










Serviços Personalizados
Journal
Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkCiencias marinas
versão impressa ISSN 0185-3880
Cienc. mar vol.42 no.2 Ensenada Jun. 2016
https://doi.org/10.7773/cm.v42i2.2592
Artículos
Cambios temporales de la biomasa y distribución de los mantos de Sargassum en la costa sureste de la península de Baja California
1 Centro Interdisciplinario de Ciencias Marinas, Instituto Politécnico Nacional (CICIMAR-IPN), Avenida Instituto Politécnico Nacional s/n, La Paz 23096, Baja California Sur, México.
2 Centro de Investigaciones Biológicas del Noroeste (CIBNOR), Avenida Instituto Politécnico Nacional #195, La Paz 23096, Baja California Sur, México.
Se describen y comparan con estimaciones previas los cambios en la distribución, superficie, biomasa y cosecha de mantos de Sargassum (Phaeophyceae) a lo largo de la costa sureste de la península de Baja California (México). También, la biomasa de Sargassum se correlacionó con la latitud. Se determinaron las especies y se estimó la superficie, biomasa, cosecha y estructura de tallas de 72 mantos de Sargassum en 3 zonas en 2006. La extensión de cada manto fue determinada usando GPS. El perímetro de cada manto fue dibujado y su superficie fue calculada. La biomasa promedio fue estimada usando cuadrantes que fueron distribuidos al azar sobre los mantos. Se registró la biomasa promedio más alta en Mulegé, encontrándose diferencias significativas con respecto a las otras 2 zonas de estudio (bahías Concepción y La Paz). En bahía Concepción se registró el manto más grande y la mayor superficie y cosecha. Los cambios en la distribución, superficie, biomasa y cosecha de los mantos de Sargassum a lo largo de la costa sureste de la península de Baja California fueron comparados con diferentes estadios fenológicos de Sargassum, variaciones en la disponibilidad de sustrato rocoso y características oceanográficas. Estos aspectos deben ser considerados para mantener una explotación sustentable de este recurso potencial.
Palabras clave: biomasa; golfo de California; Sargassum; cosecha; cambios estacionales
Changes in the distribution, surface canopy, biomass, and standing crop of Sargassum beds (Phaeophyceae) along the southeastern coast of the Baja California Peninsula (Mexico) are described and compared with previous estimates. Also, Sargassum biomass was correlated with latitude. Species were identified and surface canopy, biomass, standing crop, and length structure were estimated for 72 Sargassum beds in 3 localities in 2006. The extent of each bed was determined using GPS. The perimeter of each bed was drawn and its canopy area was calculated. The average biomass was estimated, using quadrats that were randomly distributed over the beds. The highest average biomass was found at Mulegé, with significant differences in the other 2 study areas (Concepción and La Paz bays). The largest bed, surface canopy area, and standing crop were recorded in Concepción Bay. Coverage area, location, and seasonal changes in surface canopy, standing crop, and biomass of Sargassum beds along the southeastern coast of the Baja California Peninsula were compared with the different phenological stages of Sargassum thalli, variations in the available rocky sea floor, and oceanographic characteristics. These aspects must be considered to maintain sustainable exploitation of this potential resource.
Key words: biomass; Gulf of California; Sargassum; standing crop; seasonal changes
Introducción
El género Sargassum C Agardh (Phaeophyceae, Fucales, Sargassaceae) presenta una amplia distribución, especialmente en regiones tropicales y subtropicales (Noiraksar et al. 2014), incluyendo el golfo de California (Pacheco-Ruíz et al. 1998). En el golfo, las especies de Sargassum constituyen la mayoría de la biomasa de macroalgas carnosas, y proporcionan hábitat para diversas comunidades de algas epifíticas y una cubierta para algas, invertebrados y peces del sotobosque (Norris 2010).
Las especies de Sargassum del golfo de California podrían servir como fuente de alginato (Rodríguez-Montesinos et al. 2008) y podrían usarse en fertilizantes para la agricultura, en forraje para cabras y ovejas, en dietas balanceadas para pollos y camarones y en la alimentación humana (Marín-Álvarez et al. 2003; Casas-Valdez et al. 2006a, 2006b; Casas-Valdez 2009). Los extractos de Sargassum presentan actividad antimicrobiana contra organismos patógenos, por lo que tienen potencial para la industria farmacéutica (Muñoz-Ochoa et al. 2010). Estudios realizados con Sargassum sinicola han identificado algunos usos potenciales de alginato como materia prima para la inmovilización de células en el tratamiento de aguas residuales y para promover el crecimiento de plantas (Yabur et al. 2007), así como su capacidad para absorber residuos de metales pesados de la industria minera (Huerta-Díaz et al. 2007, Patrón-Prado et al. 2010). A la fecha, los mantos de Sargassum no han sido cosechados en el golfo de California para propósitos comerciales (Casas-Valdez 2009). Para poder realizar una correcta planificación comercial, es necesario estimar la biomasa (Andrefouet et al. 2004).
