











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
En los últimos 2 siglos la actividad antropogénica ha incrementado la concentración atmosférica de CO2 en ~400 petagramos (1 Pg = 1015 g), lo que representa un aumento de ~40% (Zeebe 2012). El contenido de CO2 atmosférico ha cambiado de 280 μatm antes de la revolución industrial a 407 μatm en tiempos recientes (NOAA 2016), y puede llegar a 850 μatm en el año 2100 (Hoegh-Guldberg et al. 2014). Los océanos absorben alrededor de 2 Pg C año-1 del CO2 emitido por las actividades antropogénicas (Takahashi et al. 2009); sin embargo, la tasa de absorción no es lo suficientemente alta para evitar el incremento del CO2 en la atmósfera. Debido a que el CO2 es un gas de efecto invernadero, las emisiones excedentes recientes han generado un aumento de 0.85 °C en la temperatura promedio del planeta y se ha anunciado un incremento de ~4 °C para 2100 relativo a la media global estimada de 1980 a 1999 (IPCC 2014).
En promedio, los océanos funcionan como un sumidero neto de CO2, pero la dirección y magnitud del intercambio de CO2 con la atmósfera varía considerablemente en espacio y tiempo. Hales et al. (2012) caracterizaron a la región del océano Pacifico entre 22°N y 50°N (30 a 370 km frente a la costa) como una fuente de CO2 de 0.60 mol C m-2 año-1, valor 3 veces superior al estimado por Chavez et al. (2007) para la misma zona. De La Cruz-Orozco et al. (2010) evaluaron el intercambio de CO2 entre el mar y la atmósfera frente a la península de Baja California de octubre de 2004 a octubre de 2005 y, en promedio, la zona actuó como una fuente de CO2 con un flujo anual de 0.40 mol m-2. A partir de datos de la presión parcial de CO2 (pCO2) recolectados de 1993 a 2001 frente a Baja California por barcos de oportunidad, Hernández-Ayón et al. (2010) concluyeron que en un balance anual la región es una fuente de CO2, aunque presenta condiciones de sumidero en invierno y primavera y condiciones de fuente en verano y otoño.
En la corriente de California, se ha detectado agua corrosiva (pH menor que 7.80) cercana a la superficie, la cual se esperaba incidiera hasta dentro de 50 años (Feely et al. 2008, Alin et al. 2012). Aunque parte de estas características corrosivas se deben a procesos naturales de la respiración en aguas subsuperficiales, la acumulación de CO2 atmosférico continúa y está modificando la dinámica natural del ciclo del CO2 en las zonas de surgencias costeras (Feely et al. 2008).
Los eventos de gran escala como el ciclo de El Niño Oscilación del Sur (ENOS) afectan el flujo de carbono entre el mar y la atmósfera (FCO2). Durante el evento de El Niño 1997-1998, en la zona de surgencias costeras de la región central de la corriente de California (al norte de nuestra área de estudio), el FCO2 fluctuó entre -0.30 y -0.70 mol C m-2 año-1, pero los aportes fueron de 1.50 a 2.20 mol C m-2 año-1 bajo condiciones de La Niña 1999 (Friederich et al. 2002). En la zona costera del Pacífico tropical frente a México (al sur de nuestra área de estudio), el FCO2 fue de -0.40 a 2.50 mmol C m-2 d-1 durante el evento de El Niño 2009 y de -4.40 a 3.30 mmol C m-2 d-1 durante el evento de La Niña 2010 (Franco et al. 2014). Para la región sur de la corriente de California frente a la península de Baja California, nuestra área de interés, se espera que el flujo sea del océano a la atmósfera en condiciones de El Niño y en dirección inversa en condiciones de La Niña.
Con un análisis de datos de alta frecuencia (cada 3 h) de la pCO2 obtenida por medio de sensores localizados en una boya autónoma anclada frente a la costa norte de Baja California, se determinó que esta zona costera es una fuente neta de CO2 hacia la atmósfera (~1.00 mol C mm-2 año-1) (Reimer et al. 2013b, Muñoz-Anderson et al. 2015). Reimer et al. (2013b) concluyeron que durante los eventos de surgencias costeras hubo un aporte del 91% de los valores de flujo positivo (fuente) de CO2, y Muñoz-Anderson et al. (2015) encontraron que el sistema es un leve sumidero (0.06 mol C m-2 año-1) durante los periodos sin surgencias.
Son escasos los trabajos publicados tendientes a resolver la variabilidad estacional de los flujos de CO2 entre el mar y la atmósfera en la región sur de la corriente de California frente a Baja California. En este artículo se presenta el análisis de la pCO2 y el FCO2 calculado con datos in situ de 22 cruceros oceanográficos. Esta base de datos nos permite sostener que los resultados son robustos para la zona de estudio y contribuyen de forma significante al conocimiento acerca de la variabilidad espacial y estacional de la pCO2 y el FCO2 en esta región. También indican que las variaciones temporales y los controles intrínsecos están sujetos a variaciones espaciales muy dinámicas y deben ser examinados de forma individual en cada dominio espacial.
Materiales y métodos
El océano Pacífico frente a la península de Baja California presenta alta variabilidad espacial y temporal ocasionada principalmente por la dinámica física derivada de cambios en la intensidad del flujo de la corriente de California (Durazo y Baumgartner 2002); eventos de surgencias costeras (Pérez-Brunius et al. 2007); la presencia de frentes, meandros y remolinos (Barocio-León et al. 2007); y la variabilidad estacional e interanual (Aguirre-Hernández et al. 2004, Gaxiola-Castro et al. 2008). Debido a su variabilidad estacional, la región de estudio, que comprende la región sur de la corriente de California, se ha dividido en porción norte y porción sur a partir de 28°N de latitud (Durazo 2015).
