











 text new page (beta)
text new page (beta) English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroduction
The California Current System (CCS) is one of the most productive ecosystems in the world (Carr 2006). The southern portion of the CCS off the coast of Baja California (Mexico) is a transition zone with a clear seasonal signal. During winter and spring it is mainly influenced by subarctic waters, and during summer and autumn it is influenced by waters from the tropical Pacific off the coast of Mexico (Durazo and Baumgartner 2002, Durazo 2015). Off Baja California, the CCS is characterized by upwelling favorable conditions (Linacre et al. 2010), the formation of mesoscale eddies (Espinosa-Carreón et al. 2012), and the influence of interannual (El Niño and La Niña) and decadal events. Altogether, these factors control primary productivity (PP) and thus productivity in the pelagic ecosystem (Espinosa-Carreón et al. 2004).
Since 1997, the Investigaciones Mexicanas de la Corriente de California program (Mexican Research Program of the California Current; IMECOCAL, for its acronym in Spanish) has conducted in situ measurements of phytoplankton PP. These measurements have allowed us to identify temporal variations in PP at seasonal and interannual scales (Gaxiola-Castro et al. 2010), as well as significant differences in variations of integrated PP between summer and autumn (Aguirre- Hernández et al. 2004). Moreover, evidence indicates that the decrease in biomass and PP in the region is a consequence of interannual variations, such as the one associated with the anomalous intrusion of subarctic water (Espinosa-Carreón et al. 2015).
The response of primary producers to changes in oceanographic conditions, in terms of biomass, which is estimated using the concentration of chlorophyll (Chla), off the coast of Baja California has been documented based on both measurement data (Gaxiola-Castro et al. 2008, 2010) and remote sensing data (Espinosa-Carreón et al. 2004). On a seasonal scale, the southern portion of the CCS typically shows high Chla concentrations during spring due to the intensification of coastal upwelling, but these concentrations decrease during summer due to the combined effect of the weakening of upwelling favorable winds and the increase in water stratification. At the interannual scale, Chla concentrations show negative anomalies during warm periods associated with the sinking of the pycnocline, and during cold periods, when the pycnocline rises, phytoplankton biomass generally shows positive anomalies (Gaxiola-Castro et al. 2010).
One way to measure oceanic productivity is by using the optimal rate of carbon fixation normalized per unit of Chla (
Materials and methods
Data analyzed in the present study were collected during 47 oceanographic campaigns. These campaigns were carried out between 1997 and 2012 in the southern region of the CCS, off the Baja California Peninsula. In each oceanographic campaign, in situ experiments were carried out in order to determine PP in the euphotic zone (Fig. 1). The 14C fixation rate was measured with the light-dark bottle method through experiments that were carried out at around noon (local time) . Water samples were taken at optical depths corresponding to 100%, 50%, 30%, 20%, 10%, and 1% of surface irradiance. Physical depths were calculated by using the Beer law: Z = ln(Eo/Ez)/K d, where Z is depth, Eo is surfce irradiance, Ez is irradiance at depth Z, and K d is the mean diffuse attenuation coefficient, which was estimated by using depth measurements from the Secchi disc (Z d) and applying the ratio for oceanic waters K d =1.7/ Z d. Water samples were collected with 5-L Niskin bottles mounted on a General Oceanics rosette. To minimize sample contamination, silicone gaskets and elastics were used for the bottles. Conductivity, temperature, and pressure were measured with a factory calibrated CTD SeaBird 911plus at each station.