Las investigaciones sobre Sargassum en el golfo de California indican un patrón estacional de crecimiento y abundancia. En general, la máxima tasa de crecimiento y la máxima longitud del talo se registran en primavera, como ha sido documentado para S. sinicola (Núñez-López y Casas-Valdez 1997) y S. lapazeanum (Rivera y Scrosati 2006). En primavera, las tasas de crecimiento más altas coinciden con la máxima biomasa registrada para las especies en la costa este de la península de Baja California: S. sinicola (Espinoza- Avalos 1990), S. horridum (Muñeton y Hernández-Carmona 1993) y S. lapazeanum (Rivera y Scrosati 2006) en bahía de La Paz; S. sinicola, S. herporhizum y S. lapazeanum (Núñez-López y Casas-Valdez 1996) en bahía Concepción; y S. herporhizum, S. johnstonii y S. sinicola de Mulegé a San Luis Gonzaga (Pacheco-Ruíz et al. 1998). El crecimiento, la longitud del talo y la biomasa de Sargassum son mayores cuando la temperatura del agua es relativamente fría, lo que sucede en primavera, y el crecimiento es menor a finales de verano (Espinoza-Avalos 1993).
Se ha estimado la biomasa total de los mantos de Sargassum (en toneladas húmedas) en varios sitios a lo largo de la costa este de la península: bahía de La Paz (18,900 ± 928), bahía Concepción (7,250 ± 530) y de Mulegé a San Luis Gonzaga (154,559 ± 16,139) (Hernández-Carmona et al. 1990, Casas-Valdez et al. 1993, Pacheco-Ruíz et al. 1998); sin embargo, según nuestro conocimiento, éstas son las últimas estimaciones que se han publicado y se necesitan estimaciones actuales ya que la distribución y cobertura podrían haber cambiado en las últimas 2 décadas.
Para actualizar la información, se estudiaron y compararon con estudios previos la ubicación, área superficial, biomasa y cosecha de mantos de Sargassum en la costa sureste de la península de Baja California (México). También se examinó si había una relación entre la biomasa y la latitud. Para evaluar el potencial para una cosecha sustentable de Sargassum, es de suma importancia contar con una nueva base de datos de su cobertura y abundancia estacional en esta región.
Materiales y métodos
Área de estudio
Los muestreos se realizaron entre La Ventana (sur de la bahía de La Paz; 24º02´N, 109º59´W) y La Salina (27º52´N, 112º46´W) (Fig. 1). Se estudiaron 3 zonas: (I) Mulegé (26º56´N, 111º58´W a 27º59´N, 112º43´W), (II) bahía Concepción (26º52´N, 111º54′W a 26º35´N, 111º43W) y (III) bahía de La Paz (24º45´N, 110º28´W a 24º08´N, 110º24´W). Estos sitios fueron seleccionados por contar con datos de los mantos de Sargassum. Para que las comparaciones fueran más útiles, la ubicación específica de cada manto dentro de cada zona fue igual o muy similar a los sitios de estudios previos. Los muestreos se realizaron en primavera ya que los estudios previos también fueron realizados en primavera.
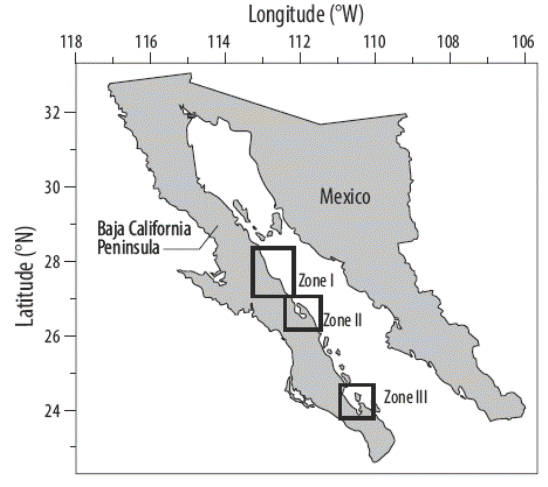
Figura 1 Área de estudio, costa sureste de la península de Baja California (México): Mulegé (zona I), bahía Concepción (zona II) y bahía de La Paz (zona III).
A lo largo de la costa de la península de Baja California, hay zonas conspícuas donde crecen los mantos de Sargassum, cada una aislada de la zona vecina. Estos mantos albergan una comunidad de especies, donde cada comunidad puede crecer hasta 25 m. Las hojas superiores llegan a la superficie y forman una densa bóveda arbórea. Nuestro estudio incluyó 23 mantos en la zona I, 22 mantos en la zona II y 27 mantos en la zona III.