En la región sur de la corriente de California, el programa Investigaciones Mexicanas de la Corriente de California (IMECOCAL) realiza cruceros oceanográficos 4 veces al año (invierno, primavera, verano y otoño) frente a la península de Baja California, entre ~30 km frente a la costa y 220 km en mar abierto. La malla regular de muestreo se conforma de 90 estaciones, separadas por ~37 km y arregladas en líneas perpendiculares a la costa; la distancia entre las líneas hidrográficas es de ~70 km (Fig.1a). En este trabajo se analizó la información hidrográfica y de la pCO2 superficial obtenida durante 22 cruceros realizados entre 2004 y 2011 (Fig. 1b).

Figura 1 (a) Zonificación del área de estudio y sitios de muestreo del programa IMECOCAL. Los círculos y cuadros vacíos indican las estaciones en la zona oceánica norte y zona oceánica sur, respectivamente. Los círculos y cuadros rellenos indican las estaciones en la zona costera norte y zona costera sur, respectivamente. (b) Distribución del número de estaciones hidrográficas visitadas durante los cruceros IMECOCAL de 2004 a 2011.
Durante las campañas oceanográficas, se hicieron lances con un CTD Sea-Bird Electronics 911 Plus, calibrado por el fabricante, acoplado a una roseta con botellas Niskin de 5 L de capacidad. El CTD contenía sensores para medir temperatura, salinidad y oxígeno disuelto. De manera adicional, se tomaron muestras de agua con botellas Niskin de 5 L de capacidad a las profundidades discretas de 0, 10, 20, 50, 100, 150 y 200 m para el análisis de oxígeno disuelto y clorofila a del fitoplancton.
Las muestras para el análisis de oxígeno disuelto se recolectaron en botellas de vidrio de 125 mL y se analizaron en el laboratorio del barco con el método microWinkler (Helm et al. 2009). El coeficiente de correlación lineal entre los valores discretos de oxígeno disuelto obtenidos por el método microWinkler y los valores derivados del CTD fue de 0.96 (n = 346, P < 0.05). A partir de los datos de oxígeno del CTD, se calculó el porcentaje de saturación de oxígeno (%SO) con las ecuaciones de Weiss (1970).
Las alícuotas de agua para el análisis de clorofila a se recolectaron en botellas de plástico oscurecidas de 1 L de capacidad, y la concentración se determinó con los fluorímetros Turner Designs 10-AU-05 y Trilogy (modelo 7200-000) con el método de Holm-Hansen et al. (1965) modificado por Venrick y Hayward (1984). Más detalles de los métodos de recolecta de las muestras y los análisis utilizados por el programa IMECOCAL se pueden consultar en http://imecocal.cicese.mx.
La pCO2 en la superficie del mar (pCO2mar) se evaluó con un sistema LI-COR 6262 de flujo continuo totalmente automatizado que mide la fracción molar del CO2 en aire seco (xCO2). El agua de mar fue bombeada desde una toma localizada en el casco del buque (por debajo de la línea de flotación, ~2.50 m de profundidad); el agua fluía hacia el termosalinómetro y posteriormente al LI-COR 6262, ambos colocados en el laboratorio seco del buque. La precisión del LI-COR 6262 en las mediciones de la pCO2mar fue de ±1 μatm y se autocalibró cada 2 h con CO2 comercial estándar obtenido del National Institute of Standards Technology y la National Oceanic Atmospheric Administration Climate Monitoring Diagnostic Laboratory (Friederich et al. 2002). Para mayores detalles de la obtención de los datos de la pCO2 y su procesamiento, ver Reimer et al. (2013a). Debido a que los datos de la pCO2mar se obtuvieron de forma continua durante la navegación, fue necesario obtener un promedio durante el tiempo (~40 min) que el barco permaneció en cada estación de muestreo, con la finalidad de acoplarlos con los datos superficiales discretos de temperatura, salinidad, %SO y clorofila a.
El flujo de CO2 entre el océano y la atmósfera fue calculado con la siguiente ecuación: FCO2 = Kw x K0 x (ΔpCO2), donde Kw es el coeficiente de transferencia del CO2 en función de la velocidad del viento (Wanninkhof 2014), K0 es el coeficiente de solubilidad del CO2 en función de la temperatura y salinidad (Weiss 1974) y ΔpCO2 es la diferencia entre la pCO2mar y la pCO2 en la atmósfera (pCO2atm). Por convención, los valores negativos de FCO2 indican flujos de la atmósfera hacia el océano y los positivos indican flujos en sentido inverso. El CO2 en la atmósfera está bien mezclado y su presión parcial varía en un intervalo muy estrecho con respecto a su valor medio (Sarmiento y Gruber 2006); por ello, los datos diarios de la pCO2atm se obtuvieron de la página http://scrippsco2.ucsd.edu/data/atmospheric_co2. Estos datos corresponden a mediciones realizadas en el observatorio ubicado en Mauna Loa, Hawaii, de 2004 a 2011. A partir de esta información, se obtuvieron los promedios de la pCO2atm del periodo cercano a cada campaña oceanográfica, y con éstos se calculó un valor promedio de pCO2atm para cada uno de los 22 cruceros analizados. Los datos mensuales de la velocidad del viento en la superficie del océano (resolución espacial de 0.25° x 0.25°) se obtuvieron de http://podaac.jpl.nasa.gov/DATACATALOG/ccmpinfo.html, y son resultado de la calibración cruzada de varias multiplataformas (Atlas et al. 2011).
