










Serviços Personalizados
Journal
Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkCiencias marinas
versão impressa ISSN 0185-3880
Cienc. mar vol.42 no.4 Ensenada Dez. 2016
https://doi.org/10.7773/cm.v42i4.2628
Artículos
Caracterización espacial de las pesquerías en el Sistema de Cañones de Avilés (mar Cantábrico, España)
1Instituto Español de Oceanografía [Spanish Institute of Oceanography], Promontorio de San Martín SN, 39080 Santander, PO Box 240, Spain.
Las áreas marinas protegidas son una de las principales herramientas en la gestión pesquera basada en el ecosistema. Este enfoque requiere evaluar el papel de las pesquerías en el ecosistema mediante, por ejemplo, la identificación de tácticas pesqueras y el análisis de su distribución espacial, y esto permitirá identificar la coexistencia espacial de las actividades pesqueras y los hábitats o ecosistemas objeto de protección. En el Sistema de Cañones de Avilés (mar Cantábrico, España), hemos identificado el uso de 6 artes de pesca y 17 tácticas pesqueras, éstas últimas con una técnica de clasificación no jerárquica (CLARA). La distribución espacial de las actividades pesqueras demersales está fuertemente afectada por la topografía del área de estudio y la gran presencia de sustrato rocoso. Hay una clara segregación espacial entre tipos de artes de pesca, principalmente debido a los requerimientos operativos de las artes de pesca y la distribución de las especies objetivo. Esto minimiza los conflictos entre los distintos componentes de flota. Las tácticas pesqueras dirigidas a especies pelágicas se localizaron principalmente en la zona litoral y en la plataforma continental, mientras que aquellas dirigidas a especies demersales se distribuyeron por la plataforma continental y en el talud continental. Las tácticas pesqueras de palangre de fondo son las que tuvieron una distribución espacial más restringida.
Palabras clave: pesquerías; sistema de seguimiento de buques; distribución espacial del esfuerzo; Sistema de Cañones de Avilés; tácticas pesqueras
Marine protected areas are one of the main tools in ecosystem-based fishery management. This approach requires analyzing the role of fisheries in the ecosystem by, for example, identifying fishing tactics and analyzing the spatial distribution with sufficient precision to allow us to determine whether fishing activities and the habitat or ecosystem to be protected can coexist spatially. In the Avilés Canyon System (Cantabrian Sea, Spain), we detected 6 different gears and 17 fishing tactics were identified by a non-hierarchical classification technique (CLARA). The spatial distribution of demersal fishing activity is strongly affected by the topography of the study area and the large presence of rocky ground. There is a clear spatial segregation between gear types, mainly due to the operating requirements of the gears and the distribution of target species. This minimizes conflicts between different components of the fleet. The fishing tactics targeting pelagic species were mainly located on the coast and shelf, while those targeting demersal species were used on the shelf and slope. Longline fishing tactics are those that had a more restricted spatial distribution.
Key words: fisheries; vessel monitoring systems; spatial distribution of fishing effort; Avilés Canyon System; fishing tactics
Introducción
La actividad pesquera tiene importantes consecuencias tanto para las comundiades explotadas como para los ecosistemas donde tienen lugar estas actividades. La actividad pesquera es parte del ecosistema, y sus efectos son numerosos y están ampliamente estudiados (Hollingworth 2000, Cury et al. 2005). Entre los efectos más importantes, podemos destacar la destrucción de hábitats, la mortalidad de especies no objetivo, los cambios demográficos en las poblaciones y los cambios en el funcionamiento y estructura del ecosistema (Jennings y Kaiser 1998, Pikitch et al. 2004, Sánchez y Olaso 2004).
Este enfoque ha modificado las bases de la gestión pesquera dirigiéndose hacia lo que se ha llamado gestión pesquera basada en el ecosistema (Sinclair et al. 2002, Link 2005). De acuerdo con Pikitch et al. (2004), el principal objetivo bajo este enfoque es preservar la salud de los ecosistemas marinos y de las pesquerías que dependen de ellos. Por lo tanto, se han desarrollado los conceptos de aproximación ecosistémica a las pesquerías (AEP) y áreas marinas protegidas (AMP, incluidas las reservas marinas), el primero como método de gobernanza y el segundo como herramienta de gestión (Jones 2001, Pauly et al. 2002, Claudet y Pelletier 2004, García y Cochrane 2005). Para la implementación de una AEP, es necesario obtener la mejor comprensión posible del funcionamiento del ecosistema (Gislason et al. 2000, Sinclair et al. 2002, Fréon et al. 2005) teniendo en cuenta a las pesquerías como parte del ecosistema y no sólo como una actividad pesquera a regular. En este sentido, conocer el comportamiento de las actividades pesqueras, sus especies objetivo y la distribución espacial donde trabajan es esencial para el desarrollo de futuros planes de gestión que minimicen los efectos socioeconómicos en la actividad pesquera y maximicen la protección de los hábitats y las especies objetivo.
En este contexto, los países de la Unión Europea deben de crear una red de AMP que asegure la conservación y protección de hábitats, recursos y áreas de cría y reproducción, la conectividad entre áreas, etc. En muchas de las áreas seleccionadas como candidatas a AMP hay una continua e intensiva explotación pesquera y, por lo tanto, cualquier tipo de medida de regulación puede tener como consecuencia importantes conflictos sociales (Baelde 2005). Por consiguiente, es esencial tener conocimiento de la actividad pesquera y su relación con el ecosistema para caracterizar espacialmente los posibles efectos en el ecosistema.
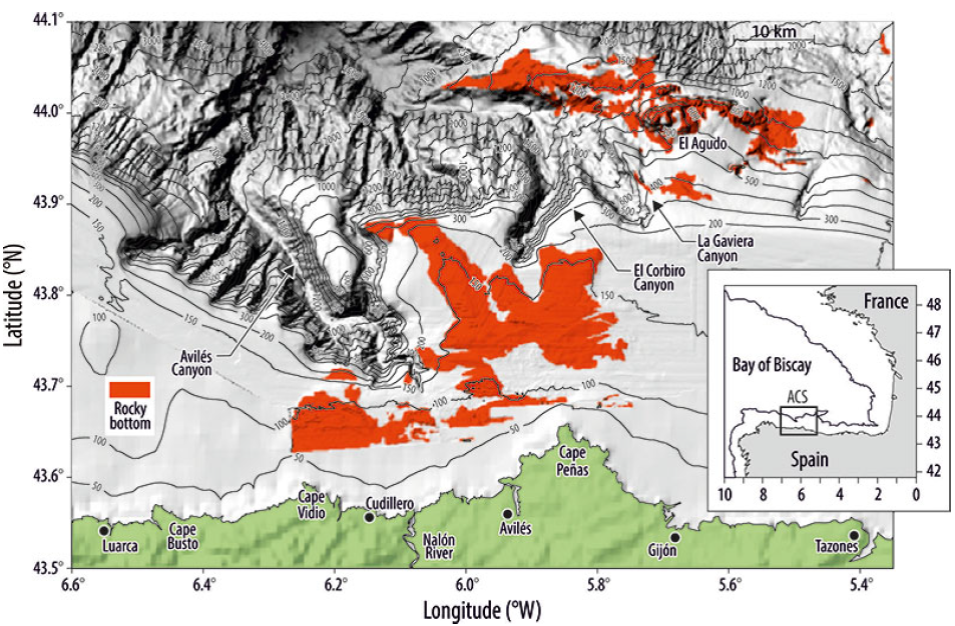
El Sistema de Cañones de Avilés (Fig. 1) es una de las 10 áreas elegidas por el Ministerio de Medio Ambiente en España a ser designadas AMP e integradas dentro de la red Europea Natura 2000. En su área de influencia, son utilizadas casi todos los tipos de arte de pesca (arrastre, palangre, cerco y enmalle). Esto hace de esta área un excelente ejemplo de cómo una zona estructural y geográficamente compleja (Gómez-Ballesteros et al. 2014) es explotada por pesquerías mixtas.
