










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCiencias marinas
versión impresa ISSN 0185-3880
Cienc. mar vol.39 no.1 Ensenada mar. 2013
https://doi.org/dx.doi.org/10.7773/cm.v39i1.2175
Artículos
Influencia de las características ambientales de los manglares sobre foraminíferos bénticos recientes en el golfo de Urabá, Caribe colombiano
Influence of the environmental characteristics of mangrove forests on recent benthic foraminifera in the Gulf of Urabá, Colombian Caribbean
Eliana Gómez*, Gladys Bernal
Grupo de investigación OCEANICOS (Oceanografía e ingeniería costera), Escuela de Geociencias y Medio Ambiente, Facultad de Minas, Universidad Nacional de Colombia-Sede Medellín, Carrera 80 No 65-223.
* Corresponding author. E-mail:egomezoc@unal.edu.co
Received July 2012,
received in revised form November 2012,
accepted January 2013.
RESUMEN
En el golfo de Urabá (Colombia) ocurren varios bosques de manglar con diferentes características. En 82 muestras de sedimentos se estudiaron las especies de foraminíferos y su dependencia ambiental. Diez especies de concha algutinada y una de concha calcárea dominaron en estos manglares. No se observaron foraminíferos en zonas sometidas a alta energía hidráulica (oleaje) o alta intervención antropogénica. Se encontraron dos asociaciones características. La primera, representada por Milliammina fusca, la especie más dominante, se asoció con sustratos más finos y menores salinidades. La segunda, representada por Amonia beccarii, la especie calcárea más abundante, se asoció con ambientes contaminados y de bajo oxígeno. Haplophragmoides canariensis y Trochammina squamata, especies pequeñas, tuvieron preferencia por lugares con abundante materia orgánica. Los resultados permitieron plantear un modelo conceptual que relaciona los foraminíferos bénticos de los manglares con la hidrodinámica, el tamaño de grano, el contenido de materia orgánica y la contaminación.
Palabras clave: ecología, foraminíferos bénticos, estuario, manglares.
ABSTRACT
Mangrove forests with different characteristics have developed in the Gulf of Urabá (Colombia). Eighty-two sediment samples were studied for species of foraminifera and their environmental dependence. Ten agglutinated species and one calcareous species dominated in these mangroves. Foraminifera were absent in mangrove areas exposed to high hydraulic energy (waves) or high human affectation. Two characteristic assemblages were found. The first one, represented by Milliammina fusca, the most dominant species, was related to finer substrates and lower salinities. The second one, represented by Ammonia beccarii, the most common calcareous species, was found in polluted low-oxygen environments. Haplophragmoides canariensis and Trochammina squamata, small species, showed a preference for places with abundant organic matter. A conceptual model is proposed that relates the benthic foraminifera to the hydrodynamics, grain size, organic matter content, and pollution.
Key words: ecology, benthic foraminifera, estuary, mangrove forests.
INTRODUCCIÓN
Los foraminíferos bénticos (FB) en ambientes de manglar se han caracterizado por presentar concha aglutinada (Murray 2006) y por ser de tamaños más pequeños que en otros ambientes, incluso menores que 63 μm. Las asociaciones pueden variar en el porcentaje de foraminíferos aglutinados, abundancias y diversidad, desde el manglar hacia fuera de la costa (Culver 1990).
Al parecer los factores ambientales específicos que actúan sobre los FB en manglares son diversos y pueden tener relaciones complejas. Estudios han mostrado que la disponibilidad de oxígeno (Vanicek et al. 2000, Diz y Francés 2008), el tamaño de grano (Debenay et al. 2002, Diz y Francés 2008, Chatelet et al. 2009, Mojtahid et al. 2009), la cantidad y calidad de materia orgánica (Wolfgang 1996, Chatelet et al. 2009, Nooijer et al. 2008), la exposición aérea y elevación vertical (Debenay et al. 2002), la influencia de agua marina y dulce (Debenay y Thi-Luan 2006, Mojtahid et al. 2009), la geomorfología (Alongi y Robertson 1992), la composición vegetal (Morrisey et al. 2003) y las olas y corrientes (Culver 1990) son algunos de los factores que tienen influencia en la distribución y abundancia de los FB en estos ecosistemas.
En el límite oeste del Caribe colombiano, cerca de Panamá, se localiza el golfo de Urabá (GU) (fig. 1). El GU es un cuerpo de agua semicerrado de aproximadamente 2000 km2 y presenta diferentes tipos de manglares a lo largo de la costa, que ocupan unos 70 km2. Este sistema estuarino recibe descargas de varios ríos, en especial del río Atrato, el segundo caudal más grande que desemboca en el mar Caribe (2421 m3 s–1). Este río forma un delta que separa el GU en dos partes, de las cuales la parte sureña es un cuerpo semicerrado denominado bahía Colombia. El río tiene una gran influencia en los patrones de distribución de sedimentos y contaminantes dentro del GU; la circulación general hace que las aguas dulces de la pluma turbia salgan recostadas por el margen oriental (Montoya 2010).
