










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkCiencias marinas
versão impressa ISSN 0185-3880
Cienc. mar vol.35 no.4 Ensenada Dez. 2009
Artículos de investigación
Occurrence of a subsurface anticyclonic eddy, fronts, and Trichodesmium spp. over the Campeche Canyon region, Gulf of Mexico*
Ocurrencia de un giro anticiclónico, frentes y Trichodesmium spp. sobre la región del Cañón de Campeche, Golfo de México
J Aldeco1, 3, MA Monreal–Gómez2, M Signoret1, DA Salas–de León2*, DU Hernández–Becerril2
1 Departamento El Hombre y su Ambiente, Universidad Autónoma Metropolitana–Xochimilco, Calzada del Hueso 1100, Col. Villa Quietud, 04960 México DF, México.
2 Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México, Circuito Exterior S/N, Cd. Universitaria, 04510 México DF, México. * E–mail: salas@mar.icmyl.unam.mx
3 Posgrado en Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México, Circuito Exterior S/N, Cd. Universitaria, 04510 México DF, México.
Recibido en marzo de 2009.
Aceptado en septiembre de 2009.
Abstract
Eddies and fronts created by geostrophic balance between counter–rotating currents are hydrodynamic processes that concentrate or dilute plankton and suspended material by physical processes of accumulation, retention, or dispersion. Over Campeche Canyon, in the southern Gulf of Mexico, observations revealed an anticyclonic eddy, a cyclonic current off the continental shelf, and an associated thermal front in their common boundary. Along this front there was high Trichodesmium spp. occurrence. Three water masses were found in the upper 120 m layer: Caribbean Tropical Surface Water (CTSW), Caribbean Subtropical Underwater (CSUW), and Gulf Common Water (GCW). The frontal zone between the warm core and cyclonic circulation was observed below the hot summertime mixed layer. The presence of Trichodesmium spp. was associated with the front, indicating its influence on the distribution of this diazotrophic cyanobacterium that enters the Campeche Canyon region via modified CTSW. The associated phytoplankton was dominated by tropical oceanic species of diatoms and dinoflagellates, especially species of Ceratium.
Key words: anticyclonic eddy, fronts, Trichodesmium, Campeche Canyon, Gulf of Mexico.
Resumen
Los giros y frentes creados por balance geostrófico entre corrientes que rotan en sentido contrario son procesos hidrodinámicos que concentran o diluyen plancton y material en suspensión mediante procesos físicos de acumulación, retención o dispersión. Sobre la región del Cañón de Campeche, en el sur del Golfo de México, se ha observado un giro anticiclónico, una corriente ciclónica frente a la plataforma continental y un frente térmico asociado que se presenta en la frontera común. A lo largo de este frente hay gran abundancia de Trichodesmium spp. En la capa superior de 120 m se identificó la presencia de tres masas de agua: Agua Tropical Superficial del Caribe (CTSW), Agua Subtropical Subsuperficial del Caribe (CSUW) y Agua Común del Golfo (GCW). La zona frontal entre el núcleo cálido y la circulación ciclónica se observó por debajo de la cálida capa de mezcla de verano. La mayor abundancia de Trichodesmium spp. estuvo asociada al frente que mostró su influencia sobre la distribución de esta cianobacteria diazotrófica que llega a la región del Cañón de Campeche como parte del CTSW modificada. El fitoplancton asociado estuvo dominado por especies oceánicas tropicales de diatomeas y dinoflagelados, particularmente especies del género Ceratium.
Palabras clave: giro anticiclónico, frentes, Trichodesmium, Cañón de Campeche, Golfo de México.