En la Unión Europea, la información de la distribución espacial de la actividad pesquera se puede obtener de 2 fuentes: cuadernos de pesca y datos de sistemas de seguimiento de buques (SSB). El acoplamiento de los datos de los SSB y cuadernos de pesca ofrece la posibilidad de obtener un análisis espacial preciso de la actividad pesquera y su relación con los ecosistemas afectados. Este estudio analiza y caracteriza por primera vez la actividad espacial de las pesquerías españolas por táctica pesquera (definida como una combinación de arte de pesca, especie objetivo y área de trabajo) en el sur del golfo de Vizcaya (España). Esto nos permitirá analizar la actividad pesquera según la especie objetivo y relacionar cada tipo de actividad a las características del área. Además podremos analizar el uso que hacen del área cada tipo de arte y táctica pesquera y su relación con las otras actividades pesqueras. Esta información deberá ser una de las herramientas principales para el desarrollo de los futuros planes de gestión, como el del AMP del Cañón de Avilés.
Materiales y métodos
Área de estudio
El área de estudio está localizada en el mar Cantábrico (sur del golfo de Vizcaya) y, por lo tanto, dentro de la Región IV de OSPAR. Toda la zona se caracteriza por una plataforma continental estrecha y cañones submarinos profundos. El Sistema de Cañones de Avilés (Fig. 1) es una región compleja de cañones y valles formada principalmente por 3 cañones de diferentes características morfoestructurales (Gómez-Ballesteros et al. 2014). La plataforma continental cercana es generalmente estrecha; el ancho varía de 12 km (en la cabecera del Cañón de Avilés) a 40 km. Las profundidades máximas son de unos 200 m y ocasionalmente hasta de 300 m al este del cañón. La plataforma continental muestra afloramientos rocosos de diferentes relieves y morfologías, y una superficie casi horizontal que corresponde a las zonas sedimentarias con cobertura de sedimentos no consolidados o superficies fuertemente erosionadas. Las fuertes corrientes sobre la plataforma continental son responsables del mínimo recubrimiento sedimentario en el área.
Fuentes de datos
La información de la batimetría se obtuvo durante 2 campañas llevadas a cabo en el B/I Vizconde de Eza entre 2010 y 2011 utilizando la ecosonda multihaz Simrad EM-300 (Gómez-Ballesteros et al. 2014).
La distribución espacial del esfuerzo de la flota industrial fue analizada utilizando la información de los cuadernos de pesca y de los SSB. La información de los cuadernos de pesca correspondió al periodo 2002-2010 para todo el mar Cantábrico (sur del golfo de Vizcaya), y fue utilizada para la identificación de artes de pesca y tácticas pesqueras (Marchal et al. 2006). La información de los SSB correspondió al periodo 2006-2010, y únicamente se usaron los datos correspondientes a la zona de estudio. Estos datos se utilizaron para el análisis espacial de las tácticas pesqueras identificadas previamente mediante los cuadernos de pesca. La información del SSB y los cuadernos de pesca fue facilitada por el Ministerio de Agricultura, Alimentacíon y Medioambiente (MAGRAMA) de España.
Respecto a los SSB, el dispositivo de localización envía cada 2 h una señal (ping) que registra el código del barco emisor, la fecha, la hora, la posición (latitud y longitud), la velocidad en nudos, el rumbo y si el barco está realizando operaciones de pesca o no. Se han aplicado muchos filtros y tipos de procesado para eliminar aquellas señales no asociadas a la actividad pesquera (Lee et al. 2010, Hintzen et al. 2011). En este estudio se utilizó el siguiente procesado: se obtuvó el intervalo de tiempo y la distancia euclidiana entre señales sucesivas, y cada uno de estos valores fue asociado a la primera señal del correspondiente par de puntos; se identificó el comienzo y la finalización de las mareas de pesca, cuando el tiempo transcurrido entre señales sucesivas fue más de 4 h; se calculó la velocidad media de la embarcación entre señales (pings) sucesivas; se eliminaron todas aquellas embarcaciones para las cuales hubiera menos de 10 señales a lo largo de un año; y se eliminaron todas aquellas señales que estaban a menos de 3 millas del puerto pesquero más cercano. A cada señal que coincidía con una marea de los cuadernos de pesca (en función del código del buque y la fecha de captura) se le asignó un arte de pesca y una táctica pesquera. En función de la distribución de frecuencias de velocidades medias, se definió un rango de velocidades de trabajo para cada arte de pesca y todas aquellas señales con velocidades fuera de rango se eliminaron (Hintzen et al. 2011). Para la zona de estudio, al 51.6% de los 46,075 días de pesca procedentes de los SSB se les pudo asignar un arte de pesca y una táctica pesquera a partir de la información de los cuadernos de pesca para el periodo entre 2006 y 2010.
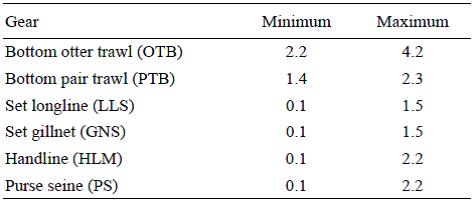
Para determinar los rangos de velocidades en los que se consideraba que había actividad pesquera, se utilizó la distribución de frecuencias de velocidades medias (Fig. S1). La identificación de rangos de velocidades de trabajo se puede hacer bien identificando los cambios de tendencia mediante modelos de regresión (regresión segmentada) o utilizando la información proporcionada por la flota y los observadores a bordo. Para las pesquerías móviles (arrastre y líneas de manos al arrastre), se utilizan ambos métodos de forma combinada, mientras que para las pesquerías estáticas (pesca con cerco, enmalle, palangre y línea de mano estática), es mejor usar la información de observadores a bordo y la procedente de la flota en función de la distribución de frecuencias. Los rangos de velocidades empleados en el trabajo por arte de pesca pueden verse en la Tabla 1.
Tabla 1 Rango de velocidades (nm) por arte de pesca en el que se consideraba que se estaba pescando.
En las plataformas continentales anchas con fondo sedimentario, como en el mar del Norte, las áreas de pesca generalmente son muy amplias. En estos casos, después de aplicar los filtros y analizar los datos del SSB, la distribución del esfuerzo de pesca puede ser observada claramente. Por lo tanto, son escasas las áreas con puntos que no corresponden a actividad pesquera. Sin embargo, las áreas de pesca sobre plataformas continentales estrechas con fondos rocosos distribuidos en mosaico o en el talud continental son generalmente angostas y cortas. En estos casos, es frecuente encontrar áreas donde aparentemente existe actividad pesquera, cuando en realidad no existe tal actividad. Para eliminar actividad pesquera falsa, es necesario aplicar umbrales para definir cuándo realmente existe actividad pesquera.