Proporción de especies y longitud
A lo largo de esta costa se encuentran varias especies de Sargassum. La proporción de cada especie se determinó mediante un muestreo aleatorio en cada manto. Se distribuyeron 3 cuadrantes al azar. Todos los individuos dentro de un cuadrante fueron recolectados y subsecuentemente identificados a nivel de especie con base en 2 guías de identificación (Setchell y Gardner 1924, Dawson 1944). Se midió la longitud de los especímenes de Sargassum; 50 talos de cada especie fueron medidos desde el órgano de fijación hasta el ápice para obtener la longitud promedio en las 3 zonas. La longitud está directamente relacionada con la biomasa, como lo describen Rivera y Scrosati (2006).
Área superficial, biomasa y cosecha
Los mantos de Sargassum fueron clasificados como pequeños (1 a 500 m), medianos (501 a 1,500 m) o grandes (>1,501 m) según su longitud (Hernández-Carmona et al. 1990, Casas-Valdez et al. 1993). Los mantos de Sargassum yacen paralelos a la línea de costa. La longitud del manto es la distancia lineal de cada manto paralelo a la costa, y la anchura es la distancia lineal desde la parte más cercana a la costa hasta la parte más lejana de la costa, perpendicular a la línea de costa. Para no sobrepasar un nivel de error del 10%, se midió la anchura de los mantos cada 50 m en el caso de los pequeños, cada 100 m en el caso de los medianos y cada 200 m en el caso de los grandes. Los perímetros y el área superficial de los mantos fueron calculados con el programa ArcView 3.2 (ESRI 2008).
Para determinar la biomasa algal promedio, se usaron los cuadrantes como unidades de muestreo. Todos los talos de Sargassum en cada cuadrante fueron recolectados a mano y pesados (peso húmedo [PH]) al 0.1 kg más cercano. Se usaron 10 cuadrantes para los mantos pequeños, distribuidos al azar en todo el área superficial, usando una tabla de números aleatorios; se usaron 15 cuadrantes para los mantos medianos y 20 cuadrantes para los mantos grandes (Hernández-Carmona et al. 1990, Casas-Valdez et al. 1993). Para determinar la cosecha de cada manto, se multiplicó su biomasa promedio por su área superficial total. Para calcular la cosecha de cada zona, se sumó la cosecha de los mantos en cada zona.
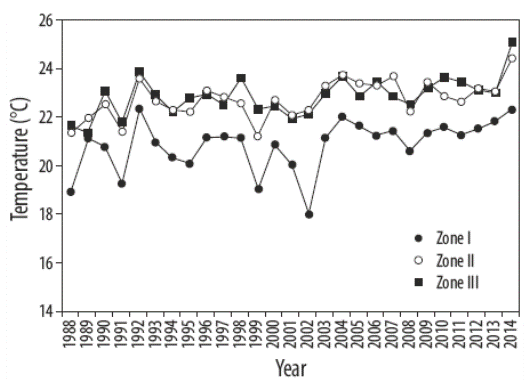
Para todos los mantos, se registraron el tipo de sustrato (arena, roca o canto rodado) y la profundidad de cada estación. A partir de estas observaciones se construyó una tipología del manto algal. La temperatura superficial del mar se obtuvo de datos satelitales para 1988 a 2014 (http://oceanwatch.pifsc.noaa.gov/). La relación de la biomasa de Sargassum con la latitud y la temperatura del mar se determinó mediante un análisis de correlación lineal (Cohen et al. 2003).
Cambios espaciales y temporales
Se realizó un análisis de varianza para determinar si existían diferencias significativas entre las 3 zonas en cuanto a la biomasa, área superficial, cosecha y longitud promedio de las especies de Sargassum. Las diferencias entre los valores medios fueron analizadas mediante la prueba de Tukey, con el paquete SAS 6.12, estableciendo un valor de significancia de P < 0.05. Los datos obtenidos en la primavera de 2006 para el área superficial, la biomasa y la cosecha de los mantos de Sargassum fueron comparados con los datos estimados para la primavera de 1988 (Hernández-Carmona et al. 1990), 1991 (Casas-Valdez et al. 1993) y 1995 (Pacheco-Ruíz et al. 1998). El área de estudio de Pacheco- Ruíz et al. (1998) se extendió de 26º55´N, 111º58´W a 27º57´N, 111º45´W, el de Casas-Valdez et al. (1993) de 26º55´N, 111º54´W a 26º35´N, 111º45´W, y el de Hernández-Carmona et al. (1990) de 24º22´N, 110º12´W a 24º08´N, 110º24´W. Los muestreos se realizaron en primavera ya que Sargassum crece más rápido y alcanza su máxima longitud y biomasa durante esta temporada, como ya se mencionó. Mediante un análisis de varianza se determinó si existían diferencias en la biomasa entre años.
Resultados
Cambios espaciales
Se identificaron 2 especies de la familia Sargassaceae: Sargassum sinicola subsp. camouii (EY Dawson) JN Norris & Yensen y S. lapazeanum Setchell & NL Gardner. Ambos fueron encontrados en la bahía de La Paz (zona III) con una proporción de talos de 70% y 30%, respectivamente. En las zonas I y II se encontró solamente S. sinicola subsp. camouii.