Introducción
El Cañón de Campeche y el Cañón De Soto son los dos rasgos fisiográficos más importantes del Golfo de México. Las condiciones hidrodinámicas en torno a estos rasgos batimétricos aún no han sido estudiadas en detalle. Se considera que estos cañones condicionan la hidrodinámica y la distribución de los sedimentos dando origen a micro ambientes (Escobar–Briones et al. 2008), y que juegan un papel importante en la estructura de las corrientes a nivel regional (Salas–de León et al. 2004). Por otro lado, los giros y frentes formados por balance geostrófico entre corrientes que giran en sentido contrario, son procesos hidrodinámicos que concentran o diluyen plancton y material en suspensión mediante procesos físicos de acumulación, retención o dispersión (Franks 1992). La generación de giros subsuperficiales, sus corrientes y los frentes asociados que se forman por su interacción entre el talud continental y los cañones en el Golfo de México, es un tema de especial interés debido a que son fenómenos que pueden explicar la ocurrencia de intensas corrientes subsuperficiales poco comunes que se observan en el Golfo de México (Oey y Zhang 2004), así como de los filamentos de fitoplancton que se han observado asociados a estas corrientes.
En este estudio se analiza la relación existente entre los procesos hidrodinámicos de los giros y frentes creados por balance geostrófico entre corrientes que se desplazan en sentido contrario, y la abundancia de Thichodesmium y fitoplancton observados sobre la región del Cañón de Campeche.
Área de estudio
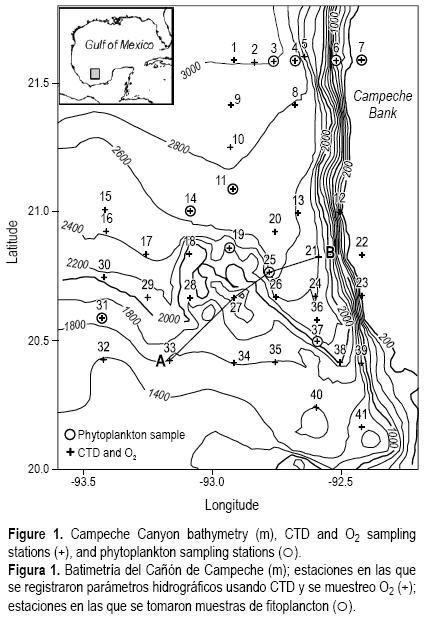
El Cañón de Campeche es una zona de transición en el sur del Golfo de México. Se considera que es el resultado de procesos tectónicos adyacentes (Antoine y Bryant 1968). El cañón es un área activa en cuanto a exportación de carbono biogénico (Escobar–Briones et al. 2008). Es una región pequeña y topográficamente compleja localizada en el noreste de la Bahía de Campeche (parte occidental del Banco de Campeche) en el sur del Golfo de México; se extiende desde aproximadamente 160 m de profundidad en el borde del talud continental hasta unos 2800 m en su punto más profundo (fig. 1), y tiene 125 km de largo y 30 km de ancho en la parte central (Creager 1958).
Las masas de agua superficiales y subsuperficiales en el Golfo de México son tanto de origen local como externo. El Agua Común del Golfo (GCW; T ~ 22.5°C y 36.3 < S < 36.4) (Vidal et al. 1994) se forma localmente, mientras que el Agua Tropical Superficial del Caribe (CTSW; T > 28°C y S > 36.4) se forma fuera del golfo y fluye hacia él en la capa superficial de la Corriente de Yucatán. Debajo de esta masa de agua se encuentra el Agua Subtropical Subsuperficial del Caribe (CSUW; 22.2 < T < 26°C y 36.4 < S < 36.7) (Schroeder et al. 1974). El CTSW incrementa su temperatura y salinidad conforme se desplaza hacia el oeste del Banco de Campeche (Nowlin 1972, Schroeder et al. 1974, Furnas y Smayda 1987). Esta masa de agua superficial es empujada por el viento. El CSUW entra con el flujo principal de la Corriente de Lazo, una parte de ella se distribuye al oeste del Golfo de México, mientras que otra parte contribuye en gran medida al volumen del giro anticiclónico que se desprende de la Corriente de Lazo, o sale del Golfo de México por el Estrecho de Florida.