Se aplicaron umbrales para detectar si, una vez filtrados los datos procedentes de los SSB, los puntos correspondían realmente a actividad pesquera. A cada ping se le asignó el valor de esfuerzo que le correspondía (ver sección "Estimación de la distribución espacial del esfuerzo relativo"). Posteriormente, se aplicó el umbral de esfuerzo por debajo del cual se considera que la actividad no existe o es despreciable. Existen múltiples técnicas para definir este umbral (Jiménez-Valverde y Lobo 2007). Las que ofrecieron mejor resultado fueron las de aplicar umbrales en base a cuartiles. Se eliminaron puntos de presencia de actividad de pesca de acuerdo con la distribución de las frecuencias del esfuerzo (Fig. 2). La aplicación de otros estadísticos, basados en generar pseudo-ausencias para realizar la discriminación de puntos de presencia verdaderos, no fue fructífera. Esto se debe a que el esfuerzo se concentró en sitios específicos y, por lo tanto, al generar pseudoausencias y eliminar todas aquellas que coinciden con puntos reales de presencia, las pseudoausencias solamente cayeron en zonas donde realmente hubo ausencia de actividad; consecuentemente, los umbrales fueron tan bajos que no fue posible discriminar entre presencias reales y falsas. Los umbrales se pueden aplicar por arte de pesca o táctica pesquera. Si la importancia relativa del umbral es baja, es mejor aplicar los umbrales por táctica pesquera; en cualquier otro caso, es más efectivo aplicarlo por arte de pesca. En nuestro caso, al comparar la distribución del esfuerzo con la distribución de pings (Fig. 2), se observó que cuando se aplicó un umbral de 0.2, todos aquellos puntos que estaban fuera de las áreas de pesca fueron eliminados sin afectar la estimación del esfuerzo en las principales áreas de actividad pesquera. Después de evaluar la distribución espacial del esfuerzo para todas las artes de pesca, se aplicó el mismo umbral para todas ellas.

Identificación de tácticas pesqueras
La terminología utilizada en el presente trabajo fue definida por ICES (2003) y usada por Marchal et al. (2006): flota, grupo de barcos con similares características, y táctica pesquera, grupo de mareas dirigidas a la misma especie, población pesquera o una combinación de éstas usando un arte de pesca similar en el mismo periodo del año y en la misma área. La identificación de tácticas pesqueras fue hecha por arte de pesca. Para estandarizar las capturas por marea para cada una de las artes de pesca, la composición específica de cada una de las mareas se expresó en porcentaje respecto al total de la captura de la marea (Jiménez et al. 2004, Campos et al. 2007). Aquellas especies que representaron menos del 1% del total de las capturas se agruparon en la matriz bajo la denominación "otros". Dado que existen dudas de la clasificación de los tiburones de profundidad ya a bordo, este grupo taxonómico fue separado en 2 categorías: tiburones de profundidad y otros tiburones, que incluye el resto de los tiburones.
Con el objeto de identificar las tácticas pesqueras por arte de pesca, se realizó una agrupación no jerárquica denominada CLARA (Agrupación para Grandes Matrices, Kaufman y Rousseeuw 1986). Se utilizó el ancho de silueta (Rousseeuw 1987) para seleccionar el número de agrupaciones, y para validar la consistencia de la clasificación de la agrupación, se siguió el criterio de Struyf et al. (1996), en el cual se definen 4 niveles para el coeficiente global de silueta y para el coeficiente parcial de silueta: 0.71-1.00, estructura consistente; 0.51-0.70, estructura razonable; 0.26-0.50, estructura débil; <0.26, no existe un patrón. Una explicación detallada del uso de esta técnica en pesquerías puede verse en Castro et al. (2010).
Los análisis se realizaron con R (R Development Core Team 2005). Los análisis de componentes principales y de correspondencias se realizaron con la paquetería "ade4" y el análisis de agrupación se realizó con las paqueterías "cluster" (Maechler et al. 2005) y "stats" (R Development Core Team 2005).
Estimación de la distribución espacial del esfuerzo relativo
La distribución espacial del esfuerzo se estimó usando una rejilla de 250 m. A cada nodo se le aplicó un radio de búsqueda de 250 m y se sumó el tiempo asignado a cada ping de presencia que estaba dentro de cada radio. El asignar un radio igual al tamaño de la rejilla permite mejorar la precisión geográfica del esfuerzo, sobreestimando aquellas zonas donde existe mayor número de pings.
Estimación del solapamiento entre artes de pesca demersales
Se analizó el solapamiento entre las artes de pesca demersales: arrastre de fondo con puertas (OTB), arrastre de fondo a la pareja (PTB), enmalle fijo (GNS) y palangre de fondo fijo (LLS). A la hora de analizar el solapamiento entre actividades pesqueras, hay que tener en cuenta que el SSB indica la posición del barco, no la posición del arte de pesca. Con el objeto de evitar falsos positivos o falsos negativos, se tuvieron en cuenta valores mayores que el 20% respecto al valor máximo del esfuerzo, y la resolución de la capa fue rebajada a 1,000 m. Para analizar el solapamiento, a cada pixel con actividad de las capas analizadas se le asignó el valor 1, y se sumaron las capas de las artes de pesca a analizar (PTB vs OTB, GNS vs LLS, y artes de pesca fijos vs artes de pesca móviles). El resultado final fue una capa ráster con los siguientes valores: NA, ausencia de la actividad; 1, actividad de uno de los 2 tipos de artes analizados; y 2, solapamiento de la actividad.
Resultados
Tácticas pesqueras en el área de estudio
Todas las artes de pesca (OTB, PTB, GNS, LLS, cerco [PS] y línea de mano [HLM]), estuvieron presentes en el área de estudio. El palangre de fondo dirigido a Dicentrarchus labrax (lubina) y el cerco dirigido a Engraulis encrasicolus (anchoa) no se tuvieron en cuenta en el presente estudio porque estuvieron pobremente representados (3 y 6 mareas de pesca, respectivamente). Los valores del coeficiente global de silueta por arte de pesca fueron iguales o mayores que 0.5; por lo tanto, todas las clasificaciones podrían ser consideradas robustas (Tabla 2). Se identificaron 17 tácticas pesqueras. En todas las tácticas pesqueras la especie objetivo aparece en el 100% de las mareas de pesca (frecuencia de ocurrencia, FO), excepto en el arrastre con puertas mixto (OTB_MIX), el cerco dirigido a Trachurus spp. (PS_HOM) y la línea de mano dirigida a túnidos (HLM_TUN).
Tabla 2 Composición de la captura expresada en porcentaje de peso total y frecuencia de ocurrencia (FO) por arte de pesca (GCS, coeficiente global de silueta) y táctica pesquera (PCS, coeficiente parcial de silueta). Sólo se muestran las especies que superaron el 1% de la descarga.