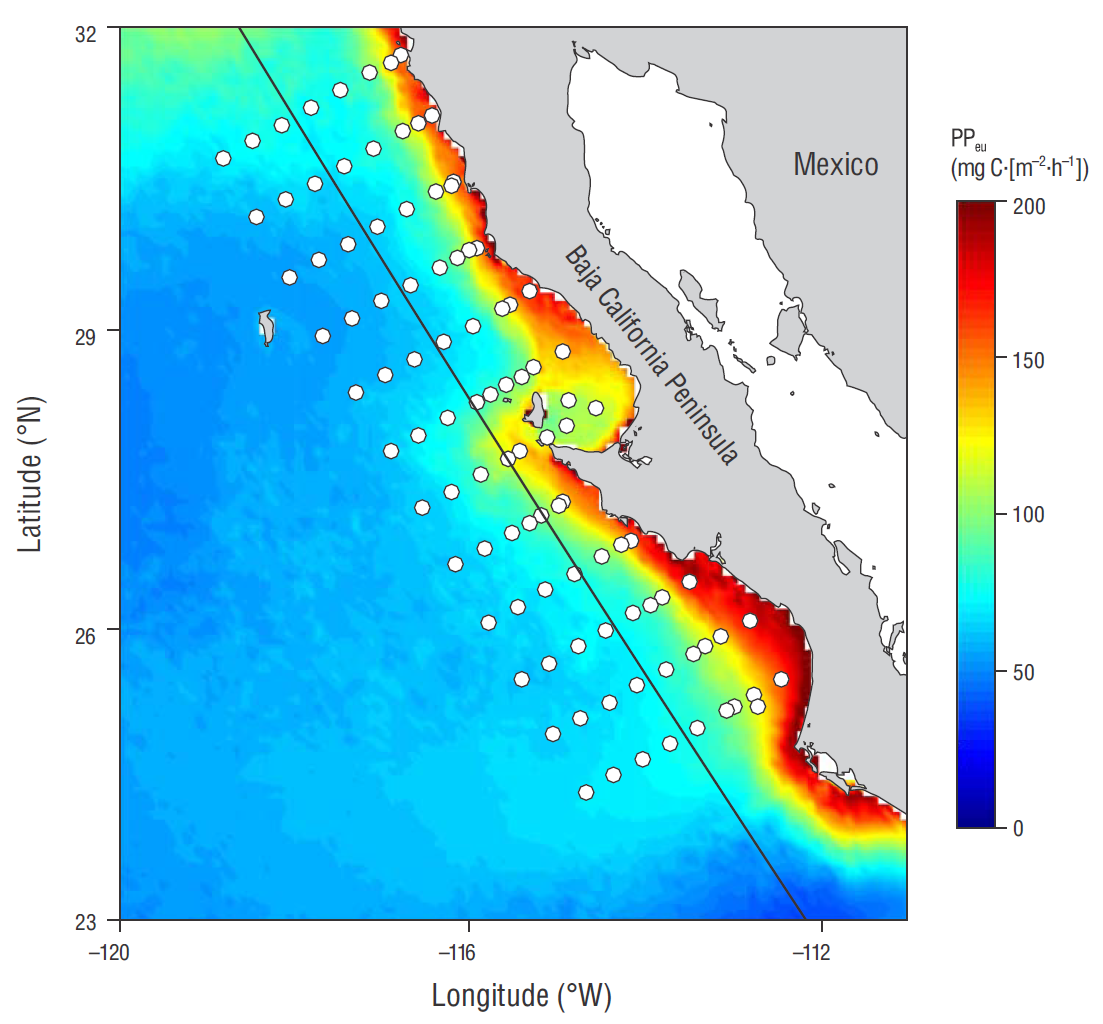
Figure 1 IMECOCAL monitoring network (white circles) . Circles indicate the stations where in situ experiments were carried out on at least one occasion to determine primary productivity (PP). The color scale shows integrated PP in the euphotic zone (PPeu) as calculated with the Vertically Generalized Production Model (Behrenfeld and Falkowski 1997); map shows the mean value calculated using the monthly data measured by MODIS-Aqua from 2002 to 2016 (https://oceancolor.gsfc.nasa.gov/). The transect located parallel to the coast (black line) was used to generate Figure 5.