En el GU se encuentran diferentes bosques de manglar (fig. 1). (1) Manglares del delta del río Atrato: 78.3%, bosques muy bien desarrollados (Urrego et al. 2010) en canales distributarios, bahías deltaicas y pantanos entre barras; poca afectación antropogénica, sólo hay pequeñas poblaciones de máximo 350 habitantes en el área (Taborda-Marín et al. 2008), pero alto grado de infección fitosanitaria (32.5%, Urrego et al. 2010). (2) Manglares de la ensenada de Rionegro: 7.2%, bosques en estado sucesional temprano alrededor de una laguna costera delimitada por barras arenosas; presentan elevada influencia antropogénica, 70 hectáreas perdidas en 2004 por diversas causas (Corpourabá 2011). (3) Manglares de las bahías de Turbo y El Uno: 2.9%, las bahías están separadas del cuerpo principal del GU por grandes barreras depositadas por el río Turbo, el segundo en importancia en el GU; bosques altamente intervenidos cerca de la zona urbana del municipio de Turbo, el más grande en las riberas del golfo, (135,967 habitantes, http://www.turbo-antioquia.gov.co). (4) Manglares en las bocas de cauces: 17%, pequeños bosques en las desembocaduras de ríos menores, a menudo muy afectados por la tala.
El GU ofrece una oportunidad para estudiar la ecología de los FB en manglares, por su variedad de entornos, desarrollo y afectación antropogénica. Los FB del GU han sido estudiados por Vargas (2011), quien encontró que la salinidad es el principal factor que controla la distribución de éstos en el gran estuario, pero la fauna de FB en los manglares no ha sido estudiada. El objetivo de este trabajo fue determinar la biodiversidad y distribución de FB en los bosques de manglares del GU y su relación con algunos factores ecológicos como sedimentos, propiedades del agua, morfología y grado de afectación antropogénica.
METODOLOGÍA
Se analizaron un total de 82 muestras tomadas de los sedimentos asociados con los manglares del GU en junio y agosto de 2009 (época húmeda) y febrero de 2010 (época seca) a profundidades de entre 3 cm y 5 m (fig. 1). El número de muestras tomadas fue proporcional al área de cada bosque de manglar y con una separación aproximada de 2km entre las estaciones, distribuidas de la siguiente manera: 53 muestras en los manglares del delta del Atrato, 14 en la ensenada de Rionegro, 6 en las bahías de Turbo y El Uno, y 9 en las bocas de cauces a lo largo del margen del golfo. La mayoría de las muestras fueron tomadas al lado de los mangles (42) y las demás, en las lagunas o canales asociados. Sedimentos superficiales fueron recolectados con una draga Ekman y se tomaron muestras de 100 cm3 del primer centímetro. A las muestras se les adicionaron 20 gotas de alcohol al 95%, y 15 minutos después se les adicionó alcohol al 70% y luego se conservaron en hielo (Murray 2006). Aunque los FB pueden vivir enterrados varios centímetros, se muestreó el primer centímetro superficial buscando que las conchillas contenidas en el sedimento analizado fueran recientes. Además, se ha demostrado que el primer centímetro superficial es el más representativo ecológicamente (Schonfeld et al. 2012), que en ambientes someros no se han visto cambios significativos en las asociaciones con la profundidad (Sen Gupta 1999) y que la profundidad a la cual los FB penetran en los sedimentos es controlada en gran parte por el contenido de oxígeno (Gooday et al. 2000); por tanto, en ambientes de manglar, ricos en materia orgánica, se esperan hábitos preferencialmente epibénticos. Sin embargo, hay que tener en cuenta que las interpretaciones ambientales a partir de los sedimentos más superficiales pueden subestimar algunas especies infaunales.
La salinidad y temperatura fueron medidas con una sonda multiparamétrica YSI. La sonda para medir oxígeno presentó problemas en el campo y no se obtuvieron datos de oxígeno. Otros parámetros de calidad de agua fueron tomados de los informes del Monitoreo de Calidad del Agua del Invemar (REDCAM 2010, http://www.invemar.org.co/psubcategorias.jsp?idsub=252&idcat=105).
Los sedimentos fueron lavados a través de tamices de 355, 150 y 63 μm. Con el fin de diferenciar los organismos vivos durante la colecta, las tres fracciones fueron teñidas con rosa de bengala como se describe en Murray (2006). Cada fracción seca fue dividida dependiendo de su tamaño y se hicieron conteos en alícuotas previamente pesadas hasta obtener al menos 100 individuos, para un total aproximado de 300 individuos por muestra. En algunas muestras no fue posible completar los 300 individuos debido a la escasez de foraminíferos. El número de individuos por muestra fue calculado tomando en cuenta los pesos de las alícuotas medidas y de las fracciones.