Se calculó la climatología estacional de la temperatura, salinidad, clorofila, %SO, pCO2 y FCO2 durante el periodo de muestreo (5 inviernos, 6 primaveras, 5 veranos y 6 otoños). Debido a que la malla de muestreo no siempre se cubrió en su totalidad, los promedios se calcularon con el número de datos disponible en cada crucero usando una media aritmética. Para minimizar errores de medición, se eliminaron aquellos datos cuyo valor estuvo fuera del intervalo de 3 desviaciones estándar. Para conocer la respuesta de la pCO2 y el FCO2 ante el ciclo ENOS en la región de estudio, se analizaron los efectos durante la fase cálida (El Niño), que impactó en el otoño de 2004, y la fase fría (La Niña), que impactó en la primavera de 2011, ambos eventos identificados de acuerdo con el Índice Multivariado de ENOS (http://www.cpc.ncep.noaa.gov/products/analysis_monitoring/ensos-tuff/ensoyears.shtml).
Resultados
La climatología estacional (2004-2011) de la temperatura superficial del mar mostró un gradiente latitudinal, donde los valores mayores se ubicaron en la porción sur (de la línea 120 hacia el sur) y los valores menores en la porción norte (arriba de la línea 120) (Fig. 2). La temperatura superficial del mar presentó 2 condiciones muy diferenciadas: valores menores en invierno (16.50 °C; Fig. 2a) y primavera (16.30 °C; Fig. 2b), y valores mayores en verano (19.90 °C; Fig. 2c) y otoño (20.20 °C; Fig. 2d). En general, la porción norte presentó un patrón longitudinal muy persistente de temperatura, en el cual los valores más bajos se presentaron cerca de la costa y los más altos en mar abierto. En la porción sur predominó el patrón latitudinal con isotermas perpendiculares a la costa, particularmente en verano y otoño (Fig. 2c, d).

Figura 2 Climatología estacional (2004-2011) de la temperatura superficial del mar (SST, °C) en (a) invierno, (b) primavera, (c) verano y (d) otoño.
La distribución promedio de la salinidad superficial del mar presentó un gradiente latitudinal con isohalinas perpendiculares a la costa (Fig. 3 a, c). Solamente en primavera se apreció un patrón longitudinal en la zona costera norte (ZCN), desde punta Eugenia hasta Ensenada (Fig. 3b), donde las isohalinas paralelas a la costa se extendieron ~50 km hacia mar abierto. Los valores promedio de salinidad fueron mayores en otoño (33.65; Fig. 3d) e invierno (33.62; Fig. 3a), y menores en primavera (33.53; Fig. 3b) y verano (33.57; Fig. 3c).

Figura 3 Climatología estacional (2004-2011) de la salinidad superficial del mar en (a) invierno, (b) primavera, (c) verano y (d) otoño.
La clorofila y el %SO presentaron un gradiente costa-océano, en el cual los valores mayores se ubicaron cerca de la costa y los menores hacia mar abierto. La mayor concentración promedio de clorofila se presentó en primavera (0.88 mg m-3; Fig. 4b) y las isolíneas de 0.50 mg m-3 se extendieron ~100 km hacia mar abierto. Es notoria la lengüeta de concentración de clorofila con valores superiores a los 0.50 mg m-3 que sale de punta Eugenia y se extiende hasta ~140 km fuera de la costa (Fig. 4a, c) en todas las estaciones del año. En la ZCN se presentaron los máximos valores relativos de clorofila en cada estación del año. Durante invierno y primavera, las concentraciones de clorofila fueron de 0.50 a 1.50 mg m-3 en la zona costera sur (ZCS). El patrón longitudinal de distribución del %SO fue más pronunciado en verano (Fig. 5c), cuando las isolíneas de 100% de saturación se observaron paralelas a la península entre la costa y ~100 km mar adentro, particularmente en la ZCN. En invierno predominaron valores por debajo del 100% de saturación (97%SO; Fig. 5a), lo cual indica que predomina el efecto termodinámico. En primavera (Fig. 5b) destaca el área frente a Ensenada con valores superiores al 110% de saturación e isolíneas paralelas a la península entre la costa y ~100 km mar adentro. En verano permanecieron las condiciones de saturación superiores al 100% frente a la costa de Ensenada, pero las isolíneas paralelas a la península cubrieron un área entre la costa y ~30 km mar adentro.

Figura 4 Climatología estacional (2004-2011) de la concentración de clorofila a (mg m-3) en la superficie del mar en (a) invierno, (b) primavera, (c) verano y (d) otoño.
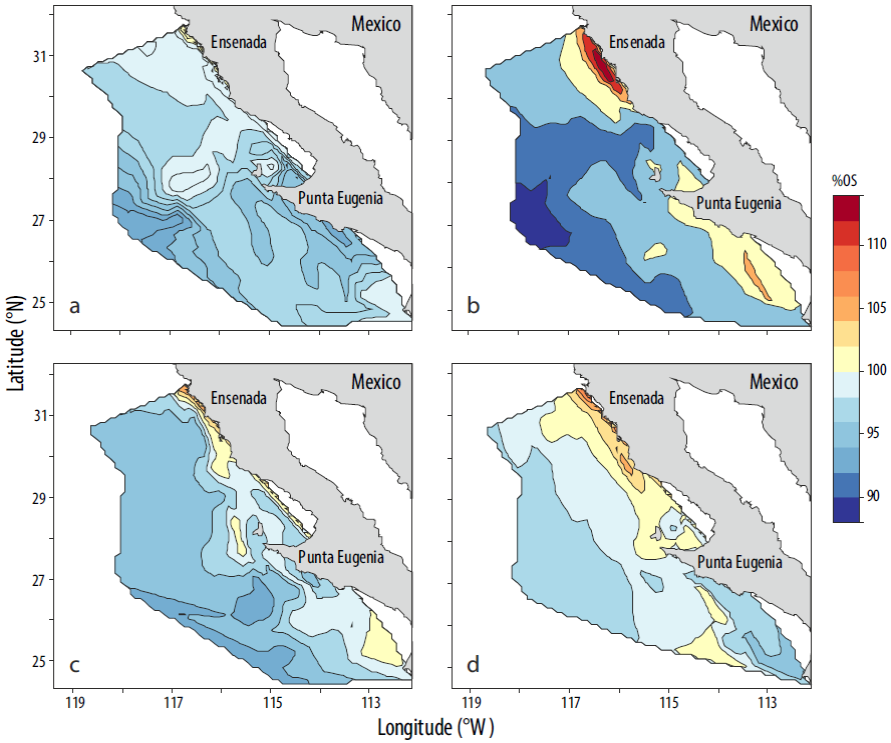
Figura 5 Climatología estacional (2004-2011) del porcentaje de saturación del oxígeno disuelto (%0S) en la superficie del mar en (a) invierno, (b) primavera, (c) verano y (d) otoño.