Water samples collected at each depth were filtered with a 150-μm mesh to remove macrozooplankton and placed in 250-mL polycarbonate bottles, which were inoculated with 5 μCi of NaH14CO3. For each optical depth, 1 dark and 2 light bottles were used. They were placed in transparent acrylic tubes. Bottle arrangements were then returned to the sam-pling depth, and they were incubated for 1.5 to 2.0 h. After incubation, samples were filtered through a 45- μm GN-6 membrane. Each filter was placed in 20-mL glass vials, and 0.5 mL of 10% HCl were added to remove NaH14CO3 excess. After 3 h, 10 mL of scintillation cocktail (Ecolite) were added to each vial. Radioactivity was determined with a Beckman LS-6500 scintillation counter. PP estimations were calculated using disintegrations per minute and corrected by the dark bottle (Parsons et al. 1984). PP profiles were normalized using the Chla concentration, and
To quantify the Chla concentration (mg m-3) at each optical depth, 1 L of water was collected and filtered through GF/F Whatman glass fiber filters at a positive pressure. Filters were then placed in HistoPrep tissue capsules, properly labeled, and stored in liquid nitrogen for later analysis in the laboratory. At the laboratory, the Chla sample was placed in 10 mL of 90% acetone for 24 h at ~4 ºC in complete darkness (Venrick and Hayward 1984). Pigment concentration was quantified using the fluorometric method (Yentsch and Menzel 1963, Holm-Hansen 1965) and calibrating the fluorometer with a pattern of Chla (Sigma).
