










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkCiencias marinas
Print version ISSN 0185-3880
Cienc. mar vol.37 n.1 Ensenada Mar. 2011
Variaciones estacionales del flujo vertical de materia orgánica particulada en la región central del Golfo de California
Seasonal variability of the particulate organic matter vertical flux in the central region of the Gulf of California*
J García–Pámanes1*, A Trasviña–Castro2, JR Lara–Lara3, C Bazán–Guzmán3
1 Departamento de Biología Marina, Universidad Autónoma de Baja California Sur, Km 5.5 carretera al sur, Apartado postal 19–B, La Paz 23081, Baja California Sur, México.
2 Centro de Investigación Científica y de Educación Superior de Ensenada, Unidad La Paz, Departamento de Oceanografía Física, Calle Miraflores No. 334, Col. Bella Vista, La Paz 23050, Baja California Sur, México.
3 Centro de Investigación Científica y de Educación Superior de Ensenada, División de Oceanología, Carretera Tijuana–Ensenada No. 3918, Zona Playitas, Ensenada 22860, Baja California, México.
*Corresponding author.
E–mail: jgpamanes@uabcs.mx
Received January 2010
Accepted October 2010
RESUMEN
Se efectuaron cuatro cruceros en la Cuenca de Guaymas (otoño de 1994, y primavera, verano y otoño de 1995) para estudiar las variaciones estacionales de los flujos de materia orgánica particulada (MOP) y la cantidad exportada desde los primeros 200 m de la columna de agua. Se usó un arreglo de trampas de sedimentación de deriva colocadas a 50, 100 y 200 m en la capa superior de la columna de agua para medir los flujos de partículas. Las profundidades promedio de la capa de mezcla variaron entre 20.8 m en verano de 1995 y 29.6 m en primavera de 1995, y las de la zona eufótica entre 33.2 m en primavera de 1995 y 51.6 m en verano de 1995. El promedio de la biomasa del fitoplancton presentó un intervalo entre 36.9 mg Cla m–2 en otoño de 1995 y 85.9 mg Cla m–2 en primavera de 1995. Los flujos máximos de MOP ocurrieron en otoño de 1994 y primavera de 1995, y los mínimos en verano y otoño de 1995. El porcentaje de la producción primaria diaria exportada desde los primeros 200 m varió entre 3.6% y 8.9% y se encuentra dentro del intervalo documentado para otras regiones. Los valores de C:N se mantuvieron homogéneos, independientemente de la profundidad, variando entre 9.0 y 13.8. El tiempo de residencia del carbono orgánico particulado varió entre 3.7 y 11.2 d, siendo mayor en verano que en otoño. Consecuentemente, los organismos heterótrofos que viven en el piso oceánico reciben alimento en pulsos estacionales. El carbono total exportado para el área se estimó en cerca de 300 × 103 t año–1, lo cual sugiere que durante el verano esta región se comporta como un sumidero de carbono; sin embargo, es necesario contar con mayor información para conocer el balance anual.
Palabras clave: flujos de materia orgánica particulada, tiempos de residencia, Cuenca de Guaymas, Golfo de California.
ABSTRACT
Seasonal variability of particulate organic matter (POM) fluxes and the amount exported from the upper 200 m of the water column was studied during four oceanographic cruises in Guaymas Basin during autumn 1994 and spring, summer, and autumn 1995. We used an array of drifting sediment traps to measure the particulate fluxes at 50, 100, and 200 m depths in the upper water column. The mean depths of the mixed layer varied from 20.8 m in summer 1995 to 29.6 m in spring 1995, and of the euphotic zone from 33.2 m in spring 1995 to 51.6 m in summer 1995. Mean phytoplankton biomass ranged from 36.9 mg Chla m–2 in autumn 1995 to 85.9 mg Chla m–2 in spring 1995. Higher POM fluxes occurred during autumn 1994 and spring 1995, while lower fluxes occurred during summer and autumn 1995. The percentage of daily primary productivity exported from the upper 200 m of the water column varied from 3.6% to 8.9% and is within the range reported for other regions. The C:N ratios were homogeneous, regardless of depth, and varied from 9.0 to 13.8. The particulate organic carbon residence time varied from 3.7 to 11.2 d, and was higher in summer and lower in autumn. Consequently, benthonic heterotrophic organisms get their food from seasonal pulses. The total exported carbon for the area was estimated to be about 300 × 103 t yr–1, which suggests that during summer, this region acts as a carbon sink; however, more data are needed to understand the yearly balance.
Key words: particulate organic matter fluxes, residence time, Guaymas Basin, Gulf of California.
INTRODUCCIÓN
Conocer el destino de la materia orgánica particulada (MOP) generada en las aguas superficiales de los océanos es relevante por dos razones. Por un lado, la calidad y cantidad de la materia orgánica que se sedimenta fuera de las capas superficiales es de vital importancia para toda la vida heterotrófica bajo la zona eufótica (Smetacek et al. 1978). Por otro lado, el flujo descendente de las partículas biogénicas de la zona eufótica es un proceso central en el ciclo del carbono oceánico (Miquel et al. 1994) y en el secuestro de CO2. Debido a esto, se ha despertado un considerable interés por conocer el papel que juegan los océanos como sumideros para el CO2 atmosférico (Sarmiento y Siegenthaler 1992).
El amplio intervalo de variaciones regionales y estacionales en la cantidad y composición de la materia sedimentada capturada con las trampas de sedimento es una función de la complejidad del sistema pelágico superior (Peinert et al. 1989, Boyd y Newton 1995). La estructura y organización de la comunidad planctónica son unos de los factores que controlan la producción exportada. Los factores biológicos que afectan la exportación pelágica lo hacen a través de un número diverso de mecanismos, incluyendo la sedimentación de células fitoplanctónicas y la formación y hundimiento de cápsulas fecales y otros desechos orgánicos. Sin embargo, el reciclado en la zona eufótica también es un proceso importante (Miquel et al. 1994).