En la parte sureste del área de estudio se ha observado un marcado frente térmico en el borde del Banco de Campeche que coincide con un domo de baja temperatura a profundidades medias cerca de la base de la capa de mezcla. Este domo de isotermas a lo largo del escarpe de Campeche, generalmente es una respuesta subsuperficial al fuerte gradiente de profundidad entre la parte externa de la plataforma continental y el cañón. El domo de agua intermedia en el escarpe de Campeche y en la cabeza del cañón, como se ha podido deducir de los bajos valores de oxígeno presentes, juega un papel importante en la distribución de estructuras biológicas (Salas–de León et al. 2004, Escobar–Briones et al. 2008).
Existen pocos estudios concernientes a las comunidades planctónicas sobre el Cañón de Campeche. Hernández–Becerril et al. (2008) encontraron una predominancia del cocolitofórido Emiliania huxleyi y de la diatomea Nitzschia bifurcata. En verano la mayor cantidad de biomasa zooplanctónica se encuentra debajo de la termoclina, en la capa superior de 60 m de profundidad (Mojica–Ramírez 2003), mientras que en invierno, durante la temporada de tormentas invernales localmente conocidas como "Nortes", las comunidades de zoo–plancton estuvieron dominadas por copépodos y la mayor abundancia de zooplancton se encuentra en torno a la termoclina, esto como resultado del incremento del espesor de la capa de mezcla (Ramírez–Cruz 2005).
Materiales y métodos
Los datos hidrográficos y muestras de fitoplancton incluyendo tricomas de Trichodesmium se obtuvieron desde el B/O Justo Sierra de la Universidad Nacional Autónoma de México, del 6 al 15 de agosto de 1999. Los datos de conductividad, temperatura y presión se registraron en 41 estaciones mediante un CTD Neil Brown Mark III. La salinidad y densidad potencial (σθ) fueron calculadas de acuerdo con Fofonoff y Millard (1983). Para la toma de muestras a diferentes profundidades para el análisis de oxígeno y salinidad se usó una roseta General Oceanic equipada con botellas Niskin. Los datos de salinidad utilizados para calibrar las mediciones de este parámetro con el CTD se obtuvieron con un salinómetro de inducción. El contenido de oxígeno se midió inmediatamente después de tomadas las muestras de agua de las botellas de la roseta, mediante un oxímetro YSI 5000 (±18.75 µmol L–1) previamente calibrado en condiciones atmosféricas, de acuerdo con las indicaciones del fabricante.
Las masas de agua fueron identificadas usando un diagrama T–S. Con el fin de analizar la estructura hidrográfica se estableció una sección vertical AB para analizar la temperatura en la capa superior de 250 m. La estructura horizontal debajo de la capa de mezcla fue investigada mediante la topografía de la superficie isoterma de 22°C, y con la distribución de la temperatura, salinidad y topografía dinámica a los 50 m de profundidad. El patrón de la circulación fue inferido de la topografía dinámica.
Las muestras de fitoplancton, incluyendo los tricomas de Trichodesmium, fueron recolectadas en 10 puntos de muestreo (fig. 1) usando una red con luz de malla de 54 um mediante arrastres verticales desde 120–100 m de profundidad hasta la superficie. A excepción de una muestra, todas se tomaron a más de 200 km de la costa y a una profundidad total mayor a 2000 m. La estación 7 se ubicó sobre el Banco de Campeche a una profundidad total de 160 m. Las muestras se fijaron con formaldehído neutralizado al 4% y se almacenaron para su análisis posterior. Los tricomas de Trichodesmium fueron cuantificados directamente en el microscopio y referidos al volumen de agua filtrado. La densidad de tricomas por unidad de volumen de agua (tricomas L–1) se calculó considerando el área de la boca de la red y la distancia de arrastre. La composición taxonómica de todo el fitoplancton se determinó de acuerdo con Tomas (1997).