Se determinó la biocenosis (individuos teñidos con rosa de bengala), para diferenciar los organismos vivos de los muertos en el momento de la captura. Sin embargo, el total de individuos (vivos + muertos) fue utilizado en el análisis final. La conveniencia de trabajar con el total de individuos se discute en Debenay et al. (2002) y Chatelet et al. (2004), quienes muestran que el total de los individuos es un buen indicador de condiciones ambientales.
Para la identificación de las especies se usó la clasificación de Loeblich y Tappan (Sen Gupta 1999) y las planchas de Barker (1960), Buzas et al. (1977), McCulloch (1977), Buzas y Severin (1982), Boltovskoy e Hincapié de Martínez (1983), Parada y Pinto (1986), Parada (1991), Debenay et al. (2002), Debenay y Thi-Luan (2006) y Bernal et al. (2008).
Paralelo con la recolección de muestras para foraminíferos, se tomaron muestras para análisis sedimentológicos. Se secaron a 60 ºC. Se midió la materia orgánica (MO) por calcinación y los carbonatos se midieron con un calcímetro de Bernard. La granulometría de arenas y lodos se hizo por separado, a través de tamizado en seco y el método de la pipeta. En muestras pequeñas (con alto contenido de agua respecto al sedimento) sólo se estimaron los porcentajes de lodos (%L) y el tamaño de granos. Los parámetros granulométricos de la media, la asimetría y la selección fueron obtenidos por el método de los momentos (Tucker 1988).
Para analizar los datos se utilizaron métodos estadísticos como matrices de correlación y análisis de conglomerados. Se calculó el índice de diversidad de Shannon como se describe en Mojtahid et al. (2009). Finalmente, se realizó un modelo conceptual de los principales factores ecológicos que están afectando la distribución y abundancia de los foraminíferos bénticos en los manglares del GU.
RESULTADOS
Sólo en 35 localidades se encontraron FB (fig. 2), con abundancias máximas de 66 ind cm–3. Las mayores abundancias ocurrieron en los manglares del delta del río Atrato y de las bahías Turbo y El Uno (hasta 66 y 26 ind cm–3, respectivamente); sin embargo, sólo en dos muestras de 6 de esta última localidad hubo foraminíferos. En la ensenada de Rionegro sólo se encontraron FB en dos muestras, con valores máximos de 3 ind cm–3. En los pequeños bosques de la ribera del GU, en las bocas de ríos menores, no se encontraron FB.
Un total de 18 géneros y 26 especies fueron identificados (fig. 3); 12 de las especies pertenecen al orden Rotaliida (concha calcárea) y 14 al orden Textulariida (concha aglutinada). Las especies aglutinadas presentaron las abundancias más altas, y Milliammina fusca fue la más abundante. Por su parte, Ammonia beccarii fue la especie calcárea con mayor ocurrencia. Los FB aglutinados estuvieron presentes en todas las muestras donde se presentaron foraminíferos, con valores entre 55% y 100% (a excepción de la ensenada de Rionegro, donde sólo se presentó la especie calcárea A.beccari). El índice de Shannon Weaver presentó valores entre 0 y 0.66.
Once especies de FB, diez de ellos aglutinados, tuvieron las mayores abundancias (> 5 ind cm–3): M. fusca (61%), Haplophragmoides canariensis (7%), Ammobaculites exiguus (7%), Trochammina squammata (5%), Ammobaculites exiliis (5%), Ammotium salsum (3%), Ammobaculites americanus (3%), Arenoparrella mexicana (2%), Ammonia beccarii (2%), Trochammina inflata (1%) y Haplophragmoides wilbertii (1%). Las especies que se tiñeron con rosa de bengala fueron M. fusca y A. exilis. Haplophragmoides canariensis y T. squammata mostraron preferencia por la fracción más pequeña (63–150 μm); 87% de los individuos de H. canariensis y 67% de T. squamata midieron menos de 150 μm. Por su parte, A. exiguus, A. exilis, A. salsum, T. inflata y A. mexicana prefirieron la fracción de 150–355 μm.