Se registró la menor (P = 0.00002) longitud promedio de los talos de S. sinicola subsp. camouii en la bahía de La Paz (0.9-2.1 m) y la mayor (P = 0.00002) en Mulegé (1.5-19.0 m). En la zona II, la longitud fue de 0.1 a 3.0 m (n = 2,917). La longitud de los talos de S. lapazeanum en la zona III (0.9-1.3 m) fue menor (n = 1,947; P = 0.00002) que la de S. sinicola subsp. camouii.
Los mantos de Sargassum se encontraron en aguas someras (0.30-4.0 m) en la zona III, a profundidades intermedias (0.5-5.0 m) en la zona II y en un amplio intervalo de profundidades (1.0-25 m) en la zona I. Crecen sobre piedra, rocas y cantos rodados en costas expuestas y protegidas. No se observaron mantos de Sargassum creciendo sobre fondos arenosos.
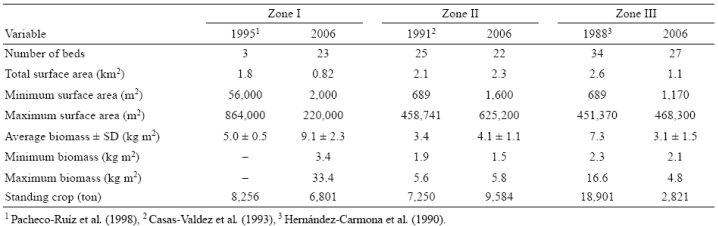
En 2006, se encontraron 72 mantos de Sargassum en la costa sureste de la península. Cubrieron un área superficial de 4.22 km2, con una cosecha estimada de 19,206 t. Se encontraron 23 mantos en la zona I, 22 en la zona II y 27 en la zona III (Fig. 1, Tabla 1). La biomasa promedio más alta se registró en Mulegé (zona I), observándose diferencias significativas (P = 0.000001) con respecto a las otras 2 zonas de estudio (Tabla 1). El área superficial y la cosecha fueron mayores en bahía Concepción (zona II).
Tabla 1 Cambios temporales del número de mantos, el área superficial, la biomasa y la cosecha de Sargassum spp. en Mulegé (zona I), Bahía Concepción (zona II) y bahía de La Paz (zona III).
En general, la biomasa promedio de los mantos de Sargassum incrementó con la latitud (R2 = 0.59, P = 0.0592) (Fig. 2), esto es, está inversamente relacionada con la temperatura del agua. La temperatura promedio en primavera decreció a mayor latitud (Fig. 3), mientras que la biomasa promedio de los mantos decreció a mayor temperatura (R2 = 0.52, P = 0.0000). Se observó una correlación entre la longitud de S. sinicola subsp. camouii y la profundidad de los mantos (R2 = 0.71, P = 0.0000).
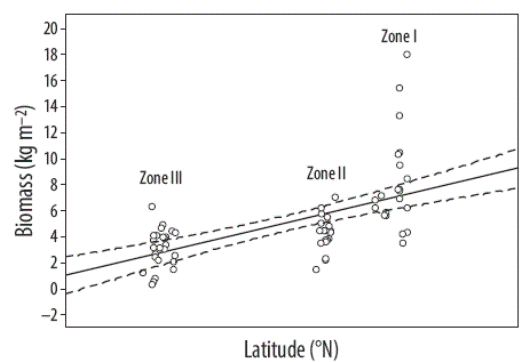
Figura 2 Relación entre la biomasa de Sargassum y la latitud en la costa sureste de la península de Baja California (las líneas discontinuas son residuales; la línea sólida es la línea ajustada).
Cambios temporales
Zona I
En 1995, Pacheco-Ruíz et al. (1998) encontraron 3 mantos grandes y el área superficial del manto más pequeño fue de 56,000 m2. En 2006, encontramos 23 mantos más pequeños y el área superficial varió de 1,450 a 220,000 m2. El área superficial de los mantos de Sargassum en 1995 fue 45% más grande que en 2006 (1.8 y 0.82 km2, respectivamente). La biomasa promedio en 1995 (5 kg m-2 PH) fue 55% menor que en 2006 (9.1 kg m-2 PH), pero la cosecha en 1995 (8,256 t PH) fue 18% mayor que en 2006 (6,801 t PH) (Tabla 1).
Zona II
En 1991, Casas-Valdez et al. (1993) encontraron 25 mantos, mientras que en 2006 encontramos 22. Los mantos de 3 sitios (Buenaventura Norte, Cajede y Tasajera) estuvieron ausentes en 2006. El área superficial de los mantos varió considerablemente entre 1991 y 2006: la superficie de algunos mantos permaneció casi igual, como la de los mantos 29 (27,374 a 27,200 m2) y 24 (49,070 a 46,700 m2); la superficie de algunos aumentó mucho, como la de los mantos 45 (65,446 a 625,200 m2), 37 (689 a 137,750 m2) y 25 (9,609 a 382,800 m2); y la superficie de algunos disminuyó mucho, como la de los mantos 43 (458,741 a 14,950 m2), 39 (145,847 a 7,200 m2) y 31 (108,062 a 1,600 m2).