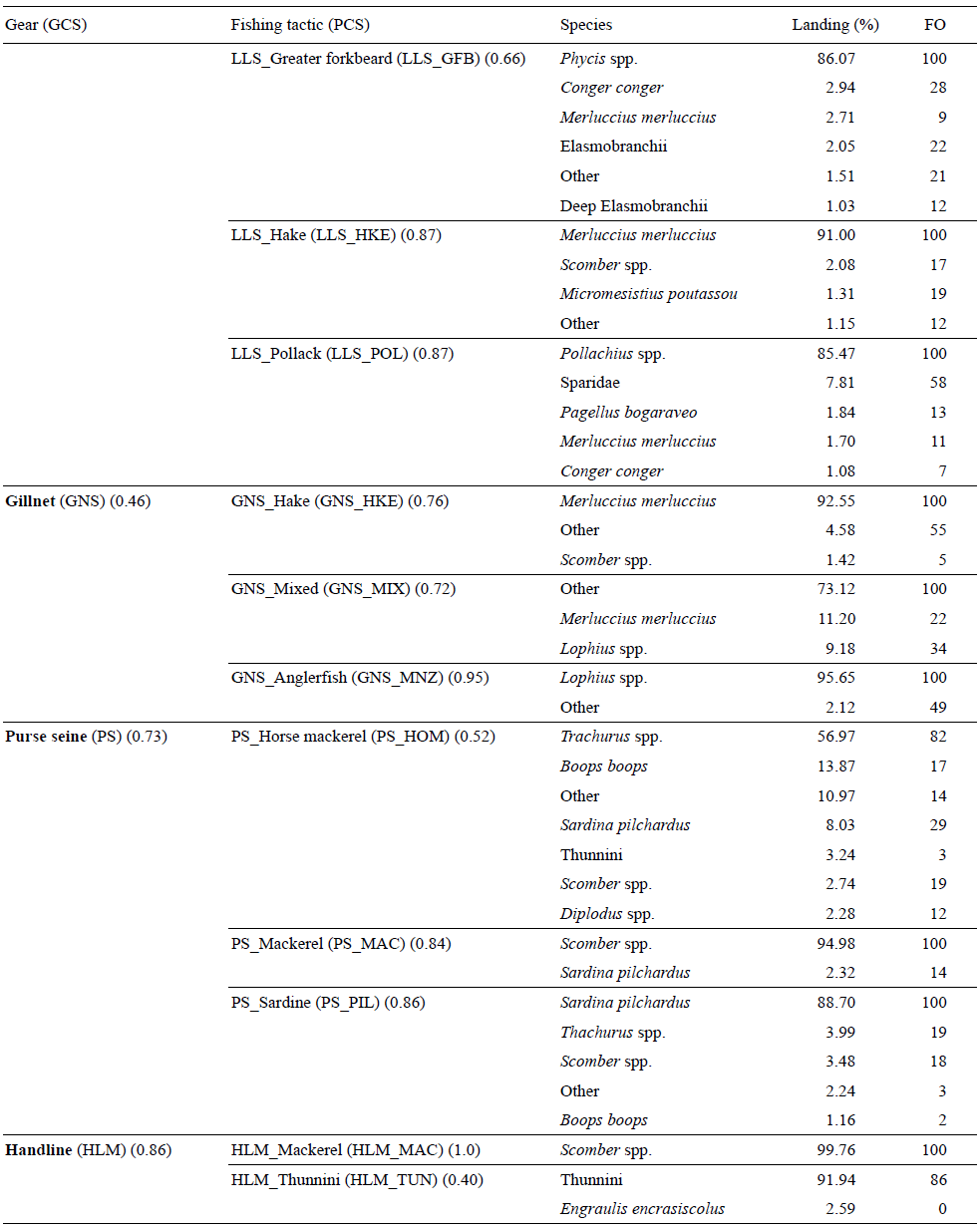
Se identificaron 3 tácticas pesqueras con arrastre de fondo con puertas. La pimera fue una táctica mixta (OTB_MIX) dirigida a varias especies demersales, ninguna de las cuales superó el 17% de los desembarques y casi el 45% de los desembarques correspondió a 3 especies (Lophius spp., rapes; Lepidorhombus spp., gallos; y Merluccius merluccius, merluza); las especies más importantes en FO fueron Lophius spp. La segunda táctica pesquera fue dirigida a Trachurus spp. (jurel) (OTB_HOM), el cual supone el 80% de las capturas, con alguna especie demersal como captura accesoria. La tercera táctica fue dirigida a Scomber spp. (caballa) (OTB_MAC), y Trachurus spp. fue captura accesoria (FO = 43). En el caso del arrastre a la pareja, se identificaron 2 tácticas pesqueras, la primera dirigida a Micromesistius poutassou (lirio) (PTB_WHB) con Merluccius merluccius (merluza) como principal especie no objetivo (FO = 80) y la segunda dirigida a Scomber spp. (PTB_MAC) con lirio como principal especie accesoria (FO = 43).
Se identificaron 4 tácticas pesqueras con palangre de fondo fijo. Una táctica fue dirigida a Conger conger (congrio) (LLS_COE) y otra a Phycis blennoides (locha) (LLS_GFB), y cada una de estas especies fue la principal especie accesoria de la otra táctica. La tercer táctica fue dirigida a merluza (LLS_HKE), sin que hubiera ninguna otra especie accesoria relevante, y la cuarta fue dirigida a Pollachius spp. (LLS_POL) con los espáridos como principales especies accesorias, las cuales fueron descargadas en el 58% de las mareas de pesca (FO) y compusieron sólo el 7.81% de las capturas en peso. En todas las tácticas pesqueras, las especies accesorias tuvieron poca importancia en comparación con las especies objetivo, tanto en FO como en peso.
Tres tácticas pesqueras emplearon redes de enmalle de fondo. Una fue dirigida a la merluza (GNSHKE) y otra a Lophius spp. (rapes) (GNS MNZ), sin que hubiera una especie accesoria característica. La tercer táctica fue dirigida a especies del grupo "otros" (GNS_MIX) con la merluza y los rapes como principales especies accesorias.
En el área de influencia del Sistema de Cañones de Avilés, el cerco y la línea de mano fueron utilizados en la captura de especies pelágicas. En el caso de la red de cerco, se identificaron 3 tácticas pesqueras: una dirigida al jurel (PS_HOM) con Boops boops (FO = 17) y Sardina pilchardus (FO = 29) como principales especies accesorias, otra dirigida a la caballa (PS_MAC) y la tercera dirigida a la Sardina pilchardus (sardina) (PS_PIL) con el jurel (FO = 19) y la caballa (FO = 18) como especies accesorias. Por último, se identificaron 2 tácticas pesqueras con línea de mano: una dirigida a la caballa (HLM_MAC) y la otra dirigida a los túnidos con cebo vivo (HLMTUN).
Distribución espacial de las pesquerías por tipo de arte de pesca
El arrastre de fondo con puertas se realizó en fondos sedimentarios por fuera de la isóbata de los 100 m, que es el límite legal (Fig. 3a). En la zona occidental del área de estudio, el esfuerzo estuvó distribuido cerca, pero por debajo, de los 200 m de profundidad, aunque se detectó actividad pesquera entre 100 y 200 m y a más de 500 m. En la zona oriental, la estructura fue similar, aunque la actividad fue más alta cerca de la isóbata de los 100 m. El arrastre a la pareja se llevó a cabo en 3 zonas, en los flancos del cañón de Avilés y en el afloramiento rocoso al sur de El Agudo entre las isóbatas de 200 y 500 m (Fig. 3b).