Algunas estimaciones indican que el contenido de carbono orgánico del material particulado que se hunde excede de 1.2 a 5.1 veces el contenido en el zooplancton que migra hacia arriba (Harding et al. 1987). Longhurst y Williams (1992) también encontraron que la cantidad de carbono orgánico transportado por copépodos migrantes es pequeña comparada con el flujo de carbono exportado vía partículas en los primeros 200 m.
El Golfo de California ha sido definido por diversos autores como un ecosistema altamente productivo (Zeitzschel 1969, Valdez–Holguín y Lara–Lara 1987), pero se desconocen muchos de los procesos físicos y biológicos que intervienen en la transferencia de materia de un nivel trófico a otro y de material particulado hacia el fondo marino.
Lara–Lara et al. (2007) reportaron datos preliminares sobre los flujos de MOP para esta región. En este trabajo se presentan con mayor profundidad las interrelaciones de dichos flujos con las características físicas de la región, como son la variabilidad de la capa de mezcla, la capa isotermal y el transporte horizontal de las parcelas de agua. Asimismo, se estimó el monto total de carbono exportado para el área de estudio.
El objetivo del presente trabajo fue conocer la variabilidad estacional de los flujos de la MOP total y las fracciones de carbono y nitrógeno, así como estimar la cantidad de materia exportada de los primeros 200 m de la columna de agua hacia el fondo de la región central del Golfo de California.
ÁREA DE ESTUDIO
El Golfo de California se encuentra ubicado entre la Península de Baja California y los estados de Sonora y Sinaloa (fig. 1), en el noroeste de México. Es una cuenca de evaporación donde la entrada de agua dulce es menor que la evaporación, sobre todo al norte de las grandes islas (Maluf 1983). La cuenca mide cerca de 1000 km de largo y 150 km de ancho en promedio.
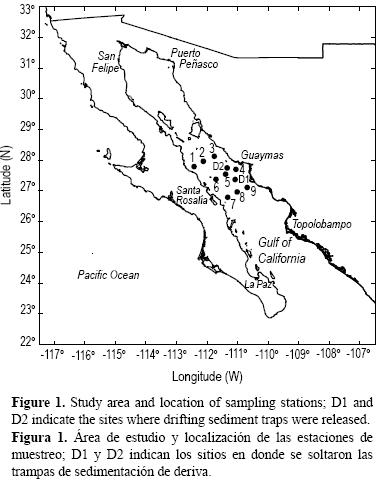
La Cuenca de Guaymas se localiza inmediatamente al sur de la zona de las grandes islas y pertenece a la cuenca sur. La plataforma continental es muy estrecha (menos de 10 km) frente a Baja California y alcanza unos 30 km, en promedio, en la costa continental de México.
Oceanográficamente, la capa superior y la profunda del océano en esta región se caracterizan por la presencia de tres masas de agua: el Agua del Golfo de California, el Agua Subsuperficial Subtropical y el Agua Intermedia del Pacífico. La estructura vertical de estas masas de agua es muy variable durante todo el año (Torres–Orozco 1993). Álvarez–Borrego y Schwartzlose (1979) señalan que hacia fines de primavera, verano y otoño el Agua Superficial Ecuatorial (ASE) invade hasta la porción sur de las islas Ángel de la Guarda y Tiburón, pero Torres–Orozco (1993) encontró que el ASE sólo se presenta en cantidades significativas durante años El Niño. La temperatura superficial en la cuenca alcanza valores mínimos en febrero de entre 14 y 18 °C, aumentando progresivamente hacia el verano, cuando se alcanzan valores entre 29 y 31 °C (Roden y Groves 1959). En octubre se desarrolla una transición entre los patrones antes descritos y ocurren temperaturas más altas en la costa de Baja California, y las surgencias en la costa opuesta bajan la temperatura superficial (Badan–Dangon et al. 1985, Álvarez–Borrego y Lara–Lara 1991).
MATERIALES Y MÉTODOS
Se efectuaron cuatro cruceros en la región central del Golfo de California durante el otoño (4 a 15 de noviembre) de 1994 y la primavera (22 de marzo a 7 de abril), el verano (22 de agosto a 5 de septiembre) y el otoño (21 de noviembre a 7 de diciembre) de 1995. Se cubrió una red de nueve estaciones distribuidas en tres transectos (fig. 1). La distancia entre estaciones fue de 40 km y entre transectos de 64 km. Las estaciones donde se soltaron las trampas de sedimentación se localizaron entre las estaciones de la costa de Sonora y el centro del golfo. En el muestreo de otoño de 1994 la estación se localizó entre el transecto central y el sureño (estación D1), mientras que en los muestreos de 1995 se localizó entre el transecto norteño y el central (estación D2). En todos los casos el barco derivó junto con las trampas de sedimentación, efectuando muestreos cada 6 h.
Se realizaron perfiles de temperatura y salinidad con un CTD SeaBird, y de fluorescencia natural con un perfilador de fluorescencia natural PNF–300 (Biospherical Instruments). Adicionalmente, se tomaron muestras discretas de agua a 0, 1, 2.3 y 4.6 profundidades ópticas y a 100 m de profundidad para estimar la concentración de clorofila a. Las profundidades ópticas se determinaron usando información del perfilador PNF.
La definición de capa de mezcla (σt, pcm) sigue a Sprintall y Tomczak (1992), expresada según la siguiente ecuación:

donde σt,0 es el valor de σt en la superficie, ΔT es el cambio de temperatura (en nuestro caso –0.5 °C) y δσt /δT es el coeficiente de expansión térmica.
La capa isotermal (CIT) se calculó según Kara et al. (2000), expresada como:

Estos autores consideran la profundidad de 10 m como la temperatura superficial (SST) para eliminar el "efecto de piel", en nuestro caso ΔT = 0.5 °C.
