usually occurs immediately after birth (Hartman 1979). This mother and calf pair remained at the same location during 3 consecutive days. The area where the mother-calf pairs were repeatedly sighted (Balmoc stream, near the Susil stream confluence) had very weak water flow (2 m/min), available vegetation, and little boat traffic (Fig. 1b). In Florida and the lower Usumacinta River basin, mothers with calves prefer to be in calm waters with food availability (Gannon et al. 2007, Jiménez-Domínguez y Olivera-Gómez 2014), which is in agreement with our observations.
The percentage of calves during the present study (10%) is slightly higher than that reported for Belize and Chetumal Bay (Mexico) (7.4%), a region with one of the largest manatee populations in the Caribbean (Morales-Vela et al. 2000). This highlights the reproductive importance of the San Pedro River system for manatees.
Variation in manatee density between sessions appears to be related to the movement of animals throughout the San Pedro River system and surrounding areas during the rising and falling of the water level (Fig. 4). Manatees used 3 routes to enter and exit the system: the mouth of the San Pedro River, the Limón River, and the Susil and Balmoc lagoons. The low manatee density observed in January and the ensuing constant increase through March can be explained as follows. In January, water level was high and manatees could have scattered throughout the flooded areas (including the 2 un-surveyed lagoons) instead of moving to the Usumacinta River. In fact, in January, only one manatee was recorded in the first section of the San Pedro River system and none were observed at the mouth. When water level was falling, manatees gathered in the San Pedro and Limón Rivers and the 2 streams because flooded areas were reduced and depth of perennial lagoons decreased. The low number of manatees in April and May is explained by the drastic reduction of accessible areas within the system. During this time manatees could have redistributed to the Limón River, areas outside the system, and areas nearby the Usumacinta River. However, we do not have records for these areas since we did not survey the Limón River during this time and coverage of the Usumacinta River was limited. The increased density values observed in June may be explained by the rising water level, when access to streams and lagoons was regained. Thus, changes in density are related to the rising and falling of the water lever in the lagoon system, supporting the assertion that manatees respond to flood pulses and desiccation periods (Fig. 4).

Figure 4 Plot of mean (gray dots) and smoothed mean (gray line) water depth, and of mean (Black dots) and smoothed mean (black line) manatee density.
We did not observe the possible evasive behavior of manatees in the presence of boats mentioned by González-Socoloske and Olivera-Gómez (2012). These authors indicated that manatees seem to avoid boats by swimming towards the river banks or to the deepest parts of the channel. During our study, manatees remained in almost the same positions after a couple of sampling rounds over them. The fact that manatees tended to stay in the central part of the river during the surveys reduced the error of absence, that is, that manatees were outside sonar detection range during the survey. A change in site preferences was evident at twilight, when manatees tended to remain near the river banks, but in the morning (8:00-10:00) and afternoon (13:00-15:00) they stayed in the central part of the water course. This change in site preferences may be related to feeding habits, since local fishermen have reported manatees feeding on the San Pedro River shore vegetation at sunrise. Some authors have reported that manatees move to feeding areas at sunrise, twilight, and nighttime (Powell 1996, Jiménez 2002).
The present study is the first to estimate manatee density in the San Pedro River System during flood pulses, and we were able to determine maximum and mean density in a key area. The Usumacinta Basin is one of the most important habitat regions for manatees in Mexico (Colmenero-Rolón 1984, SEMARNAT/CONANP 2010) . Considering that the San Pedro River system is one of the most important habitat areas for manatees in that basin (Jiménez-Domínguez and Olivera -Gómez 2014, Puc-Carrasco et al. 2016) and that population size in it is ~28 (or at most 59 using unrealistic scenarios), the current number of manatees in the Usumacinta Basin might be extremely lower than previously estimated. Our results indicate that earlier best guesses overestimated the number of manatees in the Usumacinta Basin. Other recent data increasing this concern include densities obtained from 35 SSS surveys at 6 selected sites on the coastal plain of Tabasco and in the San Pedro River, which indicate maximum mean abundance of 5.2 manatees/10 km (Puc-Carrasco et al. 2016), suggesting 66 to 93 manatees at these sites. Other regional sites where manatee abundance is probably significant are the Bitzales River (Puc-Carrasco et al. 2016); the Chico-Catazajá River system, shared by Tabasco and northern Chiapas (Rodas et al. 2008, Romero-Berny et al. 2014); and the Chacamax River and nearby Pochote fluvio-lagoon system (Álvarez-Flores et al. 1988) located 68.5 and 73.0 km from the San Pedro River, respectively. These areas, especially the last 2, need to be studied because there are no field-based estimates of manatee abundance for them.
The largest population of manatees on the Caribbean coast of Mexico is located in Chetumal Bay, off the southern part of the state of Quintana Roo (Morales-Vela and Olivera-Gómez 1997); Morales-Vela and Olivera-Gómez (1994) reported a relative abundance index of 12.3 manatees per flight hour and an estimated number of 90 animals (range 51-130) for this population. Manatee abundance is very low along the northern and western coasts of the Yucatán Peninsula (Mexico), as confirmed by the only manatee sighted after 17 h of aerial surveys (Morales-Vela et al. 2003). There are other areas in Mexico where manatee abundance is high, but Chetumal Bay and the Usumacinta River basin are some of the most important. The low manatee density estimated for the San Pedro River system signals an urgent need to determine the current population status of these animals in Mexico.
Belize and Mexico are reportedly the only 2 countries with populations of several hundred Antillean manatees (Deutsch et al. 2008). In Belize, great efforts have been made to determine manatee abundance via aerial surveys (O’Shea and Salisbury 1991, Morales-Vela et al. 2000, Auil 2004), and currently the best count from nationwide surveys is 507 (Auil-Gomez 2014).
All field-based studies contrast the expected 1,500 (1,000-2,000) manatees reported for Mexico and the 1,000 manatees reported for Belize (UNEP 2010). The minimum estimated population sizes for Mexico (1,000) and Belize (700), which are the current estimates reported in the IUCN Red List of Threatened Species (Deutsch et al. 2008), could be overestimated. These estimates appear to be approximate maximum rather than minimum population sizes.
More efforts should be made to estimate manatee population density in different parts of the world with murky waters and where manatees have been reported. The use of sonar is a valuable technique to obtain a more precise estimation of actual manatee population sizes, as shown by the present study.