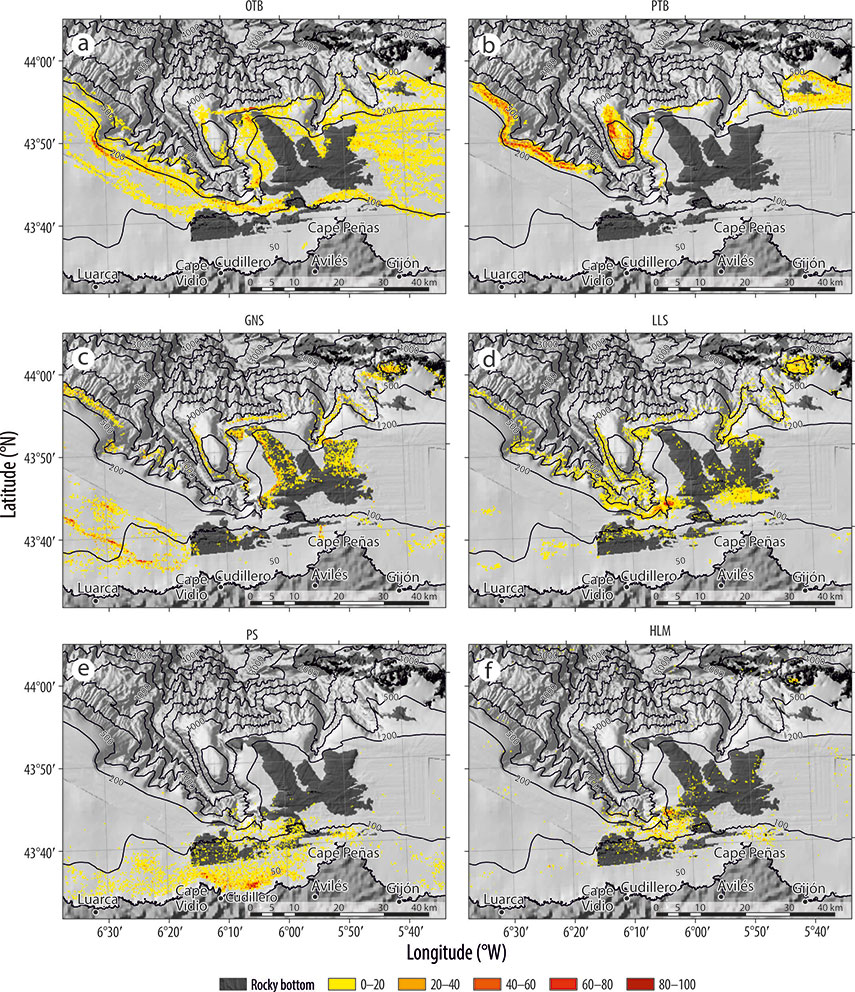
Figura 3 Distribución espacial por arte de pesca: (a) arrastre de fondo con puertas (OTB), (b) arrastre de fondo a la pareja (PTB), (c) enmalle de fondo fijo (GNS), (d) palangre de fondo fijo (LLS), (e) cerco (PS) y (f) línea de mano estática (HLM). La escala muestra el esfuerzo relativo.
En cuanto a los aparejos fijos, el enmalle (Fig. 3c) estuvo presente por casi toda la zona de estudio salvo en el extremo oriental cerca de los 200 m. Dos zonas concentraron la mayor parte del esfuerzo: la zona rocosa del este entre las isóbatas de 500 y 1,000 m, y la zona que va del oeste al sudeste cerca de los 100 m. Es necesario tener en cuenta que no tenemos información ambiental sobre esta última zona (Fig. 1), por lo que desconocemos si existen zonas rocosas.
En el caso de la pesquería con palangre de fondo, las 3 principales áreas de esfuerzo se localizaron sobre fondos rocosos: la cabecera del cañón de Avilés, el afloramiento rocoso de El Agudo y el área frente a cabo Peñas (Fig. 3d). Además, encontramos actividad pesquera en los flancos de los 3 cañones entre las isóbatas de 500 y 1,000 m de profundidad.
La mayor parte de la actividad pesquera con cerco se localizó en la zona occidental frente a cabo Peña por debajo de 100 m de profundidad (Fig. 3e). En cuanto a las líneas de mano, la actividad principal se localizó en la cabecera del cañón, aunque el esfuerzo pesquero se esparció por toda el área de studio (Fig. 3f).
Solapamiento de la actividad pesquera demersal
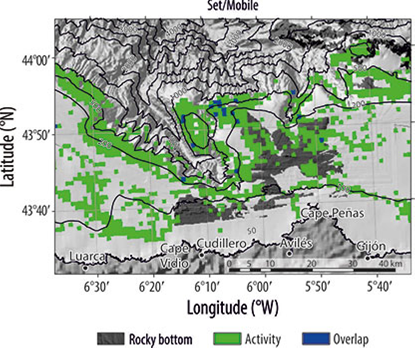
El solapamiento de los 2 tipos de arrastre ocurrió en la zona noroccidental, y fue escaso en la parte nororiental del cañón de Avilés (Fig. 4a). El solapamiento entre enmalle y palangre estuvo limitado a la cabecera del cañón y en El Agudo (Fig. 4b). Cuando se comparó la distribución espacial entre los tipos de artes de pesca, móviles y fijos, el solapa-miento fue muy bajo, y sólo ocurrió en la parte nororiental del cañón de Avilés (Fig. 5).
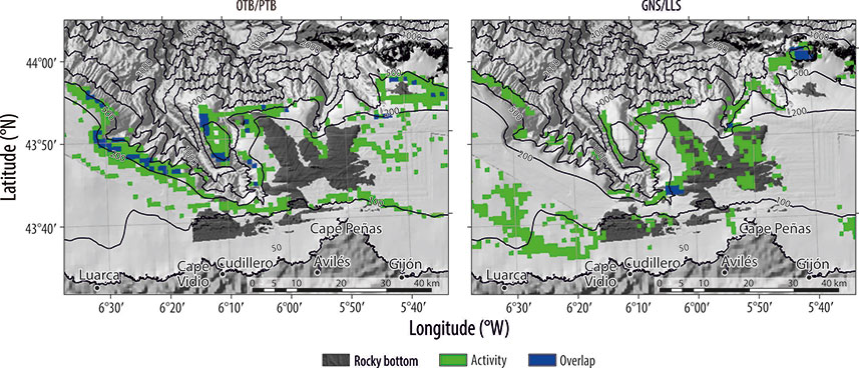
Figura 4 Distribución espacial del solapamiento entre pesquerías demersales: (a) arrastre de fondo con puertas (OTB) y arrastre de fondo a la pareja (PTB) y (b) enmalle de fondo fijo (GNS) y palangre de fondo fijo (LLS).
Distribución espacial de las pesquerías por táctica pesquera
La distribución espacial de cada una de las tácticas pesqueras puede verse en las Figuras S2-S5. En el caso de las tácticas con arrastre de fondo con puertas, la diferencia más importante fue que la táctica dirigida al jurel (Trachurus spp.) (OTBHOM) se llevó a cabo principalmente en la zona occidental y a lo largo de la isóbata de los 100 m en la zona oriental. Esta táctica pesquera y la táctica dirigida a la caballa (OTB_MAC) se desarrollaron principalmente a profundidades menores que 200 m (Fig. S2a-b). La táctica pesquera mixta (OTB_MIX) se aplicó en toda el área ocupada por este arte de pesca (Fig. S2c). La táctica OTBMAC se llevó a cabo durante los primeros 5 meses del año.