La profundidad de la zona eufótica se obtuvo a partir de perfiles de luz del perfilador de fluorescencia natural y es la profundidad donde se alcanza el 1% de la luz en superficie.
Se usó un arreglo de trampas de sedimentación de deriva para muestrear flujos de partículas en los primeros 200 m de la columna de agua (estaciones D1 y D2, fig. 1). Se tomaron muestras a 50, 100 y 200 m, y en cada uno de los niveles se usaron cuatro trampas. La construcción, el armado, los procedimientos de limpieza y el cuidado de las trampas de sedimentación se realizaron siguiendo el protocolo del programa BATS (Bermuda Atlantic Time–series Study). Para recolectar las partículas se utilizaron filtros de membrana de policarbonato de 0.8 de tamaño de poro y 90 mm de diámetro. No se agregaron preservativos. Todo el zooplancton reconocible ("nadadores") se removió de los filtros. El carbono y nitrógeno orgánico particulado (COP y NOP) del material obtenido de las trampas se determinaron mediante un analizador de elementos CE440 (Leeman Labs).
El tiempo de residencia del COP (τCOP, en días) se calculó utilizando la ecuación propuesta por Eppley et al. (1983):
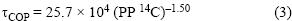
donde PP 14C es la producción primaria medida por el método de 14C (en mg C m–2 d–1). Dado que se ha encontrado que las mediciones de PP realizadas por el método de 14C tienen una buena correlación (r = 0.78 a 0.82) con mediciones estimadas por el método de fluorescencia natural (Chamberlin et al. 1990, Valdez–Holguín et at. 1995, García–Mendoza y Maske 1996, Aguirre–Gómez 2002), se utilizó esta última para el cálculo del tiempo de residencia.
La biomasa del fitoplancton se determinó mediante el contenido de clorofila a. Para esto, se filtró al menos 1 L de muestra en filtros GF/F. Para la extracción de clorofila se colocaron los filtros conteniendo la muestra en viales con 10 mL de acetona al 90% por 24 h, en un lugar fresco y oscuro (Venrick y Hayward 1984). La fluorescencia de las muestras se determinó utilizando un fluorímetro Turner modelo 111 siguiendo la técnica de Yentsch y Menzel (1963) y Holm–Hansen et al. (1965). La calibración se realizó con clorofila a pura (Sigma Chemical Co.).
Para calcular los montos de exportación total de MOP se consideró el área comprendida dentro de las estaciones de muestreo (128 × 80 km) y la MOP capturada por las trampas de sedimento a 200 m.
Para examinar las diferencias entre las estaciones del año se usó la estadística no paramétrica: la U de Mann–Whitney y el análisis de varianza de Kruskal–Wallis (Siegel 1986).
RESULTADOS
Condiciones oceanográficas
Un resumen de los datos de las condiciones hidrográficas que prevalecieron durante el periodo de estudio se presenta en la tabla 1. Durante el otoño de 1994 la profundidad de la capa isotermal y la de la capa de mezcla fueron muy similares y someras, mientras que la zona eufótica fue ligeramente más profunda. En la primavera de 1995 la capa isotermal disminuyó ligeramente, la profundidad de la capa de mezcla se incrementó y fue la más profunda de todo el periodo de muestreo, mientras que la profundidad de la zona eufótica disminuyó, alcanzando los menores valores de todo el periodo de muestreo. Para el verano de 1995 disminuyó la capa isotermal, siendo la más somera de todo el periodo de estudio, y la profundidad de la capa de mezcla y la de la zona eufótica aumentaron, siendo las más profundas de todo el periodo de muestreo. En otoño de 1995 la capa isotermal fue la más profunda de todo el periodo de estudio, mientras que comparada con la estación previa, la profundidad de la capa de mezcla aumentó y la zona eufótica disminuyó a valores intermedios a los observados durante otoño de 1994 y primavera de 1995. En verano ocurrió el fenómeno opuesto al invierno: la capa isotermal y la de mezcla fueron delgadas o inexistentes. No se encontró ninguna correlación clara entre la profundidad de la capa de mezcla o de la capa isotermal con la de la zona eufótica.
La variabilidad hidrográfica de la capa superior del océano presentó cambios notables de un crucero a otro, tanto en la temperatura como en la salinidad superficial. Los valores que se discuten aquí son los promedios de todos los lances realizados por crucero, a la profundidad de 5 m. La temperatura más baja se presentó durante la primavera de 1995 y la más alta en el verano de 1995. Sobresale la gran diferencia de 4.72 °C que se observa entre otoños sucesivos. En el otoño de 1994 el promedio de la temperatura superficial fue de 25.55 ± 0.10 °C, mientras que en el de 1995 disminuyó a 20.76 ± 0.18 °C. Debido a la poca variabilidad de las masas de agua superficiales, la salinidad también presentó una variabilidad mucho menor que la temperatura. El valor más alto se registró en el otoño de 1994 y el menor durante la primavera de 1995 (tabla 1).
Biomasa fitoplanctónica
La información obtenida en las cuatro estaciones del año muestreadas se resume en la tabla 2. En la figura 2 se presenta la distribución de la concentración de la clorofila a integrada (0–100 m).
Durante el otoño de 1994 (fig. 2a) los valores más altos de clorofila a se registraron en las estaciones más norteñas y las concentraciones fueron comparativamente más elevadas en la costa de Baja California que en la costa de Sonora. Estadísticamente (P > 0.05) no hubo diferencia significativa entre costas y se puede considerar como un área homogénea. En las estaciones de deriva los valores fueron similares a los de las estaciones del centro del Golfo de California; la concentración de clorofila a integrada de las estaciones de deriva equivale a la mitad del valor promedio en toda el área muestreada.