En general, la pCO2mar presentó un patrón de distribución latitudinal, con valores mayores en la porción sur y menores en la porción norte (Fig. 6); estos valores estuvieron asociados al cambio espacial de la temperatura superficial del mar. La isolínea de 380 μatm perpendicular a la costa se ubicó a ~29°N en otoño (Fig. 6d) y se desplazó hasta ~27°N en invierno (Fig. 6a). En primavera (Fig. 6b), sobresale el área costera al sur de Ensenada con valores de pCO2mar inferiores a 350 μatm e isolíneas paralelas a la península entre la costa y ~80 km mar adentro. En verano (Fig. 6c), la isolínea con valores inferiores a 350 μatm disminuyó su cobertura y cubrió un área entre la costa y ~40 km hacia mar abierto; además, predominaron valores superiores a 400 μatm en una franja oceánica a 29°N, la cual se desplazó hacia el sur del área de estudio. La zona costera al norte de punta Eugenia mostró valores de pCO2 bajos (372 μatm) de manera permanente, lo que indica que fue un posible sumidero de CO2 en el ciclo estacional. La climatología de la pCO2 manifestó un patrón temporal similar al de la temperatura: valores menores en invierno (378 μatm; Fig. 6a) y primavera (388 μatm; Fig. 6b) y valores mayores en verano (410 μatm; Fig. 6c) y otoño (395 μatm; Fig. 6d).
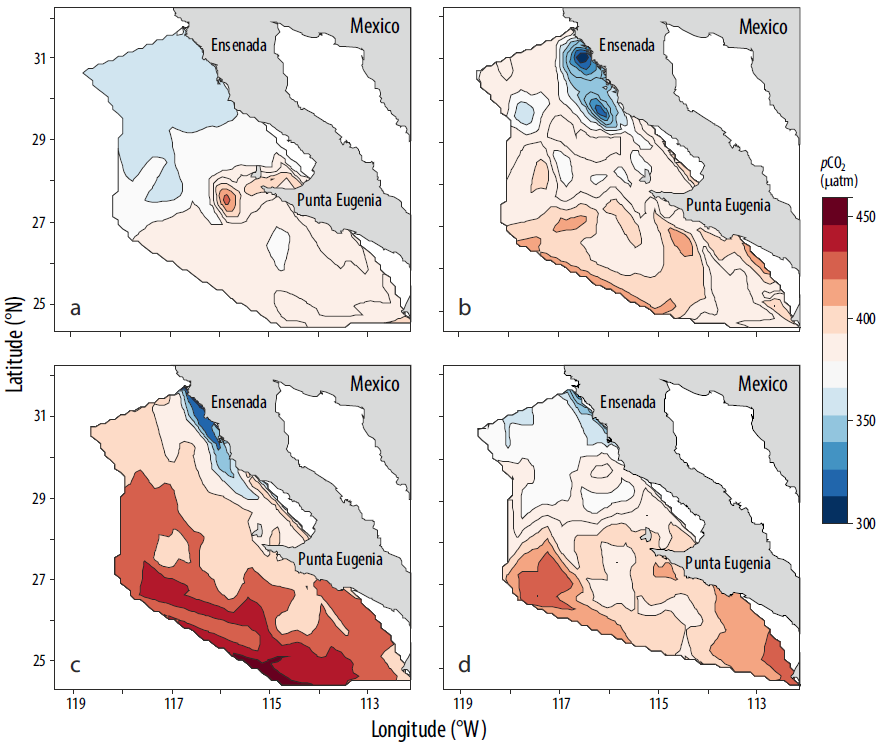
Figura 6 Climatología estacional (2004-2011) de la presión parcial de CO2 (pCO2, μatm) en la superficie del mar en (a) invierno, (b) primavera, (c) verano y (d) otoño.