El análisis de conglomerados con las abundancias de las especies más comunes en las 35 localidades donde se presentaron FB (fig. 4) indicó dos grupos principales (G1 y G2). Las localidades del G1 tuvieron las abundancias más altas de A. salsum, A. mexicana y A. beccari, mientras que las localidades del G2 tuvieron las abundancias más altas de M. fusca, H. canariensis, A. exiguus, T. squammata, A. exilis, A. americanus, T. inflata y H. wilbertii. El G1 incluyó muestras con abundancias de M. fusca menores que 8 ind cm–3, mientras que el G2 incluyó muestras con mayores abundancias (entre 8 y 42 ind cm–1). La salinidad en las estaciones del G1 llegó hasta 20 y en las del G2, hasta 12; los sedimentos del G1 tuvieron hasta 41% de carbonatos, mientras que los del G2 sólo hasta 6%. El G1 tuvo los tamaños más grandes de sedimento y las menores abundancias de FB. No hubo diferenciación del porcentaje de especies aglutinadas entre los dos grupos.
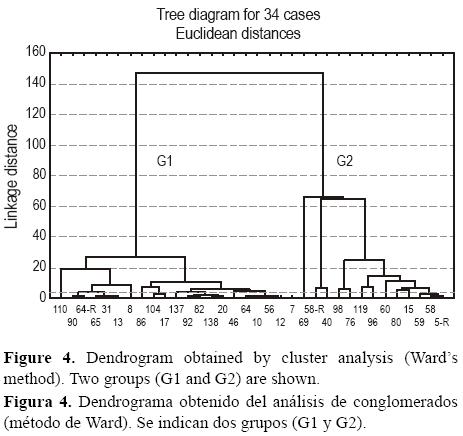
Los dos grupos se dividieron en subgrupos (g1 a g6, tabla 1). Los dos subgrupos del G1 se diferenciaron por sus contenidos de MO (mayor en g2), carbonatos (mayor en g2) y %L (mayor en g1). Los cuatro subgrupos de G2 mostraron abundancias y tamaños de grano decrecientes.
Las correlaciones entre las variables que caracterizan los foraminíferos y las variables ambientales fueron bajas. La salinidad, temperatura y profundidad no tuvieron relaciones significativas con los FB, debido a su homogeneidad entre los sitios muestreados. Correlaciones significativas se presentaron entre H. canariensis y T. squamata con MO (0.26 y 0.31) y T. inflata con % L (0.22).
De acuerdo con REDCAM (fig. 5), se registraron niveles de oxígeno disuelto menores que 4 mg L–1 (valor mínimo según la norma colombiana) en las bocas de los ríos León, Guadualito y Caimán Nuevo, localidades donde no se encontraron FB, así como en la ensenada de Rionegro, donde sólo hubo FB en dos de 12 muestras, al centro de la laguna. Valores de nutrientes (fosfatos, nitratos y amonio) por encima de la norma se encontraron en la boca del río Guadualito, la playa La Martina y la ensenada de Rionegro (sin presencia de FB). Los coliformes sólo se midieron en playas y sobrepasaron la norma en La Martina y Turbo, donde igualmente hubo déficit de FB.
Las relaciones encontradas se sintetizan en el modelo conceptual que se presenta en la figura 6. La hidrodinámica del manglar y su morfología influyen sobre el tamaño de grano; las localidades expuestas al oleaje presentan tamaños gruesos, menor %MO y ausencia de FB. Las localidades resguardadas presentan sustratos lodosos y mayor %MO, que favorecen a M. fusca. Cuando los valores de MO aumentan, disminuyen los porcentajes de carbonato. El desarrollo de foraminíferos aglutinados se ve favorecido con el pH ácido. En manglares con influencia antropogénica hay ausencia o bajas abundancias de FB, dominancia de A. beccarii y bajos valores de oxígeno disuelto en el agua (anoxia).
DISCUSIÓN
En comparación con estudios que consideran hasta 1 cm superficial y todos los tamaños de grano, las abundancias de FB en GU para las fracciones >63 μm (0–66 ind cm–3) tienen los mismos órdenes de magnitud que en otros ambientes de manglar (e.g., Guyana Francesa, 0–40 ind cm–3, Debenay et al. 2002; delta del Mekong, Vietnam, 0.2–300 ind cm–3, Debenay y Thi-Luan 2006; Ría de Vigo, España, 20–80 ind cm–3, Diz y Francés 2008). Las abundancias en los manglares son más bajas que en el cuerpo de agua principal del GU (hasta 92 ind cm–3, Vargas 2011). La riqueza de esepcies (26 especies) es congruente con resultados obtenidos por otros autores en ambientes estuarinos (Debenay et al. 2002, Debenay y Thi-Luan 2006, Bernal et al. 2008, Mojtahid et al. 2009, Semensatto et al. 2009), pero inferior a los registrados para el cuerpo de agua principal del golfo (60 especies, Vargas 2011). Así mismo, la biodiversidad fue baja (índice de Shannon Weaver: 0–0.66). Las bajas abundancias y diversidad en los estuarios y manglares se deben a condiciones no óptimas de salinidad y pH para los FB (Mojtahid et al. 2009).