En la primavera de 1995 (fig. 2b) sobresalen valores muy altos de clorofila a en las estaciones 4 y 9; sin embargo, tomando en cuenta todas las estaciones de la red, estadísticamente (P > 0.05) no hubo diferencias entre las costas y el área se puede considerar homogénea. Como se observa en la tabla 2, la concentración promedio de clorofila a fue la más alta de las cuatro estaciones del año muestreadas, representando más del doble que la del verano. En las estaciones de deriva la concentración de la clorofila a fue parecida a la que se observó en las estaciones localizadas al norte y al este, pero muy diferente a la que ocurrió en la estación al sur. En general, se puede apreciar que la concentración de clorofila a que se encontró en las estaciones de deriva fue la mitad del valor promedio para toda el área muestreada.
Durante el verano de 1995 (fig. 2c) las concentraciones más altas de clorofila a se presentaron en la costa de Baja California y las más bajas en la zona central. Tomando en cuenta todas las estaciones de la red, estadísticamente (P < 0.05) no hubo diferencias entre las costas y, por lo tanto, el área se puede considerar homogénea. En las estaciones de deriva la concentración de clorofila a fue aproximadamente la mitad de la que se obtuvo como valor promedio para toda la red y se asemeja más a los valores que se encontraron en la costa de Sonora que a los que se encontraron en la región central del golfo.
Durante el otoño de 1995 (fig. 2d) las concentraciones más altas se encontraron en la costa de Sonora; el resto del área fue muy homogénea. En las estaciones de deriva los valores fueron los más altos del año; representaron casi el doble de los registrados en la red de estaciones, invirtiéndose el patrón que se observó en las otras estaciones del año, donde las concentraciones eran más altas en la red.
Producción primaria
La información de la producción primaria (PP) en las cuatro estaciones del año muestreadas se resume en la tabla 2. En la figura 3 se presenta la distribución de la concentración de PP; los valores se integraron de ~5 m a la base de la zona eufótica.
En el otoño de 1994 (fig. 3a) los valores más altos de PP se encontraron en el centro del golfo y los más bajos en la costa de Baja California. Estadísticamente (P > 0.05) no hubo diferencias entre las costas; sin embargo, considerando todas las estaciones de la red, el área no es homogénea. En las estaciones de deriva la tasa promedio fue ligeramente más alta que la observada en las estaciones que las rodean y que el valor promedio de toda la red.
Durante la primavera de 1995 (fig. 3b) se observaron los valores más altos de PP de las cuatro estaciones del año muestreadas. Los valores más altos de PP correspondieron a la costa de Sonora y los más bajos al centro del golfo. Igual que en el caso de la biomasa fitoplanctónica, también sobresalieron los valores altos de PP en las estaciones 4 y 9, indicando que la Bahía de Guaymas es una zona de alta productividad y biomasa fitoplanctónica durante esta época del año. No hubo diferencia estadística (P > 0.05) entre las costas, pero al considerar todas las estaciones de la red, omitiendo las estaciones de deriva, se encontró que el área no es homogénea. En las estaciones de deriva se encontró un valor más parecido a los del centro del golfo, más bajo que el valor promedio para toda la red.
Durante el verano de 1995 (fig. 3c) bajó la PP, encontrándose valores parecidos a los de otoño de 1994 (fig. 3a). Los valores más altos de PP se encontraron en la zona central del golfo, mientras que los más bajos ocurrieron en la costa de Sonora. Estadísticamente (P < 0.05) hubo diferencias entre las costas de Sonora y de Baja California, existiendo interacción entre la hora de muestreo y la costa. Considerando todas las estaciones de la red, se encontró que el área no es homogénea. La PP promedio en las estaciones de deriva fue muy parecida al valor promedio para toda la red de estaciones.
En el otoño de 1995 (fig. 3d) no se muestreó el transecto norte; los valores de PP en la región muestreada se elevaron ligeramente en relación con el verano. Los valores de PP mostraron el mismo patrón de comportamiento que durante el verano: los más altos se presentaron en el centro del golfo y los más bajos en la costa de Sonora. La PP promedio en la estación de deriva fue ligeramente más alta que el valor promedio para toda la red de estaciones.
Los valores de PP encontrados durante el verano fueron muy parecidos a los de otoño de 1994 y 1995, pero menor que la mitad de los encontrados durante la primavera (tabla 2). Considerando los valores de PP de cada una de las cuatro estaciones del año muestreadas, se encontró que existen diferencias estadísticas (P < 0.05) entre el otoño de 1994 y la primavera de 1995, y entre la primavera y el verano de 1995, pero no hay diferencias entre el otoño de ambos años y el verano.
Flujos de partículas
Tal como lo reportaron Lara–Lara et al. (2007), la variación estacional de los flujos de partículas medidos se caracterizó por un flujo máximo durante otoño y un mínimo durante el verano de 1995 (tabla 3, fig. 4). La distribución de los flujos mostró dos patrones: uno caracterizado por flujos de partículas altos (primavera y otoño de 1995) y el otro coincidiendo con flujos bajos (otoño de 1994 y verano de 1995). En las épocas cuando los flujos fueron mayores la distribución del carbono y la del nitrógeno tuvieron el mismo comportamiento que el flujo total, mientras que durante la época de flujos bajos la distribución del carbono y del nitrógeno tendió a ser inversa a la del flujo total. Se observaron diferencias estadísticamente significativas (P > 0.05) entre los flujos de todas las estaciones del año.
En general, se observó que los mayores cambios en los flujos ocurrieron entre los 50 y 100 m de profundidad, aunque sólo fueron estadísticamente significativos (P > 0.05) durante la primavera; con los flujos entre 50 y 200 m sucede lo mismo que el caso anterior. Entre los flujos de 100 y 200 m no hubo diferencias estadísticamente significativas (P < 0.05) en ninguna estación del año.
El valor promedio de la razón C:N se mantuvo muy homogéneo a través de todo el periodo de estudio, independientemente de la profundidad, variando entre 9.0 y 13.8 (tabla 3), excepto dos valores muy altos a 100 y 200 m de profundidad durante el otoño de 1994 que parecen estar fuera de orden.