En el caso del arrastre de fondo a la pareja, el patrón de la distribución del esfuerzo con las 2 tácticas pesqueras es muy distinto. Mientras que la táctica dirigida al lirio (PTBWHB) ocupó toda el área de distribución del arrastre a la pareja (Fig. S2d), la táctica pesquera dirigida a la caballa se concentró en el borde continental del oeste del cañón de Avilés, entre las isóbatas de 200 y 500 m (Fig. S2e). Al igual que en el caso del OTB_MAC, la táctica PTB_MAC se llevó a cabo en los primeros 5 meses del año.
Las 3 tácticas pesqueras con el enmalle tuvieron una distribución espacial diferente (Fig. S3). La táctica pesquera mixta (GNS_MIX) pareció no tener un patrón claro, y se distribuyó en toda la zona donde estuvo presente el enmalle. La táctica pesquera de enmalle dirigida a la merluza (GNSHKE) se desarrolló en la zona occidental de la plataforma continental entre 100 y 200 m de profundidad y en la zona rocosa frente a cabo Peñas. Carecemos de información sobre el tipo de fondo en la zona occidental de la plataforma y en el extremo oriental a 100 m de profundidad. La táctica pesquera dirigida al rape (GNS_MNZ) se desarrolló a mayor profundidad, entre 500 y 1,000 m, y en los límites entre el sustrato blando y rocoso, alrededor de la zona rocosa frente a cabo Peñas.
La táctica pesquera con palangre de fondo dirigida a la locha (LLS_GFB) se desarrolló principalmente en zonas profundas de los flancos de los 3 cañones, entre 500 y 1,000 m (Fig. S4a). La táctica pesquera dirigida al congrio (LLS_COE), se llevó a cabo a menor profundidad, en la zona rocosa frente a cabo Peñas y a lo largo de la isóbata de 100 m (Fig. S4b). El esfuerzo con la táctica dirigida a la merluza (LLSHKE) se concentró en la cabecera del cañón de Avilés, en El Agudo y en los afloramientos rocosos situados frente a cabo Peñas (Fig. S4c). La táctica LLSPOL se aplicó principalmente a lo largo de la isóbata de 100 m (Fig. S4d).
La tácticas pesqueras con cerco dirigidas al jurel y a la sardina tuvieron una distribución espacial muy similar, y se realizaron muy cerca de la costa y al oeste de cabo Peñas (Fig. S5a, b). En el caso de la táctica dirigida a la caballa, el esfuerzo se concentró en la mitad de la plataforma continental, cerca de la cabecera del cañón (Fig. S5c). En lo que respecta a la estacionalidad, la táctica pesquera dirigida a la caballa, al igual que el resto de las tácticas que tuvieron a esta especie como objetivo, se llevó a cabo en los 4 primeros meses del año.
En el caso de la táctica pesquera con línea de mano dirigida a la caballa, el esfuerzo presentó una distribución muy similar a la de la táctica con cerco dirigida a esta misma especie (Fig. S5d). La táctica pesquera dirigida a túnidos se llevó a cabo en la zona rocosa de la cabecera del cañón de Avilés y en El Agudo (Fig. S5e).
Discusión
En las plataformas continentales estrechas donde existe una fuerte presión pesquera, como es el caso de la zona de estudio, las diferentes tácticas pesqueras suelen llevarse a cabo estratégicamente en zonas repartidas del espacio disponible para evitar, en lo posible, fricciones relativas al solapamiento de actividades pesqueras. En este sentido, y teniendo en cuenta la fuerte segregación espacial observada, los filtros empleados para eliminar todas aquellas señales que eran ajenas a la actividad pesquera fueron efectivos. La validez de estos filtros la podemos comprobar al analizar la distribución espacial del arrastre de fondo, ya que este aparejo puede arrastrar sólo en fondos sedimentarios y tiene restricciones legales muy precisas. Una vez aplicado el umbral, no hubo presencia de arrastre a menos de 100 m (veda espacial de pesca para el arrastre en la zona de estudio) ni en las entradas a los puertos de pesca. La flota que utiliza arrastre de fondo no puede trabajar en zonas rocosas y, como podemos comprobar, ninguna táctica pesquera con arrastre fue operada en la zona rocosa.
Para las artes de pesca ya descritas en estudios previos, la estructura de las tácticas pesqueras fue igual a la obtenida en este trabajo (Castro et al. 2010, Punzón et al. 2011). Entre las tácticas pesqueras identificadas, el enmalle mixto (GNSMIX) no debería ser considerado una táctica pesquera, a pesar de haber presentado un alto coeficiente parcial de silueta (Tabla 2). Este alto coeficiente puede deberse a la inclusión de un gran numero de especies pobremente representadas dentro del grupo de "otros". Además, hay que tener en cuenta que las principales especies accesorias fueron la merluza y el rape, y que la distribución espacial de GNSMIX parece ser el resultado de las distribuciones de GNSHKE y GNSMNZ; por lo tanto, es probable que las mareas de estas últimas tácticas pesqueras estén incluidas en la primera.
Para las plataformas continentales estrechas, el uso de los rectángulos estadísticos para definir si una pesquería debe ser considerada pesquería de profundidad, como LLS COE, puede ser difícil (Punzón et al. 2011). Como hemos visto, esta táctica pesquera se practica a lo largo de toda la plataforma continental, incluso en la zona próxima a la isóbata de 500 m, por lo que debería ser considerada una pesquería de profundidad. Por lo tanto, el SSB es una herramienta importante para caracterizar las pesquerías en plataformas continentales estrechas, especialmente las pesquerías que operan en áreas de especial interés donde se van a implementar medidas de gestión espacial.
En las revisiones realizadas por Piñeiro et al. (2001) y Punzón et al. (2011), los autores identificaron 2 pesquerías de profundidad en la región del Atlántico nororiental: GNS_MNZ y OTB MIX. En el caso de GNS_MNZ, el esfuerzo pesquero se localizó por debajo de 500 m de profundidad, mientras que en el caso de OTB_MIX, el esfuerzo a más de 500 m de profundidad se llevó a cabo en la región occidental y no fue muy intenso. Aunque se indentificó actividad pesquera de PTB_WHB y LLS_HKE cerca de la isóbata de 500 m, ésta tampoco fue muy importante, y no fue posible determinar si estas actividades pueden ser consideradas pesquerías de profundidad o no porque no fue muy claro si se llevaron a cabo por debajo de los 500 m.
La distribución espacial de las pesquerías por tipo de arte de pesca muestra que existe un escaso solapamiento en la zona de estudio, al menos en las zonas con altos niveles de esfuerzo. El escaso solapamiento implica que casi toda la zona de estudio está sometida a esfuerzo pesquero; sin embargo, hay poca competencia entre las técnicas pesqueras para recursos comunes. La casi inexistencia de solapamiento entre varios tipos de actividad debería permitirnos asignar técnicas pesqueras a aquellos pings a los que no se les pudo asignar la información de los cuadernos de pesca (Russo et al. 2011).