Con el objeto de identificar semejanzas entre estaciones se construyó un dendograma con los datos oceanográficos a 50 m de profundidad (oxígeno disuelto, temperatura, salinidad, densidad potencial, topografía dinámica y abundancia de Trichodesmium). Para realizar el análisis de conglomerados (clusters) se usó el programa Plymouth Routines in Multivariate Ecological Research (PRIMER 6) (Clark y Gorley 2006).
Resultados
Hidrografía
Como se observa en el diagrama T–S de la capa superficial de 120 m en el área de estudio (fig. 2), el CTSW y el CSUW son modificadas en el Golfo de México. La masa de CTSW modificada se caracteriza por intervalos de temperatura de 28 < T < 30°C, y de salinidad de 36.4 < S < 36.8, mientras que la masa de CSUW modificada tiene 22 < T < 26°C y 36.4 < S < 36.6. También se puede observar en el diagrama T–S el GCW, de 36.2 < S < 36.4, y con un amplio intervalo de temperatura de 22 < T < 28°C (fig. 2).

La temperatura superficial en la región fue ~29.5°C, mientras que a 120 m de profundidad fue de 18.5°C. El piso de la capa de mezcla se localizó a ~35 m de profundidad. Por debajo de la termoclina, la forma de las isotermas revela una configuración típica de un núcleo calido cerca de la estación 27 (fig. 3). La salinidad varió de 36.5 en la superficie del mar a 36.4 a 120 m de profundidad y σθ de 23 kg m–3 en la superficie a 26 kg m–3 a 120 m. Por debajo del piso de la termoclina, a 50 m de profundidad, se observaron las mismas características hidrográficas hasta profundidades mayores a 200 m. De acuerdo con lo antes mencionado, la profundidad de 50 m fue adecuada para el análisis de masas de agua ya que la temperatura y la salinidad se ajustan a la distribución de las masas de agua subsuperficiales del golfo, y muestran mejor su estructura dinámica.

La topografía de la isoterma de los 22°C (fig. 4a) fue más profunda en el suroeste que en el sureste y con 90 m de profundidad en el centro del núcleo, lo cual evidencia el giro anticiclónico. Por debajo de la capa de mezcla la distribución horizontal de temperatura, salinidad y altura dinámica a 50 m de profundidad (fig. 4) muestra que el CSUW en el suroeste y norte del área de estudio estuvo circulando en dirección anticiclónica, mientras que el GCW circulaba en dirección ciclónica en el centro y el este del área de estudio. El CTSW modificada, debido a su temperatura, se localizó en la capa de mezcla de la zona de estudio (no mostrada).
En general los valores de oxígeno disuelto (no mostrados) decrecen hacia el sur, especialmente donde existe un fuerte gradiente de temperatura debido al afloramiento subsuperficial, cerca de la pared del Cañón de Campeche (fig. 4b). En la porción sureste del área de estudio (estación 41) se observaron valores mínimos de oxígeno disuelto (146 µmol L–1), y un máximo de 361 µmol L–1 en la estación 1. Los valores mínimos pueden estar relacionados con el mínimo semipermanente mostrado por Nowlin (1972).
En la parte sureste del área de estudio la distribución horizontal de la temperatura a 50 m de profundidad (fig. 4b) mostró un evidente gradiente térmico asociado al domo de las isotermas a lo largo del margen del escarpe de Campeche. Por otro lado, a lo largo del área de estudio la temperatura fue más de 5°C menor en el sureste que en el norte y suroeste (fig. 4b), mientras que las diferencias en salinidad fueron de sólo 0.3 (fig. 4c). En la zona frontal donde se unen las corrientes anticiclónica y ciclónica, los gradientes horizontales de temperatura y de salinidad fueron de 5 x 10–2 °C km–1 y 0.18 x 10–2 km–1, respectivamente. Las isotermas a 50 m de profundidad mostraron oscilaciones anticiclónicas en la región suroeste con temperaturas alrededor de los 26°C, mientras que en la región central–oriental fue evidente una circulación ciclónica con temperaturas por debajo de 24°C. Estas circulaciones forman un frente térmico en la frontera común debido a sus gradientes horizontales. La topografía dinámica a 50 m de profundidad (fig. 4d) muestra el patrón de circulación y pone en evidencia la zona frontal entre las oscilaciones ciclónica y anticiclónica. La máxima velocidad estimada fue de 22 cm s–1 en la parte central de la zona de estudio a aproximadamente 20.75°N.