El tiempo de residencia del carbono orgánico particulado en la columna de agua varió entre 3.7 y 11.2 d (tabla 2), correspondiendo el menor tiempo a la primavera y el mayor al verano de 1995.
El porcentaje de PP diaria exportada de los primeros 200 m de la columna de agua varió entre 3.6% y 8.9% (tabla 2), correspondiendo a la primavera y el otoño de 1995, respectivamente.
DISCUSIÓN
En el Golfo de California se distinguen claramente dos estaciones, una fría y otra cálida. Las series de tiempo de las imágenes del satélite Coastal Zone Color Scanner (CZCS) muestran una variación estacional muy clara de la concentración de pigmentos en el golfo, con concentraciones máximas entre noviembre y mayo (invierno), y mínimas entre junio y octubre (verano) (Santamaría–del–Ángel et al. 1994, Thunell et al. 1994). Esta situación se refleja en nuestros resultados (tabla 2), ya que los valores máximos de la concentración de clorofila a se presentaron en primavera, que corresponde al final del periodo frío, mientras que durante el periodo cálido (verano y otoño) la concentración de clorofila a disminuyó casi a la mitad de la que había en primavera, siendo muy parecidas las concentraciones del verano y el otoño. Este mismo patrón de estacionalidad se puede observar para PP, que también es más baja durante el verano que durante el otoño. Como resultado de la estratificación, en la capa superficial de agua ocurre un agotamiento de nutrientes (Thunell et al. 1993) que se refleja en una menor tasa de PP. La diferencia de la biomasa de fitoplancton entre el verano y el otoño no es tan marcada debido a que durante el verano la profundidad de la zona eufótica es mayor que durante el otoño, por lo que los valores tienden a ser similares (Álvarez–Borrego y Lara–Lara 1991), como se aprecia en la tabla 1.
Se ha demostrado claramente en varios estudios con trampas de sedimento que el flujo de la MOP está relacionado con la producción superficial, y que los flujos varían con los cambios diarios y estacionales en la productividad (Thunell et al. 1993, 1994; Lange et al. 1994). En la región central del Golfo de California existen marcadas diferencias estacionales en los flujos del material particulado (Lara–Lara et al. 2007); sin embargo, éstas se pueden agrupar claramente en dos periodos coincidiendo con las dos estaciones del año que señalan Santamaría–del–Ángel et al. (1994) y Thunell et al. (1994): la época fría (noviembre a mayo), con concentraciones máximas de pigmentos y una alta PP, y la época calida (junio a octubre), con concentraciones mínimas de pigmentos y baja productividad.
Durante el otoño de 1994 tanto los flujos como sus perfiles verticales fueron similares a los observados en el verano de 1995 y difieren marcadamente de los observados en el otoño de 1995, cuando los flujos totales fueron entre cinco y seis veces mayores y los de carbono y nitrógeno de dos a tres veces mayores (tabla 3, fig. 4). También se observó que en el otoño de 1994 las temperaturas superficiales fueron ~4.8 °C mayores que en la misma epoca del año siguiente (tabla 1). Esto sugiere que en 1994 las condiciones de verano se prolongaron hasta el otoño. El incremento de temperatura, consistente con un cambio hidrográfico (Soto–Mardones 1997), también fue acompañado de la disminución de la bio–masa fitoplanctónica, de la PP y del flujo de MOP (García–Pámanes 1999), lo que sugiere que los flujos durante el otoño de 1994 fueron inusuales para esta época.
En todos los cruceros, el flujo vertical de la MOP disminuyó rápidamente con la profundidad (fig. 4). Los mayores cambios ocurrieron entre los 50 y 100 m. Como se observa en la tabla 1, las profundidades promedio de la zona eufótica, la capa de mezcla y la capa isotermal no excedieron generalmente de los 50 m. Esto contribuye a que la MOP se mantenga más tiempo en la capa superficial, dando oportunidad de que el material sea consumido.
La disminución del flujo de la MOP entre 50 y 100 m (fig. 4) es atribuible al consumo de los herbívoros y a la remineralización del carbono orgánico a carbono inorgánico disuelto (Bender et al. 1992). Es notable la disminución tan grande de la MOP que existe durante la primavera en los primeros 100 m de la columna de agua, donde más del 50% de la materia es removida; esto puede deberse a que en esta época se presentó la mayor biomasa del zooplancton >202 µm en todo el año y consumió el 63% de la PP diaria (García–Pámanes 1999). Siguiendo con la figura 4, los flujos de MOP entre los 100 y 200 m se mantuvieron casi constantes. Esto indica que por debajo de los 100 m el consumo y la degradación fueron mínimos.
La razón C:N de Redfield para la MOP es de 6.6 (Bender et al. 1992), mientras que las razones que se encontraron en la MOP recolectada con las trampas de sedimento variaron entre 9.0 y 13.8 (tabla 3), un poco más altas que la de Redfield. Éstas son más similares al valor de 10 reportado para el detritus retrabajado de la zona costera por Walsh et al. (1981). La desviación de la razón de Redfield puede deberse a dos causas: al crecimiento exponencial del fitoplancton bajo condiciones de limitación de nutrientes que incrementa las razones C:N y C:P (Schneider et al. 2003, Christian 2005), y a la descomposición preferencial de compuestos orgánicos que contienen nitrógeno y fósforo (Knauer et al. 1979, Wakeham y Lee 1993). La alta razón C:N encontrada a todas las profundidades (tabla 3) significa que la materia orgánica está siendo remineralizada rápidamente dentro de los primeros 50 m de la columna de agua y que el COP que es exportado al fondo está constituido por material altamente "refractario".