La composición de la fauna de FB estuvo dominada por una sola especie, M. fusca, con 61% del total. Esta especie también fue dominante, en el estuario de Tina Menor, Cantabria (Pascual et al. 2009). En el GU, M. fusca se encontró principalmente en los manglares del Atrato donde dominaron los sustratos limosos y % L altos (hasta 90%). Esta especie se ha presentado en ambientes con muy baja salinidad (Debenay et al. 2002, Murray 2006,), con %L menores que 66% (Murray 2006), en sedimentos con abundantes arcillas (Chatelet et al. 2009) y en ambientes estresados (Semensatto et al. 2009); no se favorece con depósitos de restos vegetales sobre los sedimentos (Chatelet et al. 2009).
La asociación de especies encontrada es característica de estos ambientes. Horton et al. (2003) observaron la dominancia de M. fusca, A. mexicana y H. canariensis en un ecosistema de manglar australiano. Ammobaculites exiguus se ha registrado en ambientes estuarinos someros donde la salinidad no excede 20 (Scott 2007), está asociada con sedimentos lodosos y puede ser tolerante a la exposición aérea (Semensatto et al. 2009). En los manglares del golfo, A. exiguus mostró preferencia por los sustratos areno lodosos de algunas localidades de bocas del Atrato. Por su parte, T. squamata, se ha caracterizado por encontrarse en sustratos finos (Murray 2006) y ricos en MO (Chatelet et al. 2009). En los manglares del GU estuvo relacionada con el %L y, junto con H. canariensis, con la MO.
La especie calcárea más importante fue A. beccari. En una de las muestras de la ensenada de Rionegro representó el 100% relativo a otras especies. Es una especie común, pero tiene la capacidad de colonizar ambientes estuarinos y de manglar (Debenay et al. 2002, Diz y Francés 2008, Mojtahid et al. 2009). Se asocia con altos contenidos de MO y bajas concentraciones de oxígeno en los sedimentos (Scott 2007, Diz y Francés 2008). Su dominancia en la ensenada de Rionegro podría deberse a los niveles bajos de oxígeno registrados por la REDCAM.
Para el análisis de las relaciones entre los FB y los factores ambientales se tuvieron en cuenta las abundancias totales en las muestras y las condiciones ambientales en el momento de la recolección. Una debilidad del análisis reside en que las muestras fueron recolectadas en tres meses del año, en dos estaciones diferentes. No se realizó un análisis de variabilidad estacional, el cual sería sin duda valioso, pero requeriría un diseño de muestreo específico. El hecho de usar el total de FB (vivos + muertos) encontrados en el primer centímetro superficial implica que la relación con el medio ambiente sea de una asociación de más largo plazo que la estacional.
En investigaciones anteriores se ha observado que las fracciones pequeñas (63–150 μm) pueden contener un gran número de individuos y especies de foraminíferos que raramente se observan en las fracciones grandes, especialmente en ambientes ricos en MO, donde las especies dominantes pueden encontrarse más abundantes en fracciones pequeñas (Shepherd et al. 2007). Los resultados del presente estudio confirman la importancia de analizar las fracciones pequeñas en ambientes de manglar.
La energía hidráulica es un factor abiótico influyentesobre los FB (Schonfeld 2002). Ésta ejerce un efecto importante sobre otras variables como el tamaño de grano y la MO. El tamaño de grano tiene gran influencia sobre los FB (Murray 2006), pero hay algunas ocasiones donde es difícil discriminar su influencia de la de otras variables ambientales (Debenay et al. 2002). Los sedimentos gruesos son típicos de ambientes con oleaje o corrientes y los finos de, ambientes tranquilos. La ausencia de foraminíferos en los lugares del GU expuestos al oleaje (salientes del delta del Atrato) se explica porque éstos prefieren zonas de baja energía donde se facilita el depósito de MO (alimento); además, en ambientes con fuerte oleaje las condiciones varían tan rápidamente que no permiten el establecimiento de los FB (Alve 1999). Asímismo, las fuerzas de las corrientes tienen impactos negativos en riqueza y abundancias (Schonfeld 2002). En general, en los manglares del GU se observó que los lugares expuestos al oleaje y corrientes presentaron mayor tamaño de grano y menores abundancias de foraminíferos, mientras que los ambientes resguardados y depositacionales presentaron altos %L y mayores abundancias de foraminíferos.
Los efectos de la MO sobre los FB son complejos, ya que algunos estudios muestran aumento o decrecimiento en abundancias de foraminíferos con el aumento o disminución de la MO en los sedimentos, mientras que otros revelan no tener influencia (Chatelet et al. 2009). En este estudio, no se observaron foraminíferos en los sedimentos con valores de MO <5% y >40%, y las abundancias fueron mayores en sedimentos con MO entre 15 y 30%. Estos resultados apoyan la hipótesis de Alve (1999), de que la MO favorece el desarrollo de los FB hasta que llega a ser tóxica. En cuanto a la riqueza, en los manglares del GU disminuye cuando se incrementa la MO.