En 1991, la biomasa varió de 1.9 kg m-2 PH en el manto 31 a 5.6 kg m-2 PH en el manto 45. En 2006, la biomasa varió de 1.5 kg m-2 PH en el manto 24 a 5.8 kg m-2 PH en el manto 27. La biomasa promedio fue significativamente mayor (P = 0.00244) en 2006 (4.1 kg m-2 PH) que en 1991 (3.4 kg m-2 PH). La biomasa en 2006 fue mayor que en 1991 en todos los mantos, excepto en los mantos 45, 44 y 24. La cosecha fue 24% mayor en 2006 (9,584 t PH) que en 1991 (7,250 t PH) (Tabla 1).
Zona III
En 1988, Hernández-Carmona et al. (1990) encontraron 34 mantos, mientras que en 2006 encontramos 27. La desaparición de mantos grandes en Ensenada Verde-Puertecitos (451,370 m2), Sur de Las Ánimas (60,653 m2) y Ensenada San Gabriel en la isla Espíritu Santo (162,021 m2) es importante. El área superficial total de los mantos de Sargassum en 1988 fue 2.6 km2, más de 2 veces la superficie medida en 2006 (1.1 km2) (Tabla 1). El área superficial de todos los mantos fue mayor en 1988 que en 2006, excepto el de los mantos 68 (1988 = 24,564 m2; 2006 = 43,500 m2) y 57 (1988 = 7,436 m2; 2006 = 50,900 m2).
La biomasa promedio en 1988 (7.3 kg m-2 PH) fue significativamente mayor (58%) que en 2006 (3.1 kg m-2 PH) (P = 0.000001). La biomasa para todos los mantos fue menor en 2006 que en 1988 (Tabla 1). Los mantos que presentaron las mayores diferencias fueron el 57 (4.1 kg m-2 PH en 2006 y 16.6 kg m-2 PH en 1988, una reducción de 61%) y el 47 (3.2 kg m-2 PH en 2006 y 11.2 kg m-2 PH en 1988, una reducción de 71%). La cosecha de Sargassum en 2006 (2,821 t PH) fue 85% menor que en 1988 (18,900 t PH).
Discusión
Comparación de tallas
Sargassum sinicola subsp. camouii fue encontrada en todas las estaciones de muestreo, independientemente de la profundidad, y fue la especie dominante en biomasa. Esto se puede atribuir a su estrategia de reproducción y su persistencia durante largos periodos de tiempo (Espinoza-Avalos 1990). Sargassum lapazeanum sólo fue abundante en aguas someras con fondos rocosos en la zona III.
La longitud promedio más alta de S. sinicola subsp. camouii se registró en la zona I, lo cual se puede atribuir al mayor contenido de nutrientes en aguas del norte (Halfar et al. 2004, 2006) y a la mayor profundidad del fondo marino. Pacheco-Ruíz et al. (1998) registraron una longitud promedio de 1.4 ± 0.4 m para esta zona. En nuestro estudio, se observó S. sinicola subsp. camouii a profundidades de 1 a 25 m. La longitud promedio varió de 1.3 a 19 m (media: 5.2 ± 3.1 m) y aumentó gradualmente a mayor distancia de la costa (Yeong et al. 2012). Ésta es la primera vez que se ha documentado una longitud tan larga en esta zona. Una longitud similar fue registrada para S. johnstonii en la costa noreste de la península (Pacheco-Ruíz et al. 1998). La longitud promedio de S. sinicola subsp. camouii en las 3 zonas fue mayor que la longitud máxima registrada para la misma especie en bahía Magdalena (40 cm; Sánchez-Rodríguez y Cervantes-Duarte 1999).
En bahía Concepción, en la primavera de 1991, la longitud promedio de S. sinicola subsp. camouii fue de 2.1 m y la longitud máxima fue de 7 m (Casas-Valdez et al. 1993, Núñez-López y Casas-Valdez 1997). En contraste, en 2006, la longitud promedio fue de 0.86 m. En 2006, los mantos se localizaron más cerca de la costa, a profundidades de 0.5 a 5 m; en 1991, los mantos se localizaron a profundidades de 0.5 a 15 m (Casas-Valdez et al. 1993). La profundidad a la cual crece Sargassum depende de la presencia de estrato duro (Staehr et al. 2008, Tweedley et al. 2008).
La longitud de S. lapazeanum fue menor que la de S. sinicola en bahía Concepción en 1991 (Núñez-López y Casas-Valdez 1997) y en el manto 74 en la bahía de La Paz en 2000 (Rivera y Scrosati 2006). Las diferencias en longitud son en gran medida determinadas por características genotípicas (Núñez-López y Casas-Valdez 1997).