To determine phytoplankton composition, water samples were collected during the oceanographic cruises in the 2001-2008 period. Samples were collected at a depth of 10 m and stored in 250-mL dark bottles with formaldehyde at a pH of 7.5 to 8.0. Phytoplankton analysis was carried out within the first 2 months after each cruise. To perform cell counts, 50 mL of seawater were concentrated in a sedimentation chamber. Phytoplankton counting and identification were done using an inverted microscope with 16× and 40× lenses (ZEISS VERT. A1).
SST data, measured with the AVHRR-Pathfinder sensor and processed with version 5 of the algorithm, were obtained from https://podaac.jpl.nasa.gov/. The SST data for the 1985-2009 period correspond to global images of level 3 monthly composites, which correspond to monthly average measurements adjusted to a regular georeferenced grid with a spatial resolution of 4 km.
Results
The
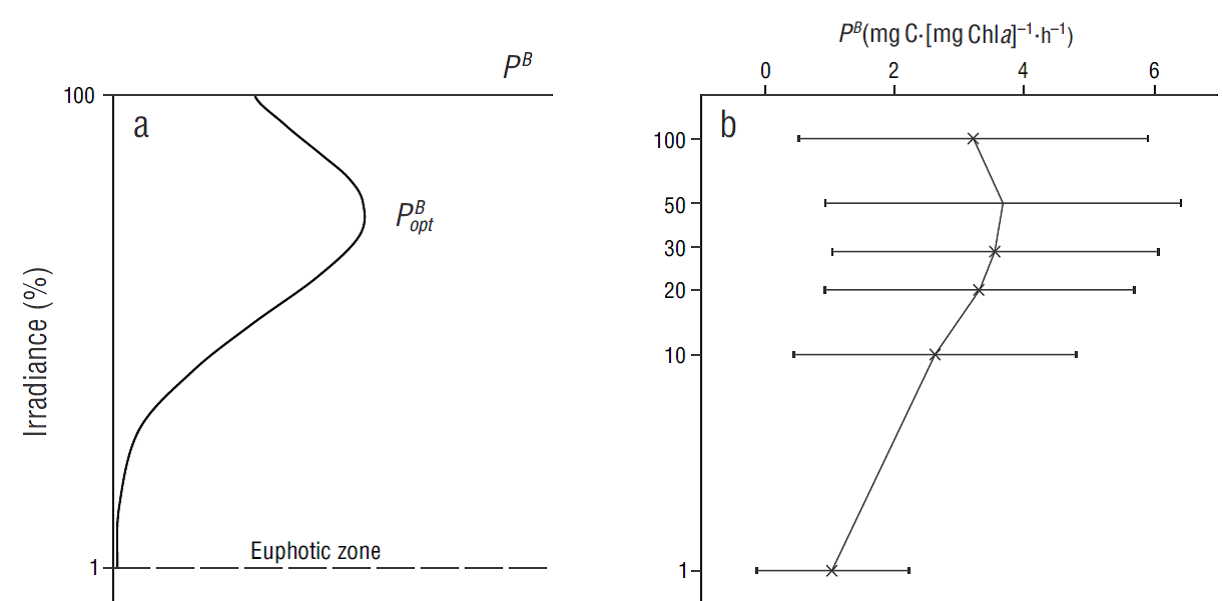
Figure 2 (a) Diagram of a vertical primary productivity profile normalized per unit of chlorophyll (P
B
) in relation to light attenuation (percentage); the position of the photosynthetic parameter
Average
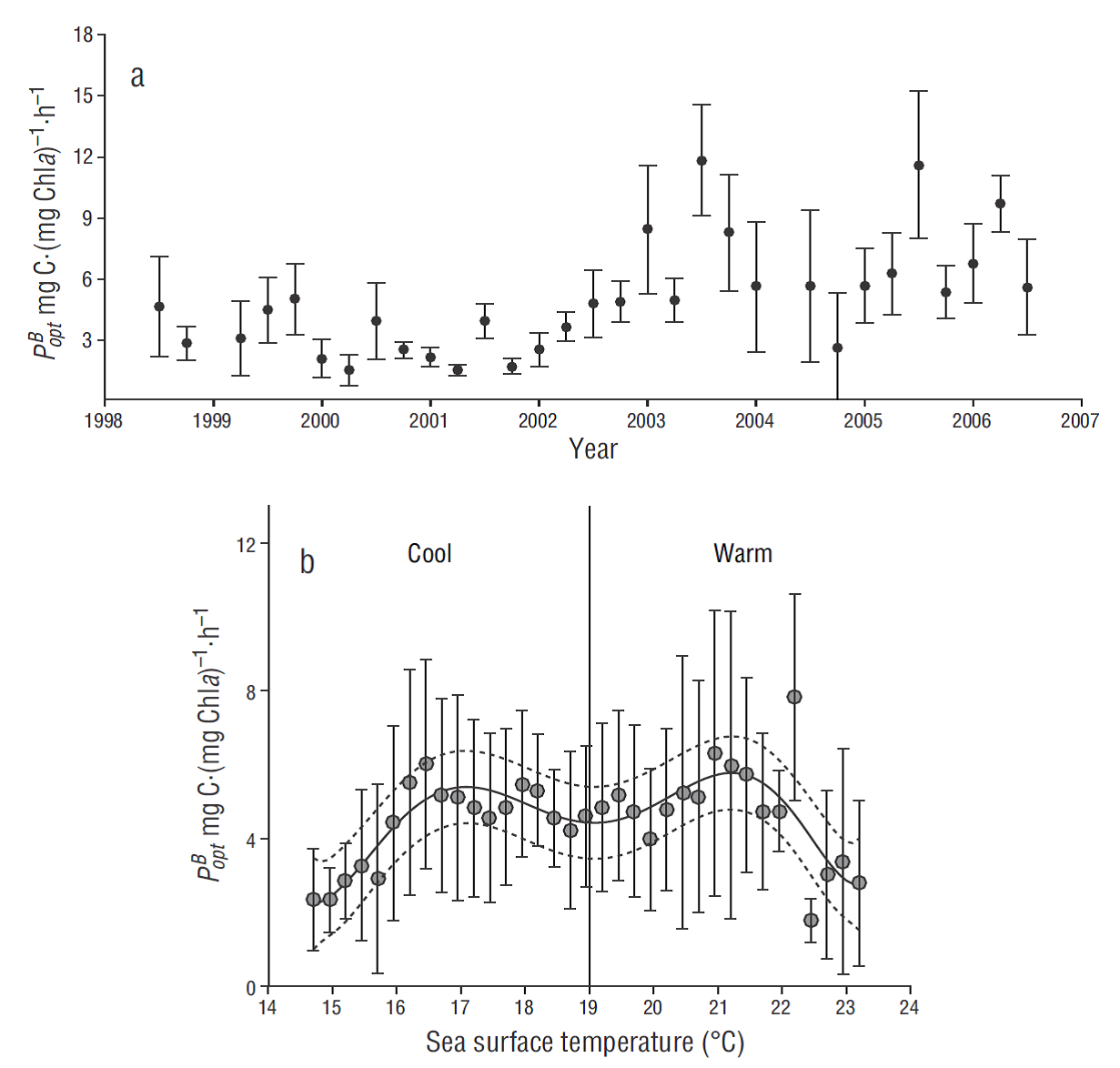
Figure 3 (a) Temporal variation of average optimal rates of carbon fixation (
In order to examine the importance of the 19 ºC surface isotherm as an indicator of the cold/warm regime change, the geographic position of this temperature value was determined on each monthly image (Fig. 4). In the region closest to the coast, the spatial dispersion of isotherms shows large latitudinal variability (~8º latitude) during the 25 years, which indicates alternating upwelling events. The average position (thick black contour) of the 19 ºC isotherm is located approximately 27ºN (off Punta Eugenia), and it is also asymmetrically located with respect to the spatial distribution of the isotherms.