El FCO2 entre el mar y la atmósfera mostró un patrón de distribución latitudinal, valores mayores en la zona sur y valores menores en la zona norte (Fig. 7), similar al de la distribución de la pCO2 (Fig. 6). En invierno (Fig. 7a), la cobertura del área con valores negativos se extendió hasta ~26°N en la zona oceánica norte (ZON) y hasta los 28°N en la ZCN. Hacia el sur, los valores fueron predominantemente positivos, en promedio 0.85 mmol m-2 d-1. En primavera (Fig. 7b), sobresale el área de la ZCN con valores de FCO2 superiores a -6.00 mmol m-2 d-1 e isolíneas paralelas a la costa que se extendieron ~80 km hacia mar abierto. Desde los ~28.5°N de la ZON se presentó una franja con valores positivos de FCO2 (1.25 mmol m-2 d-1) que se extendió en dirección a punta Eugenia cubriendo prácticamente toda la porción sur del área de estudio. En verano, los flujos negativos disminuyeron su cobertura en la ZCN y se extendieron ~40 km hacia mar abierto (Fig. 7c), y predominaron los flujos positivos. En la Z0N se presentó una franja con valores positivos de FCO2 de 1.20 a 1.80 mmol m-2 d-1 desde los 31°N, y se desplazó hacia el sur cubriendo la zona oceánica sur (ZOS) y la ZCS y alcanzando valores de hasta 6.00 mmol m-2 d-1. En otoño (Fig. 7d), la zona al norte de 28°N presentó flujos negativos de hasta -3.00 mmol m-2 d-1, pero al sur de esta latitud los valores fueron positivos, máximo 3.50 mmol m-2 d-1. La ZCN mostró valores negativos de FCO2 de manera permanente, lo que indica que fue un sumidero de CO2 en el ciclo estacional. La climatología del FCO2 manifestó un patrón temporal similar al de la pCO2: flujos negativos en invierno (-0.26 mmol m-2 d-1; Fig. 7a) y primavera (-0.37 mmol m-2 d-1; Fig. 7b) y flujos positivos en verano (2.33 mmol m-2 d-1; Fig. 7c) y otoño (0.92 mmol m-2 d-1; Fig. 7d).

Figura 7 Climatología estacional (2004-2011) del flujo de CO2 (FCO2, mmol m-2d-1) entre la atmósfera y el mar en (a) invierno, (b) primavera, (c) verano y (d) otoño.
En la serie de tiempo 2004-2011, la temperatura superficial del mar promedio para la porción norte del área de estudio fue de 14.5 a 20.0 °C, y los valores más bajos se presentaron en invierno y primavera y los más altos en verano y otoño (Fig. 8a). A partir del verano de 2007, se observó una tendencia general hacia la disminución de la temperatura en 0.10 °C. La concentración de clorofila fue mayor en primavera; la concentración fue baja (0.42 mg m-3) entre 2004 y mediados de 2007 (Fig. 8b), pero después incrementó. A partir de 2007, la pCO2 promedio mostró la tendencia a disminuir, y los valores estuvieron por debajo del valor atmosférico desde 2009 (Fig. 8c). Esta misma tendencia se presentó en el FCO2, que presentó valores negativos a partir de 2009 (Fig. 8d). Durante la primavera de 2011, la porción norte tuvo el valor máximo como sumidero de carbono de toda la serie de tiempo (-6.00 mmol C m-2 d-1).
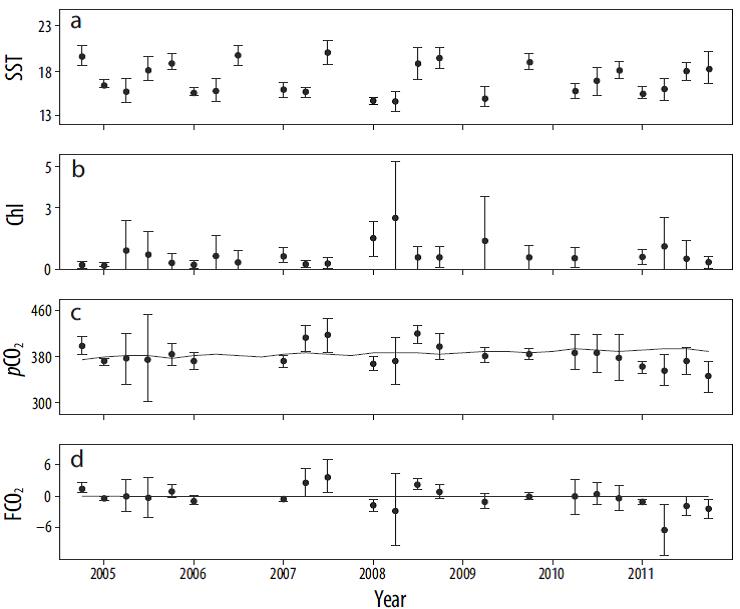
Figura 8 Serie de tiempo (2004-2011) de los promedios de (a) la temperatura (°C), (b) la clorofila (Chl, mg m-3), (c) la presión parcial de CO2 (pCO2, μatm) y (d) el flujo de CO2 (FCO2, mmol m-2 d-1) entre la atmósfera y el mar en la zona norte del área de estudio. Las barras indican el error estándar. La línea continua en (c) indica los valores de pCO2 en la atmósfera. La línea discontinua en (d) indica el equilibrio entre la atmósfera y el océano.
En la porción sur de la región de estudio, la temperatura superficial del mar promedio fue de 15.80 a 23.50 °C; los valores más bajos ocurrieron en invierno y primavera y los más altos en verano y otoño (Fig. 9a). La concentración de clorofila promedio (0.48 mg m-3) fue ligeramente más baja que en la porción norte (0.62 mg m-3), y mostró una tendencia a aumentar a partir del verano de 2006 (Fig. 9b) hasta el final del periodo de estudio. En general, la pCO2 fue superior al valor atmosférico (Fig. 9c) excepto en 2011, cuando fue inferior. A partir de 2007, se observó una tendencia hacia la disminución de la pCO2, la cual se mantuvo hasta el final del periodo de estudio. La porción sur actuó primordialmente como una fuente de CO2 a la atmósfera, con un promedio anual de 1.56 mmol m-2 d-1 (Fig. 9d) y valores próximos al equilibrio en invierno, excepto en 2011 cuando actuó como sumidero.

Figura 9 Serie de tiempo (2004-2011) de los promedios de (a) la temperatura (°C), (b) la clorofila (Chl, mg m-3), (c) la presión parcial de CO2 (pCO2, μatm) y (d) el flujo de CO2 (FCO2, mmol m-2 d-1) entre la atmósfera y el mar en la zona sur del área de estudio (línea IMECOCAL 120 hacia abajo). Las barras indican el error estándar. La línea continua en (c) indica los valores de pCO2 en la atmósfera. La línea discontinua en (d) indica el equilibrio entre la atmósfera y el océano.