Abundancia de Trichodesmium spp.
Se identificaron tres especies de Trichodesmium: T. erythraeum Ehrenberg ex Gomont, T. hildebrandtii Gomont, y T. thiebautii Gomont ex Gomont, de acuerdo a Sournia (1968). La abundancia de Trichodesmium spp. varió entre 1.5 y 175 tricomas L–1. Los valores más altos se encontraron en las estaciones 11, 19 y 25 (fig. 4d) y estuvieron asociadas con el frente térmico. Las abundancias bajas ocurrieron sobre la plataforma continental (estación 7) (tabla 1).
El dendrograma (fig. 5a) muestra la existencia de tres grupos de acuerdo a las abundancias de Trichodesmium en relación a las condiciones hidrográficas. Uno de los grupos corresponde a las estaciones donde se observó el frente geostrófico subsuperficial, donde los tricomas fueron más abundantes (estaciones 11, 14, 19 y 25). Las menores abundancias de tricomas estuvieron relacionadas con las condiciones hidrográficas que existen sobre las zonas someras del Banco de Campeche, lejos del frente (i.e., región norte, estación 7) (fig. 5b).

El fitoplancton asociado mostró 29 géneros y 66 especies identificadas (tabla 2). La mayoría de ellos se encontró en la zona frontal (estaciones 14 y 19). La composición taxonómica es característica de aguas oceánicas tropicales, especialmente la de dinoflagelados (Steidinger y Tangen 1997). Los dinoflagelados más frecuentes fueron Ceratium (16 especies), Ornithocercus (6 especies), y Dinophysis y Protoperidinium (5 especies cada una). Wood (1971) reportó para el Mar Caribe 47% de las especies identificadas en este estudio.
Discusión
El patrón de corrientes observado en el área de estudio pudo haber sido generado por al menos tres factores: (1) el rotacional negativo del esfuerzo del viento sobre el Golfo de México, el cual es anticiclónico en la parte noreste del golfo y ciclónico en la parte suroeste (Gutiérrez–de Velasco y Winant 1996); (2) la fricción de la corriente con el borde del talud continental que genera surgencias batistróficas (Hsueh y O'Brien 1971) y giros ciclónicos; y (3) el desprendimiento de los giros anticiclónicos de la Corriente de Lazo, los cuales migran hacia el oeste en el golfo (Vidal et al. 1994). Estudios efectuados con modelos de la interacción de los giros con el talud y/o las paredes verticales de las fronteras (Smith y O'Brien 1983, Shi y Nof 1993, Grimshaw et al. 1994, Nof 1999) muestran que los giros tienden a moverse hacia el norte por un efecto de imagen, hacia el sur debido a la advección inducida por el efecto p, y hacia el norte debido a la expulsión de masa hacia el sur desde el giro. Las características de las masas de agua en la zona de estudio son modificadas por la presencia de uno o más de estos procesos.
El anticiclón mexicano, como le llamaron Vázquez–de la Cerda et al. (2005) al giro previamente desprendido de la corriente de Lazo, algunas veces extiende sus meandros hacia el oeste del Banco de Campeche, y como lo establecen Nowlin y McLellan (1967), no se observa en la superficie. De acuerdo con las características de las masas de agua y de los patrones de circulación se cree que la capa de agua caliente isoterma que cubre esta agua cálida llega del Banco de Campeche.
Oey y Zhang (2004) mostraron la generación de corrientes y giros ciclónicos parásitos cuando un giro cálido choca con el talud continental, y la consecuente formación de un flujo subsuperficial intenso. Este fenómeno puede explicar la ocurrencia de la fuerte corriente y del frente observado entre la corriente ciclónica y el giro anticiclónico sobre el Cañón de Campeche y su influencia sobre la distribución de Trichodesmium spp.