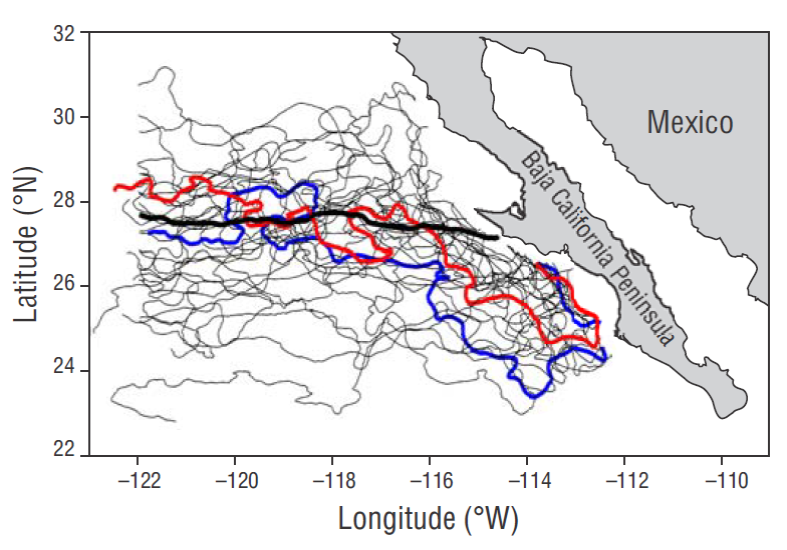
Figure 4 Monthly scatter of the 19 ºC surface isotherm from 1985 to 2009 (thin black lines). The thick black contour indicates the total average. Isotherms for January 1985 and January 2009 are shown in blue and red, respectively. The data correspond to level 3 monthly compounds calculated with AVHRR-PATHFINDER v5 .
The SST spatiotemporal evolution shows that the geographic amplitude of the 19 ºC isotherm in the study area responds mainly to the seasonal scale, with a latitudinal displacement of up to ~32ºN during summer and ~24ºN during winter (Fig. 5). In addition to the seasonal scale, the displacement of the isotherm shows the effect of warm and cold inter-annual events during some years. The temperature decrease in 1989, when the 19 ºC isotherm moved further south, and the cold events from 1999-2001 and 2006 are noteworthy. Warm periods were observed during 1992-1993, 1996-1998, 2002-2005, and 2007.