Durante el periodo de estudio (2004-2011), la región sur de la corriente de California estuvo bajo la influencia de eventos de El Niño (2004, 2007, 2009-2010) y La Niña (2008, 2009, 2011) (Bjorkstedt et al. 2012) (http://www.cpc.ncep.noaa.gov/products/analysis_monitoring/ensos-tuff/ensoyears.shtml). Para toda la región, la pCO2 promedio en condiciones de El Niño 2004 (Fig. 10) fue 44 μatm mayor que en condiciones de La Niña 2011 (Fig. 10b). En condiciones de El Niño hubo un patrón latitudinal en la distribución espacial de pCO2, donde los valores mayores (414 μatm) se ubicaron en la porción sur y los menores (399 μatm) en la porción norte. En esta última porción se obtuvo un patrón longitudinal en el cambio espacial, y los valores de pCO2 más bajos (394 μatm) se ubicaron en la zona costera. En condiciones de La Niña 2011, la pCO2 promedio fue de 360 μatm, valor inferior al de 393 μatm obtenido para la pCO2atm. En condiciones de El Niño 2004, el océano frente a Baja California actuó como una fuente de CO2 a la atmósfera (promedio de 2.00 mmol m-2 d-1; Fig. 10c), pero actuó como sumidero en condiciones de La Niña 2011 (promedio de 5.30 mmol m-2 d-1; Fig. 10d). Destaca el marcado gradiente latitudinal en la pCO2 y el FCO2 en condiciones de La Niña: valores bajos (altos) de 355 μatm (-6.40 mmol m-2 d-1) en la porción norte y valores altos (bajos) de 366 μatm (-4.00 mmol m-2 d-1) en la porción sur.
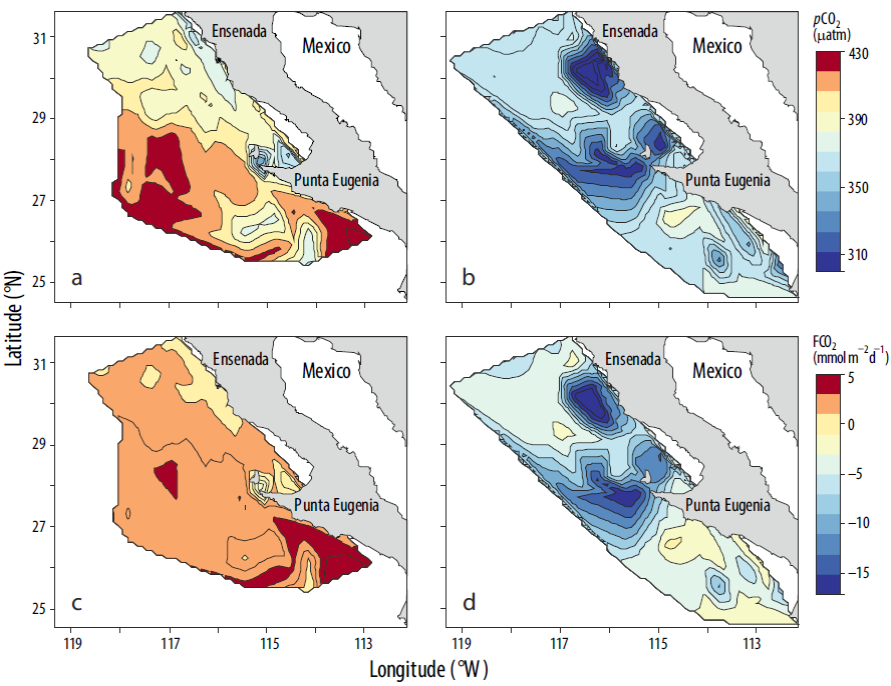
Discusión
Zona oceánica norte (ZON)
En la ZON, los valores bajos de temperatura y salinidad y valores altos de clorofila y %SO en invierno y primavera están asociados con el flujo máximo de la corriente de California hacia el ecuador (Durazo 2015), el cual transporta agua subártica con baja temperatura y salinidad. Los valores bajos de salinidad reflejan la presencia de agua subártica que cubre esta porción de la zona de estudio durante todo el año. Para identificar las variables que regulan la variabilidad de la pCO2, se realizaron correlaciones lineales simples. La variabilidad de la pCO2 en la ZON es regulada mayormente por la temperatura, y la correlación máxima entre ambas variables ocurrió en otoño (r = 0.67, P < 0.05, n = 30). Lo anterior se confirma al calcular el efecto relativo de la temperatura con respecto al efecto de los procesos biológicos en el ciclo estacional, como lo proponen Takahashi et al. (2002). Cuando el efecto de la temperatura menos el efecto biológico (ET - EB) arroja valores positivos, el índice de Takahashi indica que predomina el efecto físico. Por el contrario, si los valores son negativos, existe un predominio del efecto biológico. En la ZON, el ET - EB fue de 7.60, lo que indica un predominio del efecto físico sobre la pCO2 en el ciclo estacional.
Zona costera norte (ZCN)
En la ZCN, las surgencias costeras se presentan todo el año, aunque son más intensas en primavera y verano (Durazo 2015), lo que se manifiesta en los promedios estacionales de temperatura (14.80 °C), alta concentración de clorofila (1.80mgm-3) y valores de la SO superiores al 104%. Las isotermas paralelas a la costa se extienden entre 50 y 100 km mar adentro, particularmente en primavera cuando el viento paralelo a la costa es más intenso (Pérez-Brunius et al. 2007). En esta época, el bombeo de Ekman transporta agua subsuperficial con alta concentración de nutrientes hacia la superficie (Perez-Brunius et al. 2007, Durazo 2015), lo que permite el desarrollo del fitoplancton, el consecuente incremento en los niveles de oxígeno disuelto y, por tanto, la sobresaturación del oxígeno en esta zona de estudio.