En el caso de los 2 tipos de arrastre con los que se trabaja en la zona, el arrastre con puertas ocupó una mayor área. Esto es debido a que la red de arrastre de fondo con puertas es más pequeña longitudinalmente y tiene menos abertura horizontal y, por lo tanto, las necesidades para trabajar son menos exigentes. Es decir, debido a su mayor maniobrabilidad, puede trabajar en caladeros de pesca más estrechos y angostos, y al tener menos superficie de contacto con el fondo, el terreno no necesita ser tan uniforme. El arrastre de fondo con puertas puede ser usado en los mismos caladeros de pesca donde se usa el arrastre a la pareja. Sin embargo, en estas áreas, la actividad del arrastre con puertas fue bajo. Esto es debido principalmente a que la especie principal objetivo del arrastre a la pareja es el lirio (Castro et al. 2010), y esta especie, que habita principalmente en el talud continental, dejo de ser objetivo para el arrastre a principios del siglo XXI (Punzón et al. 2010). Por lo tanto, al no competir por los mismos recursos, las pesquerías podrían ocupar espacios distintos. Para ambas artes de pesca, las zonas de arrastre, además de quedar restringidas por el tipo de fondo, siguen la batimetría intentando mantener la profundidad durante el arrastre e interferir lo menos posible con las pesquerías artesanales. En la región situada al este de la zona de estudio, la plataforma es más ancha y aplacerada. En esta área, las orientaciones del arrastre pueden cambiar y no seguir un patrón unico como en las zonas de talud o en zonas de fondo rocoso. Esto es posible debido a la poca ocupación que hacen las pesquerías fijas artesanales de esta región oriental de la plataforma continental, que queda de uso exclusivo del arrastre. Sin embargo, en el talud se incrementa considerablemente la competencia por el espacio de las diferentes pesquerías, lo que las obliga a mantener rumbos fijos.
Los solapamientos más importantes entre las tácticas pesqueras se dan entre el enmalle dirigido al rape y el palangre dirigido a la merluza en la zona del afloramiento rocoso de El Agudo. Esta es una zona muy importante para la pesca con enmalle dirigida al rape, mientras que para la pesca con palangre dirigida a la merluza, en cuanto a esfuerzo, se trata de una zona secundaria. Para determinar el grado de conflicto de uso entre aparejos, sería necesario determinar si esta zona es explotada por flotas con alguna singularidad local.
Es importante destacar que la distribución de las distintas tácticas pesqueras depende de las características del fondo y, en el caso de los sustratos blandos, del tipo de sustrato. Esta variable condiciona, en gran medida, la distribución espacial de cada una de las tácticas pesqueras en la zona de estudio y, sobre todo, el tipo de aparejo a utilizar. Esto afecta a la interpretación de la actividad pesquera. Así, por ejemplo, el GNS_HKE, en la zona donde disponemos de información del tipo de fondo, está distribuido en la zona rocosa, mientras que en la zona occidental de la plataforma, en donde carecemos de la información del tipo de fondo, parece tener una distribución espacial que invita a pensar que sigue un patrón asociado a algun tipo de fondo específico. Por lo tanto, es difícil en muchas ocasiones definir la competencia espacial entre aparejos sin esta información.
La merluza, la caballa y el jurel fueron especies objetivo para más de una táctica pesquera en la zona de estudio. En el caso de la merluza, el solapamiento espacial entre las 2 tácticas pesqueras (LLSHKE y GNSHKE) no fue importante. Mientras que con el palangre se explota principalmente la zona de la cabecera del cañón de Avilés, con el enmalle se explota la zona rocosa frente al cabo Peñas y la zona occidental entre 100 y 200 m. Se apreció algo de solapamiento solamente en El Agudo, aunque el esfuerzo de palangre en esa zona fue muy bajo.
Entre el final del invierno y el comienzo de la primavera una parte importante del esfuerzo se dirigió a la caballa durante la migración reproductora de esta especie (Uriarte y Lucio 2001, Villamor et al. 2011). La mayoría de los aparejos utilizados en la zona de estudio son utilizados para la captura de la caballa (Punzón et al. 2004). Este efecto debería provocar un descenso en el esfuerzo de otras tácticas pesqueras, pero sólo se detectó en febrero y marzo para la táctica con arrastre de fondo con puertas dirigida al jurel. Todas las tácticas pesqueras dirigidas a la caballa ocupan toda la plataforma continental por fuera de cabo Peñas. De todas las pesquerías dirigidas a pequeños y medianos pelágicos, la pesquería de la caballa es la que se desarrolla más alejada de la costa de forma general. Solamente el esfuerzo con cerco mantuvo cierta actividad cerca de la costa, aunque la mayor parte del esfuerzo es por fuera de la isóbata de los 100 m. Esto puede deberse a que durante su migración desde el este hacia el oeste, y al verse obligada a evitar el cabo Peñas, la caballa presenta las mayores abundancias en la plataforma media y externa.
La información de los SSB combinada con la de los cuadernos de pesca nos ha permitido estudiar no sólo la distribución espaciotemporal de la actividad pesquera, sino otros aspectos que son fundamentales en la gestión de los recursos y los ecosistemas que los soportan. Nos permite completar conocimientos acerca de la estructura de los ecosistemas, a nivel de especies comerciales, además de información de áreas no accesibles para las campañas de investigación basadas en lances de arrastre que se desarrollan en la zona (Sánchez y Serrano 2003), generalmente limitadas tanto espacial (sólo en fondos sedimentarios) como temporalmente (sólo en otoño). Además nos permite realizar estudios sobre el comportamiento de las especies de interés comercial, ya que las señales del SSB asociadas a una especie capturada se pueden interpretar como presencia de esa especie. Estas características se pueden combinar con información ambiental procedente de otras fuentes y, por lo tanto, podrían permitirnos conocer mejor las condiciones ambientales en las que se realiza la captura y consecuentemente los hábitats de las especies.
Una vez obtenidos los patrones de distribución espacial de la actividad pesquera y las variables ambientales y biológicas que los condicionan, se puede modelar el comportamiento de la actividad pesquera como respuesta a determinadas medidas de gestión. Por lo tanto, antes del establecimiento de un área marina protegida, por ejemplo, se podrá simular la reubicación espacial del esfuerzo en caso de que se excluyan o modifiquen determinadas actividades. Esto nos permitirá, además, estimar las consecuencias sobre la actividad pesquera, los efectos sociales y económicos, y los cambios sobre la mortalidad de las especies objetivo.
Agradecimientos
Este estudio fue parcialmente financiado por el proyecto TEPESCO (2016, Instituto Español de Oceanografía) y el programa LIFE+ de la Comisión Europea (proyecto INDE-MARES 07/NAT/E/000732), el cual fue coordinado por la Fundación Biodiversidad del Ministerio de Medio Ambiente de España. La información de la batimetría y las características del suelo fueron el producto del trabajo invaluable de María Druet, María Gomez-Ballesteros y Araceli Muñoz-Recio.