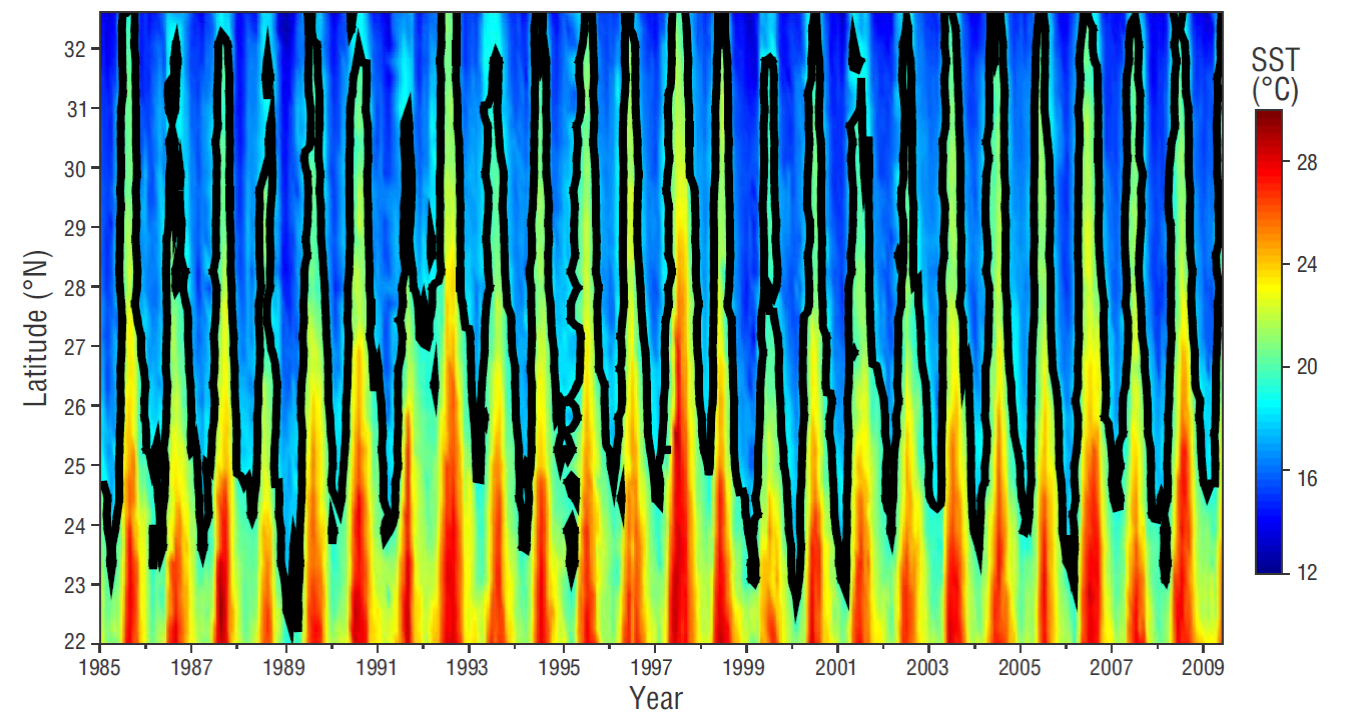
Figure 5 Spatiotemporal diagram of sea surface temperature (SST) variations for a transect located parallel to the coast (see Fig. 1).
The interannual variability was also observed in Chla concentrations at 20 m depth. The anomalies of the Chla regional average at 20 m depth, which is approximately the depth of the average
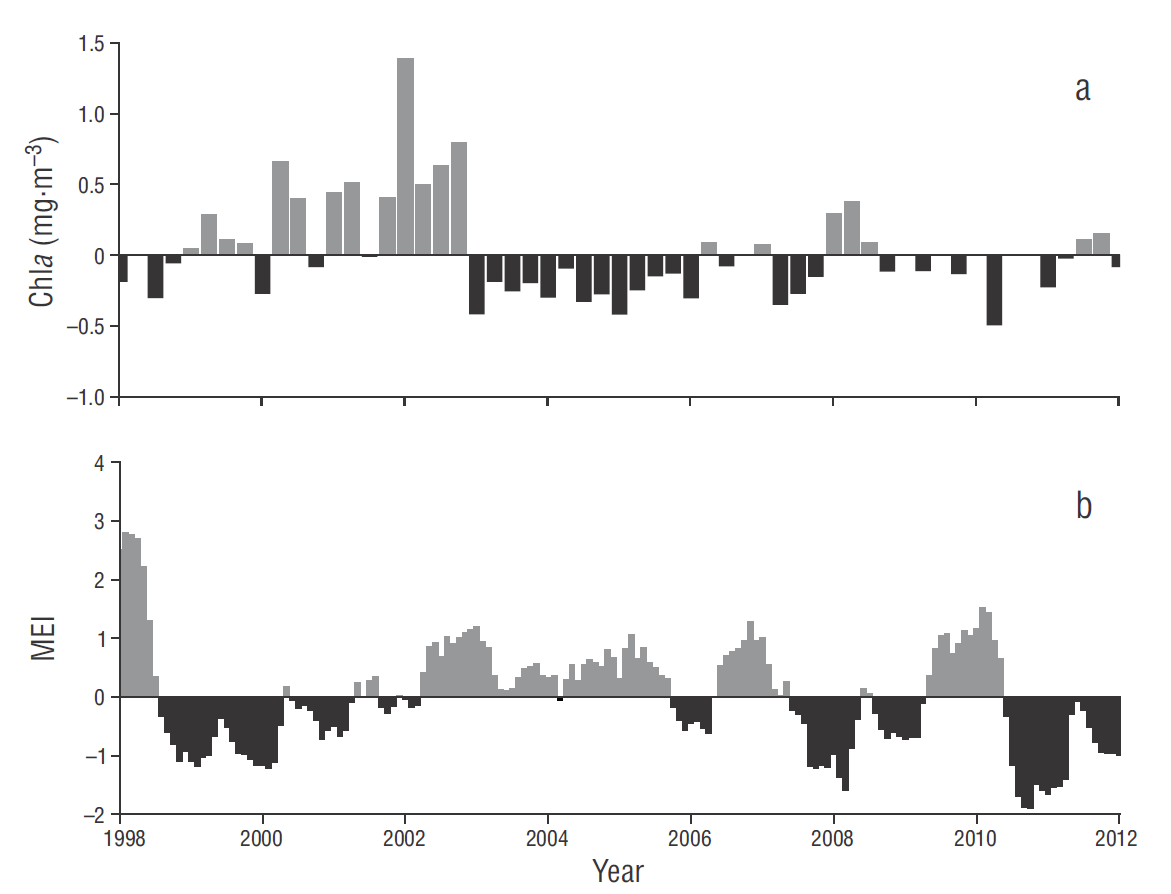
Figure 6 (a) Regional averages of chlorophyll (Chla) concentration anomalies (mgꞏm-3) measured at 20 m depth, which corresponds to the depth closest to the position of the optimal rate of carbon fixation (