La pCO2 mostró un alto coeficiente de correlación lineal inversa con %SO durante las 4 estaciones del año, y el valor máximo correspondió a otoño (r = -0.79, P < 0.05, n = 25). La correlación negativa entre pCO2 y %SO permite identificar los patrones esperados del efecto de los procesos biológicos sobre el consumo y producción de CO2 en el océano. Valores bajos de pCO2 y altos de %SO (por arriba del 100% de saturación) estarían evidenciando la prevalencia de la fotosíntesis (Carrillo et al. 2004), y la situación inversa indicaría una mayor respiración del ecosistema (Schloss et al. 2007). El índice ET - EB para la ZCN fue de -8.26, lo que confirma que la variabilidad de la pCO2 en el ciclo estacional es controlado principalmente por el factor biológico.
Zona oceánica sur (ZOS)
Cuando el núcleo de la corriente de California extiende su dominio hasta la ZOS durante el invierno y la primavera (Durazo y Baumgartner 2002, Durazo 2015), en esta zona se presenta agua con valores bajos de temperatura y salinidad y valores altos de clorofila y %SO. Durante el verano y el otoño, cuando la ZOS está bajo la influencia de agua subtropical, que fluye hacia el norte y transporta agua cálida y salina (Zaitzev et al. 2014, Durazo 2015), se observa un cambio hacia valores altos de temperatura y salinidad y valores bajos de clorofila y %SO. En la ZOS, la pCO2 estuvo correlacionada con la temperatura en verano y otoño. La correlación máxima se presentó en verano (r = 0.85, P < 0.05, n = 30). En invierno y primavera predominaron los factores biológicos; la correlación más alta fue entre la pCO2 y el %SO en primavera (r = -0.62, P < 0.05, n = 29). El índice ET - EB para esta zona fue de 20.79, lo que muestra que en la ZOS predomina el efecto de la temperatura sobre la pCO2 en el ciclo estacional.
Zona costera sur (ZCS)
En la ZCS, el máximo gradiente longitudinal de temperatura se presenta en verano, periodo reportado como de surgencias en esa zona (Durazo et al. 2010). En esta zona los máximos de temperatura y salinidad son ocasionados por agua subtropical superficial, cuya influencia es mayor en verano y otoño (Durazo et al. 2010, Zaitsev et al. 2014, Durazo 2015). En la ZCS predominó el efecto de la temperatura en otoño e invierno, y la correlación máxima fue en otoño (r = 0.64, P < 0.05, n = 25). La relación directa entre la pCO2 y la temperatura indica que el efecto termodinámico derivado de los cambios en la circulación (Hernández-Ayón et al. 2010, Durazo 2015) es uno de los principales factores que regula la pCO2 de la zona. Takahashi et al. (1993) indican que por cada grado que se incrementa la temperatura, la pCO2 aumenta 4.23%; por lo tanto, se esperaría un aumento en la pCO2 de 50 μatm, valor ligeramente superior al calculado de 39 μatm. En primavera y verano, la pCO2 es afectada por los procesos biológicos, debido a que la mayor correlación lineal se presentó con la clorofila en verano (r = -0.59, P < 0.05, n = 32). La relación inversa entre pCO2 y clorofila evidencia, como se ha reportado para otras regiones oceánicas (Mathis et al. 2010), que la pCO2 superficial del mar es modulada por la producción del fitoplancton, lo que pone de relieve el proceso fotosintético en el ciclo del carbono oceánico. El índice ET-EB fue de 18.93, y esto indica que en el ciclo estacional predominó el efecto de la temperatura sobre el efecto biológico en la variabilidad estacional de la pCO2.
Basados en el análisis anterior y en los resultados de los análisis de varianza (ANDEVA) unifactoriales que se realizaron para verificar la existencia de diferencias estadísticamente significativas entre las distintas zonas, es recomendable dividir el área de estudio de la manera que se hace en este trabajo para los estudios de la pCO2 y el FCO2. El ANDEVA muestra que existen diferencias significativas entre zonas, excepto entre la ZCN y la ZON (P = 0.97) en invierno y entre la ZOS y la ZCS (P = 0.11) en otoño.
Flujo de CO 2 entre el mar y la atmósfera
El promedio anual de FCO2 indica que la región de estudio es una fuente de CO2 a la atmósfera (2,043 t). La región cambia de ser fuente en verano y otoño a actuar como sumidero en invierno y primavera. De acuerdo con la clasificación de Paulmier et al. (2008), la zona de estudio se puede caracterizar como fuente débil de CO2 a la atmósfera, ya que emite 0.65 mmol m-2 d-1. Este valor promedio es 52% inferior al valor reportado por De la Cruz-Orozco et al. (2010) para la misma área de estudio, que fue calculado con datos recolectados en cruceros realizados entre 2004 y 2005, periodo bajo la influencia de El Niño y la intrusión anómala de un gran volumen de agua subártica (Espinoza-Carreón et al. 2015). Hernández-Ayón et al. (2010) estimaron un flujo de carbono de 2.60 mmol m-2 d-1 para la región oceánica frente a Baja California con datos recolectados entre 1993 y 2001, un valor 3 veces más alto que el calculado en el presente estudio. Hales et al. (2012), con base en información derivada de modelos, reportaron que la zona es una fuente de CO2 y emite 1.80 mmol m-2 d-1. La diferencia con nuestros resultados puede deberse a que Hales et al. (2012) ajustaron sus modelos para obtener la pCO2 con datos recolectados entre 1997 y 2005, además de que calcularon el coeficiente de solubilidad con datos de temperatura satelital y no utilizaron la misma parametrización para el cálculo del coeficiente de transferencia.