El tiempo de residencia del COP en la columna de agua fue relativamente corto (3.7–11.2 d, tabla 2). Los tiempos más largos correspondieron a la época calida de baja productividad y mayor estabilidad en la columna de agua, y los más cortos a la época fría de alta productividad y mayor dinámica (Lara–Lara et al. 2007). Estos resultados concuerdan con los reportados por Eppley et al. (1983) para la Bahía de Monterey (California), la Bahía del Sur de California y la Cuenca de Panamá. Riebesell (1992) menciona también que tiempos de residencia cortos pueden estar relacionados con la intensificación de los vientos y la profundización de la capa de mezcla, lo cual impediría la formación de burbujas en los agregados y resultaría en altas tasas de sedimentación. Esta situación se aprecia también en nuestros resultados.
Los tiempos de residencia estimados concuerdan con la variabilidad temporal en la magnitud de los flujos del COP por debajo de la zona eufótica observados en este estudio. Parte de esa variabilidad parece estar relacionada con los cambios estacionales, pero otra parte (como la de otoño de 1994) parece relacionarse con anomalías de baja frecuencia en la temperatura superficial del mar, tal como lo reportan Eppley et al. (1983) para la costa de California.
La trayectoria de las trampas durante su deriva parece estar influida por los remolinos que se han reportado en la zona (Figueroa et al. 2003). Durante el otoño de 1994 y el verano y otoño de 1995 las trampas siguieron una trayectoria curva, del sureste hacia el noroeste (ciclónica). En contraste, durante la primavera de 1995 se observó el movimiento opuesto, del noroeste hacia el sureste (anticiclónico). Esto es consistente con lo reportado por Figueroa et al. (2003) para la Cuenca de Guaymas. Asimismo, las velocidades de desplazamiento variaron entre 0.07 y 0.21 m s–1, las cuales concuerdan con las reportadas dentro de remolinos (Bray 1988, Fernández–Barajas et al. 1994). La mayor velocidad se presentó en primavera de 1995 y la más baja en otoño de 1995. La distancia que puede recorrer la MOP (24.502–140.111 km) durante su tiempo de residencia dentro de los primeros 200 m de la columna de agua, fácilmente permite el transporte de MOP de una costa a otra del Golfo de California. El exceso de producción en la costa de Sonora puede ser acarreado a un área más pobre, tendiendo a homogeneizar el área y contribuyendo al enriquecimiento de toda la cuenca. Esto, a su vez, propicia que los organismos encuentren alimento disponible en toda la región.
Bajo esta perspectiva, la hipótesis de Thunell et al. (1993) de que los flujos de la MOP son uniformes en toda la región central del Golfo de California parece correcta; sin embargo, los resultados del presente estudio, obtenidos con base en trampas lagrangianas, muestran variabilidad en los flujos de MOP en diferentes partes de la cuenca y en diferentes épocas. Por ejemplo, la estación donde se soltaron las trampas de sedimento en el muestreo de otoño de 1994 se localizó entre el transecto central y el sureño (estación D1, fig. 1), y el flujo total de MOP fue de 227.91 ± 22.31 mg m–2 d–1, mientras que en el muestreo de otoño de 1995 la estación se localizó entre el transecto norteño y el central (estación D2, fig. 1), y el flujo total fue de 1720.51 ± 183.54 mg m–2 d–1 (tabla 3). Esto podría indicar que los flujos pueden variar tanto estacional como latitudinalmente.
Algunos autores han estimado que el porcentaje de PP exportado de los 200 m superficiales del océano varía de menos de 5% a 10% de la PP neta (Berger et al. 1989, Wakeham y Lee 1993). Los resultados que aquí se reportan están dentro de este mismo orden de magnitud (tabla 2). Sorprendentemente las mayores tasas de exportación ocurrieron durante el verano, cuando tanto la biomasa del fitoplancton, la PP y la biomasa del zooplancton (8.80 mg m–3, peso seco; García–Pámanes 1999) fueron las más bajas del año. Thunell et al. (1993) también notaron este fenómeno aunque no presentan una explicación. Este comportamiento pudiera deberse a que en verano se presentan las tasas más altas de pastoreo por el mesozooplancton en el año. García–Pámanes (1999) reportó valores de 2.27 ± 1.29 d–1 que implican la remoción del 91% de PP diaria. Entonces, el mayor tiempo de residencia del COP en las aguas superficiales y las altas tasas de pastoreo permiten un mejor aprovechamiento del alimento disponible, lo cual se traduce en la formación de una mayor cantidad de heces fecales, las que a su vez transportan una proporción más alta de COP hacia el piso oceánico.
Hernández–Ayón et al. (2007) reportaron que la región central del Golfo de California (Cuenca de Guaymas) actúa como sumidero de CO2 y que la actividad biológica pudo ser el proceso causante de la disminución de CO2 total en la región al sur de las islas. En el caso del presente estudio, si se supone que el flujo de MOP es homogéneo en toda la cuenca (Thunell et al. 1993), entonces se pueden usar los flujos medidos para estimar el comportamiento de la cuenca y conocer la cantidad de carbono que se exporta de los 200 m superiores hacia el fondo (tabla 4). En la epoca fría es cuando mayor cantidad de materia se exporta (193,095 ± 41,181 t C época–1), mientras que en la epoca cálida se exporta un 27% menos (100,620 ± 59,985 t C época–1). Cabe destacar que la época fría dura dos meses más que la cálida. En promedio, en toda el área de estudio se exportan casi 300,000 t C año–1. Esta información aunada a la conclusión de Hernández–Ayón et al. (2007) permite suponer que la Cuenca de Guaymas se comporta como sumidero de carbono durante el verano. Esta exportación de carbono equivale aproximadamente al total del monóxido de carbono que en 1999 emitieron a la atmósfera conjuntamente los estados de Baja California, Baja California Sur y Sonora (INE–SEMARNAT 2006).