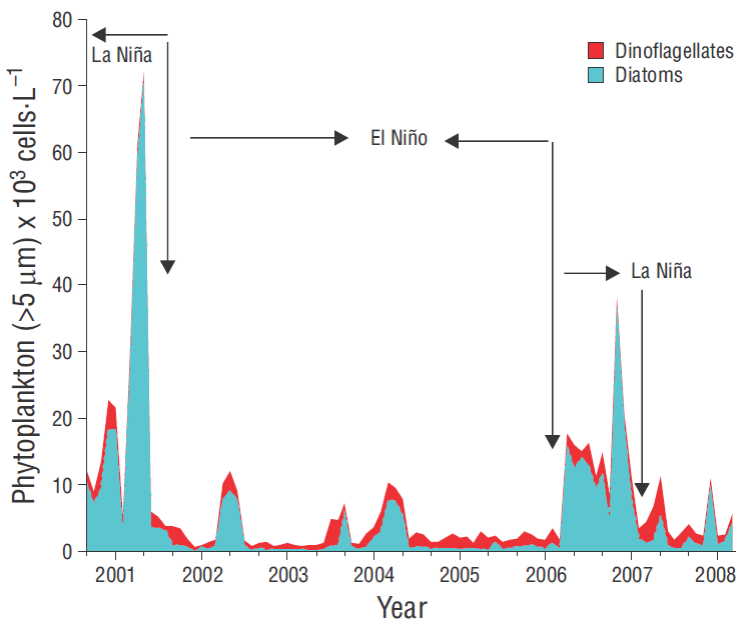
Data concerning nano -microphytoplankton abundances were only available for the winters (January) in the 2001-2008 period (Fig. 7) . Abundances were highest (70 × 103 cells L-1) during 2001, but they abruptly dropped to low concentrations (3-10 × 103 cells L-1) during the period 2002-2006. During 2006-2007, abundances again increased (20-40 × 103 cells L-1) and then decreased to less than 10 × 103 cells L-1 in 2008.