En muchas de las localidades del GU donde no se encontraron foraminíferos se han registrado valores bajos de oxígeno disuelto. Niveles bajos de oxígeno y de carbonatos podrían deberse a concentraciones de MO altas (30%–90%) que producirían anoxia y disminución del pH debido al con-sumo de oxígeno y producción de CO2 en su degradación.
Las aguas costeras del lado oriental del golfo se encuentran influenciadas por contaminación por fertilizantes, hidrocarburos y pesticidas relacionada con la agricultura del banano, principal actividad económica de la región. En bahía Turbo se presenta una alta degradación ambiental debida a basuras, aguas residuales y tráfico marítimo, que coincidió con baja abundancia y diversidad de FB (sólo en una de tres muestras se hallaron foraminíferos, con nueve especies). En el río León, la principal ruta de salida de los barcos bananeros hacia el mar, se ha notado un incremento en los hidrocarburos y pesticidas, y allí no se encontraron foraminíferos. Los anteriores resultados muestran que estas localidades probable-mente están siendo afectadas por el aumento en los valores de hidrocarburos. En otras investigaciones se ha registrado que los hidrocarburos aromáticos pueden ser letales para organismos marinos. Las dosis más bajas pueden inhibir actividades fisiológicas y causar reacciones patológicas (Sen Gupta 1999), además esto también puede ser uno de los causantes de los bajos valores de oxígeno disuelto en el agua.
La baja ocurrencia de foraminíferos en la ensenada de Rionegro podría tener que ver con la intervención antrópica y el tipo de sedimento (altos contenidos de MO y restos vegetales). Este bosque sufrió una degradación muy fuerte en 2004 y los reportes de REDCAM igualmente indican bajos contenidos de oxígeno en el agua. Estos sedimentos tuvieron los mayores porcentajes de carbonatos de los manglares estudiados (hasta 57% respecto a 6.5% en los otros bosques), con restos de conchas de almejas u ostras. Precisamente aquí, cuando se presentaron FB, éstos fueron calcáreos. Esto era de esperarse, pues al tener las aguas mayor disponibilidad de carbonato de calcio, los individuos pueden construir su concha calcárea más fácilmente.
CONCLUSIONES
Este estudio permitió determinar los principales factores ecológicos que actúan sobre los FB en los manglares del GU. La energía hidráulica es un factor importante, relacionada con el tamaño de grano y el %MO. La especie más abundante, M. fusca, se encontró relacionada con el %L; T. squammata y H. canariensis tuvieron preferencia por lugares con abundante MO. En los lugares con alta intervención antropogénica se dieron bajas abundancias de FB con altas concentraciones de nutrientes, bajos niveles de oxígeno y contaminación con hidrocarburos y pesticidas. Allí se presentó la especie A. beccarii, típica colonizadora de ambientes contaminados. Esta especie podría ser usada como bioindicadora de degradación ambiental.
Las asociaciones de FB encontradas son similares a las de la mayoría de ambientes estuarinos alrededor del mundo. Ocurrieron dos asociaciones, una conformada por A. salsum, A. mexicana y A. beccari, y otra por M. fusca, H. canariensis, A. exiguus, T. squammata, A. exilis, A. americanus, T. inflata y H. wilbertii. Especies como H. canariensis y T. squammata fueron más abundantes en la fracción de 63–150 μm, resaltando la importancia de tomar en cuenta las fracciones pequeñas en el análisis de FB en manglares.
AGRADECIMIENTOS
Agradecemos a la Gobernación de Antioquia (Colombia) por el apoyo económico que hizo posible esta investigación. Gracias a Mauricio Ruiz-Ochoa por su apoyo y asesoría, y a Ligia E Urrego por sus valiosos aportes. Jose D Ramírez colaboró con los análisis sedimentológicos de las muestras, y Mario A Jiménez y Jaime A Carmona ayudaron con la programación. Agradecemos también a los compañeros de Expedición Estuarina, golfo de Urabá, Fase I, por su acompañamiento, a los ayudantes de campo por su colaboración, así como a los habitantes de la región de Urabá por permitir los muestreos.