La biomasa promedio de Sargassum fue menor en las zonas II y III en 2006 que en la costa noreste de la península (8.7-22 kg m-2 PH en Pacheco-Ruíz et al. 1998), que es un área templada con menor temperatura superficial del mar y mayor contenido de nutrientes (Espinoza-Avalos 1993). Las zonas II y III fueron similares a otras áreas subtropicales como bahía Magdalena (3.8 kg m-2 PH; Sánchez-Rodríguez y Cervantes-Duarte 1999). La zona I en 2006 es consistente con 2 áreas sureñas (8 y 9) estudiadas por Pacheco-Ruíz et al. (1998) en 1995. La biomasa promedio fue de 9.1 kg m-2 PH, mayor que la estimación anterior (5.0 kg m-2 PH; Pacheco-Ruíz et al. 1998). Esto es entre estimaciones de regiones subtropicales y templadas. La biomasa en la zona I se relaciona con su ubicación transicional entre las zonas costeras de las partes norte y central de la península (Espinoza-Avalos 1993). Esta área es dominada por corrientes de surgencia en el canal de Ballenas, y las aguas son más frías y más ricas en nutrientes que en la costa sureste (Álvarez-Borrego 1983, Álvarez-Molina et al. 2013). Según Yeong et al. (2012), la biomasa de las especies de Sargassum aumenta gradualmente hacia fuera de la costa; en esta zona, los talos de Sargassum pueden alcanzar 25 m.
Halfar et al. (2004, 2006) midieron clorofila a como indicador de nutrientes y temperatura en la costa este de la península (a 23ºN, 25ºN, 27ºN y 29ºN) para relacionar estos factores con la producción de carbonato. La clorofila a aumenta hacia el norte: 0.3, 0.6, 1.6 y 2.2 mg m-3 a 23ºN, 25ºN, 27ºN y 29ºN, respectivamente, lo que puede explicar el aumento en biomasa de Sargassum al aumentar la latitud.
La cosecha estimada en 2006 (este estudio) y en 1988 por Hernández-Carmona et al. (1990) y 1991 por Casas-Valdez et al. (1993) fue menor que la cosecha calculada por Pacheco- Ruíz et al. (1998) en la costa noreste de la península, ya que en esta zona localizada más al norte la temperatura del agua es menor y el contenido de nutrientes es mayor (Espinoza-Avalos 1993) que más al sur (Halfar et al. 2004, 2006).
Comparaciones temporales
La diferencia en biomasa en la zona III entre 1988 y 2006 podría ser resultado de los cambios fenológicos en los mantos de Sargassum producidos por variaciones de factores ambientales o una disminución de sustrato duro. Estudios futuros podrían indicar cuales son los factores que producen estos cambios. En 2006 se registró la menor cosecha y biomasa promedio del estudio. Las diferencias en la cosecha entre 1988 y 2006 probablemente están relacionadas con la desaparición, en 2006, de algunos mantos grandes.
Núñez-López y Casas-Valdez (1996) estudiaron los cambios fenológicos de Sargassum en bahía Concepción e identificaron 8 fases de desarrollo (plántula, juvenil, joven A, joven B, maduro, senescente A, senescente B, senescente C). Encontraron que existen diferencias geográficas y estacionales en las etapas fenológicas de Sargassum. Por ejemplo, en la primavera de 1991, el mayor porcentaje de talos de Sargassum en el manto 45 eran jóvenes A (receptáculos en desarrollo) y B (receptáculos desarrollados), mientras que el manto 32 contenía plantas maduras y el manto 26 contenía principalmente plantas senescentes B. En la primavera de 1992, el manto 45 contenía principalmente plantas jóvenes (A y B), el manto 32 contenía plantas jóvenes (A) y en el manto 26 todas las plantas eran juveniles. Estas 8 fases fenológicas están directamente relacionadas con la cantidad de frondas y, por lo tanto, con la biomasa de Sargassum (Núñez- López y Casas-Valdez 1997). Las diferencias en la biomasa de los mantos de Sargassum en bahía Concepción entre 1991 y 2006, y entre los diferentes mantos en cada año, puede ser resultado de las diferencias en las fases de desarrollo de los talos de Sargassum en un tiempo específico.
El principal factor que afecta la ubicación de Sargassum es el sustrato rocoso (Espinoza-Avalos 1993). Encontramos que había una relación entre la longitud del manto y la superficie del sustrato duro. Los mantos más grandes se encuentran en sitios donde una mayor área está cubierta por piedras, rocas y/o cantos rodados (Casas-Valdez et al. 1993). El tamaño de las playas y los sustratos duros varía porque están expuestos a una dinámica estacional e interanual, las primeras principalmente debido a la energía del oleaje (más intenso en invierno) y los segundos debido a tormentas tropicales y huracanes. En las 3 zonas, no se han observado modificaciones costeras importantes o urbanización cerca de los mantos.