Discussion
At a global scale, carbon fixation via phytoplankton photosynthesis directly responds to climate variability (Behrenfeld et al. 2006) . In the last decades, changes in the oceanic conditions have favored a worrisome trend towards decreasing fixation rates. Primary productivity depends on light and nutrient availability, and these factors are controlled by different dynamic processes in the ocean at different scales (Behrenfeld et al. 2009) . PP trends have been difficult to assess due to the scarcity of available in situ data, and this is why a Chla-based index (Demarcq 2009) or models that use satellite-derived Chla data have been used (Behrenfeld et al. 2009); therefore, results are influenced by trends in Chla.
The photosynthetic parameter
The established empirical regional relation between SST and
The zone division into 2 regions (north and south), for which the boundary has been delineated at Punta Eugenia, off Baja California, has been extensively documented. Our results show that the average spatial distribution of the 19 ºC isotherm is related to the biogeographic zonation of the southern portion of the CCS (McClatchie et al. 2009), and its spatial variations clearly demonstrate the change in hydrographic conditions from cold to warm periods.
The interannual changes related to El Niño/La Niña that modify pycnocline depth generate negative Chla anomalies during warm events (deep pycnocline) and positive anomalies during cold events (shallow pycnocline) (Gaxiola et al. 2008, Espinosa-Carreón 2012). However, the changes that occurred between mid-2002 and late 2007 (El Niño) (Fig. 6b), which yielded negative Chla anomalies (Fig. 6a), caused high
Some of the main groups in the phytoplankton community composition of the southern region of the CCS are picoplankton (Prochlorococcus, Synecococcus, and picoeukaryotes; Almazán-Becerril et al. 2012) and nanomicrophytoplankton (several species of diatoms and dinoflagellates; Millán-Núñez et al. 2004, Millán-Núñez and Millán- Núñez 2010, Linacre et al. 2012). The contribution of picoplankton to biomass is of 46% during winter (Millán-Núñez and Millán-Núñez 2010), though this group is also present in the ecosystem throughout the year (Barocio-León et al. 2006, Almazán Becerril et al. 2012). Martínez-Almeida et al. (2014) reported that the contribution to picoplankton biomass in 2008 was almost constant throughout the year and that major changes occurred due to variations in diatom and dinoflagellate abundances. In addition, despite the low biomass, cells that make up picoplankton are able to contribute about 50% of global PP due to their large abundance (Casey et al. 2007, Johnson et al. 2006).
The worldwide trend towards increasing ocean temperatures affects the stability of the water column and the variability of the mixed layer, modifying nutrient availability for phytoplankton (Boyce et al. 2010) and carrying worrying consequences for each ecosystem. Nevertheless, these consequences are still not clear, for each region responds in different ways (Belkin 2009). The pelagic ecosystem off the coast of the Baja California Peninsula seasonally changes its hydrographic conditions from cold to warm periods due to its transitional nature. These oscillations have been intensified by interannual (El Niño/La Niña) and decadal events. Under these conditions, the response of primary producers has proven to be different in terms of biomass and carbon fixation rate or PP. In the northern region of the CCS, the data for integrated PP have shown a marked seasonal pattern. However, long-period trends that give evidence of climate change effects have not yet been reported (Kahru et al. 2009). Nevertheless, the clear patterns of seasonal and interannual variabilities with the temporal variation in Chla are consistent with changes in hydrographic conditions; Chla increases during cold periods and decreases during warm periods, with a long-period tendency to decrease in the oceanic zone (Behrenfeld et al. 2009). The response of primary producers in the southern region of the CCS has shown high