Sin embargo, hay que señalar que los resultados de Hernández–Ayón et al. (2007) se basaron en un solo crucero en septiembre de 1996. Como se sabe, las regiones costeras pueden variar entre fuente y sumidero de carbono estacional–mente o estar en balance durante el ciclo anual, dependiendo de la magnitud de los procesos biológicos, químicos y físicos. De tal manera, para conocer si la región de estudio se comporta como una fuente o sumidero durante todo el año, se tendría que conocer, además de los flujos verticales de carbono, los flujos océano–atmósfera de carbono durante todo el año para estimar los balances anuales.
AGRADECIMIENTOS
El primer autor contó con una beca del Consejo Nacional de Ciencia y Tecnología (CONACYT) durante su estancia en el Centro de Investigación Científica y de Educación Superior de Ensenada (CICESE). El segundo y el tercer autor son miembros del Sistema Nacional de Investigadores. El tercer autor recibió apoyo del CONACYT a través de los proyectos T9201–1111 y SEP–2004–CO1–45813. Se agradece a E Valdez–Holguín su apoyo en el trabajo de campo y laboratorio, a los capitanes y las tripulaciones de los B/Os Francisco de Ulloa (CICESE) y El Puma (UNAM) su colaboración, y a E González–Rodríguez y MA Cosío–López su ayuda en la elaboración de las series de tiempo y figuras.
REFERENCIAS
Aguirre–Gómez R. 2002. Primary production in the southern Gulf of Mexico estimated from solar–stimulated natural fluorescence. Hidrobiológica 12: 21–28. [ Links ]
Álvarez–Borrego S, Schwartzlose R. 1979. Water masses of the Gulf of California. Cienc. Mar. 6: 43–63. [ Links ]
Álvarez–Borrego S, Lara–Lara JR. 1991. The physical environment and primary productivity of the Gulf of California. In: Dauphin JP, Simoneit B (eds.), The Gulf and Peninsular Provinces of the Californias. Am. Assoc. Petrol. Geol. Mem. 47, pp. 555–567. [ Links ]
Badan–Dangon A, Koblinsky CJ, Baumgartner T. 1985. Spring and summer in the Gulf of California: Observations of surface thermal patterns. Oceanol. Acta 8: 13–22. [ Links ]
Bender M, Ducklow H, Kiddon J, Marra J, Martin J. 1992. The carbon balance during the 1989 spring bloom in the North Atlantic Ocean, 47°N, 20°W. Deep–Sea Res. 39: 1707–1725. [ Links ]
Berger WH, Smetacek VS, Wefer G. 1989. Ocean productivity and paleoproductivity: A review. In: Berger WH, Smetacek VS, Wefer G (eds.), Productivity of the Ocean: Present and Past. John Wiley, New York, pp. 1–34. [ Links ]
Boyd P, Newton P. 1995. Evidence of the potential influence of planktonic community structure on the interannual variability of particulate organic carbon flux. Deep–Sea Res. I 42: 619–639. [ Links ]
Bray NA. 1988. Thermohaline circulation in the Gulf of California. J. Geophys. Res. 93: 4993–5020. [ Links ]
Chamberlin WS, Booth CB, Kiefer DA, Morrow JH, Murphy RC. 1990. Evidence for a simple relationship between natural fluorescence, photosynthesis and chlorophyll in the sea. Deep–Sea Res. 37: 951–973. [ Links ]
Christian JR. 2005. Biogeochemical cycling in the oligotrophic ocean: Redfield and non–Redfield models. Limnol. Oceanogr. 50(2): 646–657. [ Links ]
Eppley RW, Renger EH, Betzer PR. 1983. The residence time of particulate organic carbon in the surface layer of the ocean. Deep–Sea Res. 30: 311–323. [ Links ]
Fernández–Barajas ML, Monreal–Gómez MD, Molina–Cruz A. 1994. Thermohaline structure and geostrophic flow in the Gulf of California during 1992. Cienc. Mar. 20: 267–286. [ Links ]
Figueroa JM, Marinone SG, Lavin MF. 2003. A description of geostrophic gyres in the southern Gulf of California. In: Velasco Fuentes OU, et al. (eds.), Nonlinear Processes in Geophysical Fluid Dynamics. Kluwer Academic Publishers, pp. 237–255. [ Links ]
García–Mendoza E, Maske H. 1996. The relationship of solar–stimulated natural fluorescence and primary productivity in Mexican Pacific waters. Limnol. Oceanogr. 41: 1697–1710. [ Links ]
García–Pámanes J. 1999. Flujo de carbono dentro de la zona epipelágica de la región central del Golfo de California. Ph.D. thesis, Centro de Investigación Científica y de Educación Superior de Ensenada, Mexico, 163 pp. [ Links ]
Harding GC, Hargrave BT, Vass WP, Sheldon RW, Pearre S Jr. 1987. Vertical flux of particulate matter by sedimentation and zooplankton movements in St. Georges Bay, the southern Gulf of St. Lawrence. Fluxes of particulate matter across benthic boundaries. Biol. Oceanogr. 4: 323–357. [ Links ]
Hernández–Ayón JM, Zirino–Weiss A, Delgadillo–Hinojosa F, Galindo–Bect S. 2007. Carbono inorgánico disuelto en el Golfo de California en condiciones de verano. In: Hernández de la Torre B, Gaxiola Castro G (eds.), Carbono en Ecosistemas Acuáticos de México. Instituto Nacional de Ecología, SMARNAT, Centro de Investigación Científica y de Educación Superior de Ensenada, Mexico, pp. 45–57. [ Links ]
Holm–Hansen O, Lorenzen CJ, Holmes RW, Strickland DH. 1965. Fluorimetric determination of chlorophyll. J. Cons. Int. Explor. Mer 30: 3–15. [ Links ]