Nuestros resultados muestran que la cosecha de Sargassum en las 3 zonas había variado en 2006 con respecto a los años anteriores. Es importante tomar esta variación en cuenta si se quiere lograr una explotación sustentable de este recurso abundante, el cual puede ser usado como fuente de alginato en muchos productos, dietas balanceadas, forraje, etc. Recomendamos que se realice un monitoreo periódico de los mantos para determinar si el recurso es sustentable. La alta variación del área superficial, la biomasa y la cosecha de los mantos de Sargassum no se debe a actividades antropogénicas ya que nunca se han cosechado los mantos en la costa este de la península de Baja California.
References
Álvarez-Borrego S. 1983. Gulf of California. In: Ketchum BH (ed.), Estuaries and Enclosed Seas. Elsevier, Amsterdam, pp. 427-449. [ Links ]
Álvarez-Molina LL, Álvarez-Borrego S, Lara-Lara JR, Marinone SG. 2013. Annual and semiannual variations of phytoplankton biomass and production in the central Gulf of California estimated from satellite data = Variaciones anual y semianual de la biomasa y producción fitoplanctónica en el golfo de California central estimadas de datos de satélite. Cienc. Mar. 39: 217-230. [ Links ]
Andrefouet S, Zubia M, Payri C. 2004. Mapping and biomass estimation of the invasive brown algae Turbinaria ornata (Turner) J. Agardh and Sargassum mangarevense (Grunow) Setchell on heterogeneous Tahitian coral reefs using 4-meter resolution IKONOS satellite data. Coral Reefs 23: 26-38. [ Links ]
Casas-Valdez MM. 2009. El alga marina Sargassum (Sargassaceae) en el desarrollo regional. In: Urciaga-García JI, Beltrán-Morales LF, Lluch-Belda D (eds.), Recursos Marinos y Servicios Ambientales en el Desarrollo Regional. Centro de Investigaciones Biológicas del Noroeste, Universidad Autónoma de Baja California Sur, Centro Interdisciplinario de Ciencias Marinas-IPN, La Paz, BCS, Mexico, pp. 139-156. [ Links ]
Casas-Valdez M, Hernández-Contreras H, Marín-Álvarez A, Águila-Ramírez RN, Hernández-Guerrero CJ, Sánchez Rodríguez I, Carrillo-Domínguez S. 2006a. El alga marina Sargassum (Sargassaceae): Una alternativa tropical para la alimentación de ganado caprino. Rev. Biol. Trop. 54: 83-92. [ Links ]
Casas-Valdez M, Portillo-Clark G, Águila-Ramírez RN, RodríguezAstudillo S, Sánchez-Rodríguez I, Carrillo-Domínguez S. 2006b. Effect of the marine algae Sargassum spp. on the productive parameters and cholesterol content of the brown shrimp, Farfantepenaeus californiensis (Holmes, 1990). Rev. Biol. Mar. Oceanogr. 41: 97-105. [ Links ]
Casas-Valdez M, Sánchez-Rodríguez I, Hernández-Carmona G. 1993. Evaluación de Sargassum spp. en la costa oeste de Bahía Concepción, BCS, México. Inv. Mar. CICIMAR 8: 61-69. [ Links ]
Cohen J, Cohen P, West SG, Aiken LS. 2003. Applied Multiple Regression/correlation Analysis for the Behavioral Sciences. 3rd ed. Lawrence Erlbaum Associates, Mahwah, NJ. [ Links ]
Dawson EY. 1944. Marine algae of the Gulf of California. Allan Hancock Pacific Expeditions 3: 189-454. [ Links ]
Espinoza-Avalos J. 1990. Estructura por edades y reproducción de tres poblaciones de Sargassum sinicola (Phaeophyta, Fucales) en la Bahía de La Paz, Golfo de California. Acta Bot. Mex. 11: 1-9. [ Links ]
Espinoza-Avalos J. 1993. Macroalgas marinas del Golfo de California. In: Salazar-Vallejo SI, González NE (eds.), Biodiversidad Marina y Costera de México. Comisión Nacional de Biodiversidad y CIQRO, Mexico City, pp. 328-357. [ Links ]
ESRI. 2008. ArcView Software. Geographical Information System. Environmental Systems Research Institute, Redlands, CA. [ Links ]
Halfar J, Godínez-Orta L, Mutti M, Valdez-Holguín JE, Borges JM. 2004. Nutrients and temperature controls on modern carbonate production: An example from the Gulf of California, Mexico. Geology 32: 213-216. [ Links ]
Halfar J, Godínez-Orta L, Mutti M, Valdez-Holguín JE, Borges JM. 2006. Carbonates calibrated against oceanographic parameters along a latitudinal transect in the Gulf of California, Mexico. Sedimentology 53: 1-24. [ Links ]
Hernández-Carmona G, Casas Valdez M, Fajardo León C, SánchezRodríguez I, Rodríguez-Montesinos E. 1990. Evaluación de Sargassum spp. en la Bahía de La Paz, BCS, México. Inv. Mar. CICIMAR 5: 11-18. [ Links ]