REFERENCIAS
Baelde P. 2005. Interactions between the implementation of marine protected areas and right-based fisheries management in Australia. Fish. Manage. Ecol. 12: 9-18. [ Links ]
Campos A, Fonseca P, Fonseca TJ, Parente J. 2007. Definition of fleet components in the Portuguese bottom trawl fishery. Fish. Res. 83: 185-191 [ Links ]
Castro J, Punzón A, Pierce GJ, Marín M, Abad E. 2010. Identification of métiers of the northern Spanish coastal bottom pair trawl fleet by using the partitioning method CLARA. Fish. Res. 102: 184-190. [ Links ]
Claudet J, Pelletier D. 2004. Marine protected areas and artificial reefs: A review of the interactions between management and scientific studies. Aquat. Living Resour. 17: 129-138. [ Links ]
Cury PM, Mullon C, García SM, Shannon LJ. 2005. Viability theory for an ecosystem approach to fisheries. ICES J. Mar. Sci. 62: 577-584. [ Links ]
Dray S, Dufour AB. 2007. The ade4 package: Implementing the duality diagram for ecologists. J. Stat. Softw. 22: 1-20. [ Links ]
Fréon P, Drapeau L, David JH, Fernandez Moreno A, Leslie RW, Oosthuizen WH, Shannon LJ , van der Lingen CD. 2005. Spatialized ecosystem indicators in the southern Benguela. ICES J. Mar. Sci. 62: 459-468. [ Links ]
García SM, Cochrane KL. 2005. Ecosystem approach to fisheries: A review of implementation guidelines. ICES J. Mar. Sci. 62: 311-318. [ Links ]
Gislason H, Sinclair M, Sainsbury K, Boyle R. 2000. Symposium overview: Incorporating ecosystem objectives in fisheries management. ICES J. Mar. Sci. 57: 468-475. [ Links ]
Gómez-Ballesteros M, Druet M, Muñoz A, Arrese B, Rivera J, Sánchez F, Cristobo J, Parra S, García-Alegre A, González-Pola C, Gallastegui J, Acosta J. 2014. Geomorphology of the Avilés Canyon System, Cantabrian Sea (Bay of Biscay). Deep-Sea Res. (II): 106: 99-117. [ Links ]
Hintzen N, Bastardie F, Beare D, Piet G, Ulrich C, Deporte N, Egekvist J, Degel H. 2011. VMStools: Open-source software for the processing, analysis and visualization of fisheries logbook and VMS data. Fish. Res. 115: 31-43. [ Links ]
Hollingworth CE (ed.). 2000. Ecosystem Effects of Fishing. Proceedings of an ICES/SCOR Symposium held in Montpellier, France, March 1999. ICES J. Mar. Sci. 57: 465-791. [ Links ]
[ICES] International Council for the Exploration of the Sea. 2003. Study Group on the Development of Fishery-based Forecasts. ICES CM 2003/ACFM:08, Boulogne, France. [ Links ]
Jennings S, Kaiser M. 1998. The effects of fishing on marine ecosystems. Adv. Mar. Biol. 34: 201-352. [ Links ]
Jiménez MP, Sobrino I, Ramos F. 2004. Objective methods for defining mixed-species trawl fisheries in Spanish waters of the Gulf of Cádiz. Fish. Res. 67: 195-206. [ Links ]
Jiménez-Valverde A, Lobo J. 2007. Threshold criteria for conversion of probability of species presence to either-or presence-absence. Acta Oecol. 31: 361-369. [ Links ]
Jones PJ. 2001. Marine protected area strategies: Issues, divergences and the search for middle ground. Rev. Fish Biol. Fish. 11: 197-216. [ Links ]
Kaufman L, Rousseeuw PJ. 1986. Clustering large sets (with discussion). In: Gelsema ES, Kanal LN (eds.), Pattern Recognition in Practice II. Elsevier/North Holland, Amsterdam, pp. 405-416. [ Links ]
Lee J, South AB, Jennings S . 2010. Developing reliable, repeatable, and accessible methods to provide high-resolution estimates of fishing-effort distributions from vessel monitoring system (VMS) data. ICES J. Mar. Sci. 67: 1260-1271. [ Links ]
Link JS. 2005. Translating ecosystem indicators into decision criteria. ICES J. Mar. Science. 62: 569-576. [ Links ]
Maechler M, Rousseeuw PJ , Struyf A, Hubert M. 2005. Cluster: Cluster Analysis Basics and Extensions. R package version 1.10.2, http://cran.r-project.org/web/packages/cluster/. [ Links ]
Marchal P, Andersen B, Bromley D, Iriondo A, Mahévas S, Quirijns F, Rackham B, Santurtun M, Tien N, Ulrich C . 2006. Improving the definition of fishing effort for important European fleets by accounting for the skipper effect. Can. J. Fish. Aquat. Sci. 63: 510-533. [ Links ]
Pauly D, Christensen V, Guénette S, Pitcher TJ, Sumaila UR, Walters CJ, Watson R, Zeller D. 2002. Towards sustainability in world fisheries. Nature 418: 689-695. [ Links ]
Pikitch EK, Santora C, Babcock EA, Bakun A, Bonfil R, Conover DO, Dayton P, Doukakis P, Fluharty D, Heneman B, Houde ED, Link J, Livingston PA, Mangel M, McAllister MK, Pope J, Sainsbury KJ. 2004. Ecosystem-based fishery management. Science 305: 346-347. [ Links ]
Piñeiro C, Casas M, Bañón R. 2001. The deep-water fisheries exploited by Spanish fleets in the Northeast Atlantic: A review of the current status. Fish. Res. 51: 311-320. [ Links ]
Punzón A , Hernández C, Abad E , Castro J , Pérez N, Trujillo V. 2010. Spanish otter trawl fisheries in the Cantabrian Sea. ICES J. Mar. Sci. 67: 1-13. [ Links ]
Punzón A , Serrano A, Castro J , Abad E , Gil J, Pereda P. 2011. Deep-water fishing tactics of the Spanish fleet in the Northeast Atlantic. Seasonal and spatial distribution. Sci. Mar. 75: 465-476. [ Links ]
Punzón A , Villamor B, Preciado I. 2004. Analysis of the handline fishery targeting mackerel (Scomber scombrus, L.) in the north of Spain (ICES Division VIIIbc). Fish. Res. 69: 189-204. [ Links ]
R Development Core Team. 2005. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0, URL http://www.R-project.org. [ Links ]
Rousseeuw PJ . 1987. Silhouettes: A graphical aid to the interpretation and validation of cluster analysis. J. Comput. Appl. Math. 20: 53-65. [ Links ]
Russo T, Parisi A, Cataudella S. 2011. New insights in interpolating fishing tracks from VMS data for different métiers. Fish. Res. 108: 184-194. [ Links ]
Sánchez F, Olaso I. 2004. Effects of fisheries on the Cantabrian Sea shelf ecosystem. Ecol. Model. 172: 151-174. [ Links ]
Sánchez F , Serrano A . 2003. Variability of groundfish communities of the Cantabrian Sea during the 1990s. ICES Mar. Sci. 219: 249-260. [ Links ]
Sinclair M , Arnason R, Csirke J, Karnicki Z, Sigurjonsson J, Rune Skjoldal H, Valdimarsson G. 2002. Responsible fisheries in the marine ecosystem. Fish. Res. 58: 255-265. [ Links ]
Struyf A , Mia H, Rousseeuw PJ . 1996. Clustering in an object-oriented environment. J. Stat. Softw. 1(4): 1-30. [ Links ]
Uriarte A, Lucio P. 2001. Migration of adult mackerel along the Atlantic European shelf edge from a tagging experiment in the south of the Bay of Biscay in 1994. Fish. Res. 50: 129-139. [ Links ]
Villamor B , González-Pola C , Lavín A, Valdés L, Lago de Lanzón A, Franco C, Cabanas JM, Bernal M, Hernández C , Iglesias M, Carrera P, Porteiro C. 2011. Environmental control of Northeast Atlantic mackerel (Scomber scombrus) recruitment in the southern Bay of Biscay: Case study of failure in the year 2000. Fish. Oceanogr. 20: 397-414. [ Links ]
Recibido: Diciembre de 2015; Aprobado: Octubre de 2016
 This is an open-access article distributed under the terms of the Creative Commons Attribution License
This is an open-access article distributed under the terms of the Creative Commons Attribution License