REFERENCES
Alongi DM, Robertson AI. 1992. Tropical Mangrove Ecosystems. American Geophysical Union, Washington, DC, 329 pp. [ Links ]
Alve E.1999. Colonization of new habitats by benthic foraminifera: A review. Earth-Sci. Rev. 46: 167–185. [ Links ]
Barker R. 1960. Taxonomic notes on the especies figured by H.B. Brady in his report on the foraminifera dredged by H.M.S Challenger during the years 1873–1876. Vol. 9. Society of Economy Paleontology and Mineralogy. [ Links ]
Bernal G, Ruiz-Ochoa MA, Piedrahita MT, Restrepo E. 2008. Foraminíferos en los sedimentos superficiales del sistema lagunar de Cispatá y la interacción Río Sinú-Mar Caribe Colombiano. Bol. Ciencias de la Tierra 23: 5–20. [ Links ]
Boltovskoy E, Hincapié de Martínez S. 1983. Foraminíferos del manglar de Tesca, Cartagena, Colombia. Rev. Esp. Micropaleontol. 15: 205–220. [ Links ]
Buzas M, Severin K. 1982. Distribution and systematics of foraminifera in the Indian River, Florida. Smithson. Contrib. Paleobiol. 16: 1–73. [ Links ]
Buzas M, Smith R, Beem, K. 1977. Ecology and systematics offoraminifera in two thalassia habitats, Jamaica, West Indies. Smithson. Contrib. Paleobiol. 31: 139 pp. [ Links ]
Chatelet E, Debenay JP, Soulard R. 2004. Foraminiferal proxies forpollution monitoring in moderately polluted harbors. Environ. Pollut. 127: 27–40. http://dx.doi.org/10.1016/S0269-7491(03)00256-2. [ Links ]
Chatelet EA, Bout-Roumazeilles V, Riboulleau A, Trentesaux A. 2009. Sediment (grain size and clay mineralogy) and organicmatter quality control on living benthic foraminifera. Rev.Micropaleontol. 52: 75–84. http://dx.doi.org/10.1016/j.revmic.2008.10.002 [ Links ]
Corpourabá. 2011. En la Ensenada de Rionegro-Necoclí Avanza Recuperación de las Ostras del Manglar. Corporación para el Desarrollo Sostenible del Urabá. http://www.corpouraba.gov.co/en-la-ensenada-de-rionegro-necocli-avanza-recuperacion-de-las-ostras-del-manglar. Accessed on 26 February 2013. [ Links ]
Culver SJ. 1990. Benthic foraminifera of Puerto Rican mangrove-lagoon systems: Potential for paleoenvironmental interpretations. Deep Sea Res. (B Oceanogr. Lit. Rev.) 37: 34–51. http://dx.doi.org/10.1016/S0198-0254(06)80381-X [ Links ]
Debenay JP, Thi-Luan B. 2006. Foraminiferal assemblages and the confinement index as tools for assessment of saline intrusion and human impact in the Mekong Delta and neighbouring areas (Vietnam). Rev. Micropaléontol. 49: 74–85. http://dx.doi.org/10.1016/j.revmic.2006.01.002 [ Links ]
Debenay JP, Guiral M, Parra D. 2002. Ecological factors acting onthe microfauna in mangrove swamps. The case of foraminiferalassemblages in French Guiana. Estuar. Coast. Shelf Sci. 55: 509–533. http://dx.doi.org/10.1006/ecss.2001.0906 [ Links ]
Diz P, Francés G. 2008. Distribution of live benthic foraminifera in the Ría de Vigo (NW Spain). Mar. Micropaleontol. 66: 165–191. http://dx.doi.org/10.1016/j.marmicro.2007.09.001 [ Links ]
Gooday AJ, Bernhard JM, Levin LA, Suhr SB. 2000. Foraminiferain the Arabian Sea oxygen minimum zone and other oxygen-deficient settings: Taxonomic composition, diversity, and relation to metazoan faunas. Deep-Sea Res. II 47: 25–54. http://dx.doi.org/10.1016/S0967-0645(99)00099-5 [ Links ]
Horton BP, Larcombe P, Woodroffe SA, Whittaker SA, Wright MR, Wynn C. 2003. Contemporary foraminiferal distributions of amangrove environment, Great Barrier Reef coastline, Australia:Implications for sea-level reconstructions. Mar. Geol. 198: 225–243. http://dx.doi.org/10.1016/S0025-3227(03)00117-8 [ Links ]
McCulloch I. 1977. Qualitative observations on recent foraminifera test with emphasis on the Eastern Pacific. University of Southern California, Los Angeles. [ Links ]
Mojtahid M, Jorissen F, Lansard B, Fontanier C, Bombled B, Rabouille C. 2009. Spatial distribution of live benthic foraminifera in the Rhone prodelta: Faunal response to a continental-marine organic matter gradient. Mar. Micropaleontol. 70: 177–200. http://dx.doi.org/10.1016/S0025-3227(03)00117-8 [ Links ]