INE–SEMARNAT. 2006. Inventario Nacional de Emisiones de México, 1999. Mexico, 377 pp. [ Links ]
Kara AB, Rochford PA, Hurlburt HE. 2000. An optimal definition for ocean mixed layer depth. J. Geophys. Res. 105: 16803–16821. [ Links ]
Knauer GA, Martin JH, Bruland KH. 1979. Fluxes of particulate carbon, nitrogen, and phosphorus in the upper water column of the northeast Pacific. Deep–Sea Res. 26A: 97–108. [ Links ]
Lange CB, Treppke UF, Fischer G. 1994. Seasonal diatom fluxes in the Guinea Basin and their relationships to trade winds, hydrography and upwelling events. Deep–Sea Res. I 41(5/6): 859–878. [ Links ]
Lara–Lara JR, García–Pámanes J, Bazán–Guzmán C. 2007. Flujo vertical de materia orgánica particulada en la región central del Golfo de California. In: Hernández de la Torre B, Gaxiola Castro G (eds.), Carbono en Ecosistemas Acuáticos de México. Instituto Nacional de Ecología, SEMARNAT, Centro de Investigación Científica y de Educación Superior de Ensenada, Mexico, pp. 383–395. [ Links ]
Longhurst A, Williams R. 1992. Carbon flux by seasonal vertical migrant copepods is a small number. J. Plankton Res. 14: 1495–1509. [ Links ]
Maluf LY. 1983. Physical oceanography. In: Case TJ, Cody ML (eds.), Island Biogeography of the Sea of Cortez. Univ. California Press, Berkeley, pp. 26–45. [ Links ]
Miquel JC, Fowler SW, la Rosa J, Buat–Menard P. 1994. Dynamics of the downward flux of particles and carbon in the open northwestern Mediterranean Sea. Deep–Sea Res. I 41: 243–261. [ Links ]
Peinert R, von Bodungen B, Smetacek V. 1989. Food web structure and loss rate. In: Berger WH, Smetacek SV, Wefer G (eds.), Productivity of the Ocean: Present and Past. John Wiley, New York, pp. 35–48. [ Links ]
Riebesell U. 1992. The formation of large marine snow and its sustained residence in surface waters. Limnol. Oceanogr. 37: 63–76. [ Links ]
Roden GI, Groves GW. 1959. Recent oceanographic investigations in the Gulf of California. J. Mar. Res. 18: 10–35. [ Links ]
Santamaría–del–Ángel E, Álvarez–Borrego S, Muller–Karger FE. 1994. Gulf of California biogeografic regions based on coastal zone color scanner imagery. J. Geophys. Res. 99: 7411–7421. [ Links ]
Sarmiento JL, Siegenthaler U. 1992. New production and the global carbon cycle. In: Falkowski PG, Woodhead AD (eds.), Primary Productivity and Biogeochemical Cycles in the Sea. Plenum Press, New York, pp. 317–332. [ Links ]
Schneider B, Schlitzer R, Fischer G, Nothig EM. 2003. Depth–dependent elemental compositions of particulate organic matter (POM) in the ocean. Global Biogeochemical Cycles. 17(2): 1032–1047. [ Links ]
Siegel S. 1986. Estadística no paramétrica. Editorial Trillas, Mexico. [ Links ]
Smetacek V, von Brockel K, Zeitzschel B, Zenk W. 1978. Sedimentation of particulate matter during a phytoplankton spring bloom in relation to the hydrographical regime. Mar. Biol. 47: 211–226. [ Links ]
Soto–Mardones LA. 1997. Variabilidad espaciotemporal de la temperatura superficial del mar en el Golfo de California. M.Sc. thesis, Centro de Investigación Científica y de Educación Superior de Ensenada, Mexico, 108 pp. [ Links ]
Sprintall J, Tomczak T. 1992. Evidence of barrier layer in the surface layer of the tropics. J. Geophys. Res. 97, C5: 7305–7316. [ Links ]
Thunell RC, Pride C, Tappa E, Muller–Karger F. 1993. Varve formation in the Gulf of California: Insights from time series sediment trap sampling and remote sensing. Quat. Sci. Rev. 12: 451–464. [ Links ]
Thunell RC, Pride C, Tappa E, Muller–Karger F. 1994. Biogenic silica fluxes and accumulation rates in the Gulf of California. Geology 22: 303–306. [ Links ]
Torres–Orozco E. 1993. Análisis volumétrico de las masas de agua del Golfo de California. M.Sc. thesis, Centro de Investigación Científica y de Educación Superior de Ensenada, Mexico, 80 pp. [ Links ]
Valdez–Holguín JE, Lara–Lara JR. 1987. Primary productivity in the Gulf of California: Effects of the El Niño 1982–1983 event. Cienc. Mar. 13(2): 34–50. [ Links ]
Valdez–Holguín JE, Gaxiola–Castro G, Cervantes–Duarte R. 1995. Primary productivity in the Gulf of California, calculated from the relationship between superficial irradiance and chlorophyll in the euphotic zone. Cienc. Mar. 21: 311–329. [ Links ]
Venrick EL, Hayward TL. 1984. Determining chlorophyll on the 1984 CalCOFI surveys. CalCOFI Rep. 25: 74–79. [ Links ]
Wakeham SG, Lee S. 1993. Production, transport, and alteration of particulate organic matter in the marine water column. In: Engel MH, Macko SA (eds.), Organic Geochemistry. Plenum Press, New York, pp. 145–169. [ Links ]
Walsh JJ, Rowe GT, Iverson RI, McRoy CP. 1981. Biological export of shelf carbon is a sink of global CO2 cycle. Nature 291: 196–201. [ Links ]
Yentsch CS, Menzel D. 1963. A method for the determination of phytoplankton chlorophyll and phaeophytin by fluorescence. Deep–Sea Res. 10: 221–231. [ Links ]
Zeitzschel B. 1969. Primary productivity in the Gulf of California. Mar. Biol. 3: 201–207. [ Links ]
*Descargar versión bilingüe (Inglés–Español) en formato PDF.