Huerta-Díaz MA, De León-Chavira F, Lares ML, Chee-Barragan A, Siqueiros-Valencia A. 2007. Iron, manganese and trace metal concentrations in seaweeds from the central west coast of the Gulf of California. Appl. Geochem. 22: 1380-1392. [ Links ]
Marín-Álvarez A, Casas M, Carrillo S, Hernández H, Monroy A. 2003. Performance of sheep feed rations with Sargassum spp. sea algae. Cuban J. Agric. Sci. 37: 119-123. [ Links ]
Muñeton-Gómez MS, Hernández-Carmona G. 1993. Crecimiento estacional de Sargassum horridum (Setchell & Gardner), Phaeophyta, en la Bahía de La Paz, B.C.S. México. Inv. Mar. CICIMAR 8: 23-31. [ Links ]
Muñoz-Ochoa M, Murillo-Álvarez JI, Zermeño-Cervantes LA, Martínez-Díaz SF, Riosmena-Rodríguez R. 2010. Screening of extracts of algae from Baja California Sur, Mexico as reversers of the antibiotic resistance of some pathogenic bacteria. Eur. Rev. Med. Pharmacol. Sci. 14(9): 739-747. [ Links ]
Noiraksar T, Sawayama S, Phauk S, Komatsu T. 2014. Mapping Sargassum beds off the coast of Chon Buri Province, Thailand, using ALOS AVNIR-2 satellite imagery. Bot. Mar. 57: 367-377. [ Links ]
Norris JN. 2010. Marine algae of the northern Gulf of California: Chlorophyta and Phaeophyceae. Smithsonian Contributions to Botany, Smithsonian Institution, Washington, DC. [ Links ]
Núñez-López RA, Casas-Valdez M. 1996. Fenología de las especies de Sargassum (Fucales: Sargassaceae) en tres zonas de la Bahía Concepción, B.C.S., México. Rev. Biol. Trop. 44: 455-464. [ Links ]
Núñez-López RA, Casas-Valdez M. 1997. Variación estacional de la biomasa y talla de Sargassum spp. (Sargassaceae, Phaeophyta) en la Bahía Concepción, B.C.S., México. Hidrobiológica 7: 19-25. [ Links ]
Pacheco-Ruíz I, Zertuche-González JA, Chee-Barragán A, Blanco Betancourt R. 1998. Distribution and quantification of Sargassum beds along the west coast of the Gulf of California, Mexico. Bot. Mar. 41: 203-208. [ Links ]
Patrón-Prado M, Acosta-Vargas B, Serviere-Zaragoza E, Méndez Rodríguez LC. 2010 Copper and cadmium biosorption by dried seaweed Sargassum sinicola in saline waster. Water, Air, Soil Pollut. 210: 197-202. [ Links ]
Rivera M, Scrosati R. 2006 Population dynamics of Sargassum lapazeanum (Fucales, Phaeophyta) from the Gulf of California, Mexico. Phycologia 45: 178-189. [ Links ]
Rodríguez-Montesinos YE, Arvizu-Higuera D, Hernández-Carmona G. 2008 Seasonal variation in length and chemical constituents of Sargassum sinicola Setchell & Gardner from Bahia de La Paz, Baja California Sur, México. Phycol. Res. 56: 33-38. [ Links ]
Sánchez-Rodríguez I, Cervantes-Duarte R. 1999 Longitud y biomasa de Sargassum sinicola Setchell et Gardner (Phaeophyta) en Bahía Magdalena, BCS, México. Hidrobiológica 9: 117-124. [ Links ]
Setchell WA, Gardner NL. 1924 Expedition of the California Academy of Sciences to the Gulf of California in 1921. The marine algae. Proc. Calif. Acad. Sci. Ser. 4, 12: 695-949. [ Links ]
Staehr PA, Pedersen MF, Thomsen MS, Wernberg T, Krause-Jensen D. 2008. Invasion of Sargassum muticum in Limfjorden (Denmark) and its possible impact on the indigenous macroalgal community. Mar. Ecol. Prog. Ser. 207: 79-88. [ Links ]
Tweedley JR, Jackson EL, Attrill MJ. 2008. Zostera marina seagrass beds enhance the attachment of the invasive alga Sargassum muticum in soft sediments. Mar. Ecol. Prog. Ser. 354: 305-309. [ Links ]
Yabur R, Bashan Y, Hernández-Carmona G. 2007 Alginate from the macroalgae Sargassum sinicola as a novel source for microbial immobilization material in wastewater treatment and plant growth promotion. J. Appl. Phycol. 19: 43-53. [ Links ]
Yeong BML, Wong CL, Tan SG, Zainudin NAI, Yusoh OM. 2012 Three months' monitoring of environmental factors, biomass, length and size classes variation of Sargassum species at Cape Rachado, Port Dickson. In: Pertanika Journal of Tropical Agricultural Science, Universiti Putra Malaysia Press, Serdang, Solangor, pp. 623-630. [ Links ]
Recibido: Octubre de 2015; Aprobado: Abril de 2016
 This is an open-access article distributed under the terms of the Creative Commons Attribution License
This is an open-access article distributed under the terms of the Creative Commons Attribution License