Montoya LJ. 2010. Dinámica oceanográfica del Golfo de Urabá y su relación con los patrones de dispersión de contaminantes y sedimentos. PhD thesis, Facultad de Minas, Universidad Nacional de Colombia. [ Links ]
Morrisey DJ, Skilleter GA, Ellis Burns JI, Kemp CE, Burt CE.2003. Differences in benthic fauna and sediment among mangrove (Avicennia marina var. australasica) stands ofdiferent ages in New Zeland. Estuar. Coast. Shelf Sci. 56: 581–592. http://dx.doi.org/10.1016/S0272-7714(02)00208-1 [ Links ]
Murray J. 2006. Ecology and applications of benthic foraminifera. Cambridge University Press, 426 pp. [ Links ]
Nooijer LJ, Duijnstee IAP, Bergman MJN, Van der Zwaan MJN. 2008. The ecology of benthic foraminifera across the Frisian Front, southern North Sea. Estuar. Coast. Shelf Sci. 78: 715–726. http://dx.doi.org/10.1016/j.ecss.2008.02.015 [ Links ]
Parada C. 1991. Claves de identificación para géneros de foraminíferos. Curso Universidad Nacional de Colombia. Bogotá D.C. [ Links ]
Parada C, Pinto J. 1986. Foraminíferos bénticos recientes de isla Barú. Fondo FEN Colombia. Fondo FEN Colombia, Bogotá D.C., 212 pp. [ Links ]
Pascual A, Martínez García B, Rodríguez Lázaro J, Martín-Rubio M. 2009. Distribución de los foraminíferos bentónicos recientes en el estuario de Tina Menor (Cantabria). Geogaceta 47: 45–48. [ Links ]
Schonfeld J. 2002. A new benthic foraminiferal proxy for near-bottom current velocities in the Gulf of Cadiz, northeastern Atlantic Ocean. Deep-Sea Res. (I Oceanogr. Res. Pap.) 49: 1853–1875. http://dx.doi.org/10.1016/S0967-0637(02)00088-2 [ Links ]
Schonfeld J, Alve E, Geslin E, Jorissen F, Korsun S, Spezzaferri S. 2012. The FOBIMO (FOraminiferal BIo-MOnitoring) initiative:Towards a standardised protocol for soft-bottom benthic foraminiferal monitoring studies. Mar. Micropaleontol. 94:1–13. http://dx.doi.org/10.1016/j.marmicro.2012.06.001 [ Links ]
Scott DB. 2007. Monitoring in Coastal Environments using Foraminifera and Thecamoebian Indicators. Cambridge University Press. [ Links ]
Semensatto Jr. DL, Hideki Ferreira Funo R, Dias-Brito D, Coelho Jr. C. 2009. Foraminiferal ecological zonation along a Brazilian mangrove transect: Diversity, morphotypes and the influence of subaerial exposure time. Rev. Micropaleontol. 52: 67–74. http://dx.doi.org/10.1016/j.revmic.2008.06.004 [ Links ]
Sen Gupta B. 1999. Modern Foraminifera. Kluwer Academic Publishers, 361 pp. [ Links ]
Shepherd AS, Rathburn AE, Pérez ME. 2007. Living foraminiferalassemblages from the Southern California margin: A comparison of the >150, 63–150, and >63 μm fractions. Mar. Micropaleontol. 65: 54–77. http://dx.doi.org/10.1016/j.marmicro.2007.06.001 [ Links ]
Taborda-Marín A, Maldonado-Lizarazo C, Palacio-Baena JM, Vásquez-Arango JG. 2008. Participación comunitaria en el Golfo de Urabá: El caso del corregimiento Bocas del Atrato y la vereda El Roto, Municipio de Turbo, Antioquia. Rev. Gestión Ambiente 11: 97–108. [ Links ]
Tucker M. 1988. Techniques in Sedimentology. Blackwell Scientific Publications, Oxford. [ Links ]
Urrego LE, Molina EC, Suárez JA, Ruiz HY, Polanía J. 2010. Distribución, composición y estructura de los manglares del Golfo Urabá. En: Informe técnico del proyecto Expedición estuarina Golfo de Urabá, 2013. Gobernación de Antioquia, Medellín (Colombia). [ Links ]
Vanicek V, Juracic M, Bajraktarevic Z, Cosovic V. 2000. Benthicforaminiferal assemblages in a restricted environment. An example from Mjet lakes (Adriatic Sea, Croatia). Geol. Croatica 53: 269–279. http://dx.doi.org/10.1016/j.marmicro.2007.06.001 [ Links ]
Vargas ME. 2011. Foraminíferos del Golfo de Urabá y su potencial como paleoindicadores y bioindicadores de cambios ecológicos.Msc Thesis, Facultad de Minas, Universidad Nacional de Colombia. [ Links ]
Wolfgang K. 1996. Ecological structuring and evolution of deep seaagglutinated foraminifera: A review. Rev. Micropaleontol. 39:271–281. http://dx.doi.org/10.1016/S0035-1598(96)90119-1 [ Links ]