La ZCN es un sumidero permanente de CO2 ya que el FCO2 promedio estacional fue de -5.20 mmol m-2 d-1 (715 t). La ZON es un sumidero de otoño a primavera y actúa como fuente en verano (360 t) (Fig. 7). La porción sur es una fuente de CO2 a la atmósfera durante todo el año (1,252 t), con los valores más bajos en invierno y primavera. El cambio estacional de fuente a sumidero en esta región ha sido descrito por otros autores; sin embargo, De la Cruz-Orozco et al. (2010) usaron datos obtenidos cuando el sistema estuvo bajo la influencia de la intrusión anómala de agua subártica y bajo condiciones de El Niño, Hernández-Ayón et al. (2010) utilizaron datos mayormente de la porción oceánica y Hales et al. (2012) mostraron información derivada de modelos. Por lo tanto, los resultados del presente trabajo se pueden considerar, hasta ahora, los más robustos.
En invierno y primavera, cuando la zona norte es sumidero de CO2 y la zona sur presenta los valores más bajos de FCO2, toda el área de estudio se encuentra bajo la influencia de agua subártica (Durazo 2015), que transporta agua fría y con alta concentración de oxígeno disuelto. Esta afirmación se apoya en la distribución espacial de la temperatura, cuyo valor promedio es de 16.50 °C en invierno y de 16.30 °C en primavera (Fig. 2a, b). Por otro lado, el %SO presenta los valores más altos en invierno y primavera (97.14% y 97.70%, respectivamente) (Fig. 5a, b).
La ZOS y la ZCS son fuentes permanentes de CO2 a la atmósfera (2.05 y 0.10 mmol m-2 d-1, respectivamente). Los valores positivos de FCO2 más altos se presentaron en verano y otoño, cuando la corriente de California se debilita (Durazo 2015) y en la zona sur ingresa agua de origen subtropical (Zaitsev et al. 2014) cálida, más salada y con baja concentración de oxígeno disuelto. Este cambio estacional en la circulación durante el verano y otoño da como resultado mayor temperatura (Fig. 2c, d) y salinidad (Fig. 3a, b), y menor %SO (Fig. 5b, c) en la zona sur.
p CO 2 y FCO 2 entre el mar y la atmósfera en condiciones de El Niño y La Niña
El área de estudio fue fuente de CO2 (1.90 mmol m-2 d-1) a la atmósfera durante El Niño (otoño de 2004) y actuó como sumidero (-5.30 mmol m-2 d-1) durante La Niña (primavera de 2011). Durante los eventos de El Niño aumenta la estratificación del océano, la nutriclina se profundiza (Huyer y Smith 1985) y la biomasa y producción del fitoplancton disminuyen (Hernández-De la Torre et al. 2004). El aumento en la temperatura genera mayor fugacidad del CO2, en el orden de 4.23% por cada grado centígrado (Takahashi et al. 1993). Además, el aumento de la estratificación de la columna de agua limita el ascenso de agua rica en nutrientes, lo cual genera condiciones poco propicias para el desarrollo del fitoplancton, y disminuye la utilización biológica del carbono inorgánico disuelto. Durante los eventos de La Niña se presentan situaciones contrarias; las condiciones son favorables a las surgencias y el agua rica en nutrientes transportada a la superficie favorece el crecimiento del fitoplancton, que asimila el CO2, y, por consiguiente, disminuye la pCO2 superficial del océano. La concentración de clorofila promedio fue 0.21 mg m-3 para toda el área de estudio durante el evento de El Niño 2004, pero incrementó a 1.12 mg m-3 durante el evento de La Niña 2011 (Figs. 8b, 9b). El %SO fue menor en condiciones de El Niño que en condiciones de La Niña (99.33% y 101.70%, respectivamente). En cuanto a la pCO2 y el FCO2 entre condiciones de El Niño y condiciones de La Niña, los resultados del ANOVA unifactorial indicaron que las diferencias fueron significativas (P < 0.05).
El factor termodinámico también es relevante para explicar las diferencias observadas durante las diferentes fases de ENOS. La temperatura superficial promedio durante El Niño fue de 20.92 °C y durante La Niña fue de 16.03 °C (Figs. 8a, 9a). De acuerdo con Takahashi et al. (1993), esto implica que el solo efecto termodinámico aumentaría la pCO2 en el orden 70 μatm en condiciones de El Niño con respecto a La Niña.
Impacto en el ambiente epipelágico
La explicación de la respuesta del fitoplancton, principal responsable del hundimiento de carbono biogénico hacia mayores profundidades, al incremento del CO2 en el océano hasta ahora ha sido contradictoria. Por un lado, se ha reportado que el aumento del CO2 del mar disminuye la calcificación en organismos como corales, foraminíferos y cocolitofóridos (Beaufort et al. 2011) debido a que este incremento genera agua corrosiva (disminución del pH), cuya presencia se ha comprobado en la región sureña de la corriente de California (Feely et al. 2008, Alin et al. 2012). Sin embargo, también se han reportado organismos del fitoplancton con alta tasa de calcificación, como el cocolitofórido Emiliania huxleyi (Beaufort et al. 2011, Ribero-Calle et al. 2015), que parece poseer gran capacidad adaptativa (Lohbeck et al. 2012), en agua con bajo pH (7.62). Estos son algunos ejemplos que demuestran la gran complejidad que actualmente existe para explicar la respuesta del ecosistema epipelágico al incremento del CO2. Es necesario continuar con las mediciones in situ del CO2 y contar con bases sólidas para estimar el efecto del aumento a largo plazo en el carbono inorgánico disuelto del océano sobre los organismos.