Una de las principales aportaciones de esta investigación fue la observación de Trichodesmium spp. La cianobacteria diazotrófica Trichodesmium es un gran fitoplanctonte que es común en aguas oceánicas tropicales, y es el fijador de nitrógeno dominante a escalas regionales y globales (Westberry y Siegel 2006). Trichodesmium habita en aguas superficiales de mares tropicales oligotróficos, por lo que su distribución geográfica está gobernada por la distribución de las masas de agua superficiales. Sin embargo, también se le ha encontrado en aguas costeras y aguas eutróficas de mares tropicales (Capone et al. 1997).
En el Golfo de México varios estudios han documentado la presencia de Trichodesmium y su participación en la fijación de nitrógeno (e.g., Lenes et al. 2001, Hood et al. 2004, Walsh et al. 2006). Se han reportado florecimientos esporádicos de T. erythraeum desde 1950, los cuales pueden formar filamentos pardos floculentos sobre la superficie del mar. En años recientes las imágenes de satélite han permitido observar desde el espacio los florecimientos de Trichodesmium spp. (Westberry et al. 2005), que comúnmente se ha observado en el Golfo de México y es muy variable (Hood et al. 2004). En ambientes subtropicales es abundante en aguas cálidas oceánicas estratificadas (Karl et al. 1995, Tyrrell et al. 2003), pero no ha sido reportado previamente para la región del Cañón de Campeche.
La abundancia de tricomas de Trichodesmium observada en este estudio fue relativamente baja comparada con datos de otras regiones como la zona comprendida entre Carolina del Sur y Florida (Subramaniam et al. 2002), o a escala global (Westberry y Siegel 2006). De acuerdo a las categorías de concentración de filamentos propuesta por Carpenter (1983) para un océano global (0.1, 1, 10, 100 y 1000 tricomas L–1), nuestros valores estuvieron en las 2a, 3a y 4a categorías, pero principalmente en la 3a. Karl et al. (1995) reportaron un promedio de 46 tricomas L–1 cerca de Hawaii, que es del mismo orden encontrado en este estudio.
Las cianobacterias Trichodesmium diazotróficas son un grupo funcional biogeoquímico con una distribución determinada principalmente por fenómenos físicos de mesoescala (Hood et al. 2006). Las corrientes, frentes y giros que forman la mesoescala oceánica, algunas veces referida como "clima interno del océano", son rasgos muy energéticos presentes en la circulación oceánica. Las señales biológicas se ven incrementadas en la vecindad de discontinuidades físicas como los frentes, tal y como se ha observado en este estudio con el frente térmico y la abundancia de Trichodesmium spp.
Nuestros resultados sugieren que la zona frontal detectada sobre el Cañón de Campeche puede promover la agregación de tricomas. De acuerdo con Mann y Lazier (1991), es de esperar que cualquier organismo con suficiente flotabilidad para resistir el hundimiento de agua se concentre en el frente. Franks (1992) demostró que el incremento de la biomasa planctónica en frentes puede resultar únicamente de procesos físicos en zonas de retención y acumulación.
Sobre el Cañón de Campeche, el CTSW ocupó la capa de mezcla de 35 m de profundidad. Esto sugiere que los tricomas de Trichodesmium estaban presentes en esta capa antes de su agregación en el frente. En la literatura la localización vertical de Trichodesmium se reporta dentro de la capa de mezcla, especialmente cuando la capa de mezcla es delgada (<50 m de profundidad), con gran intensidad de luz y baja concentración de nutrientes (Letelier y Karl 1998, Lenes et al. 2001, Hood et al. 2004). Carpenter et al. (2004) señalaron que los 50 m superficiales contenían 94% del total de Trichodesmium. De acuerdo con Villareal y Carpenter (1990), Trichodesmium puede ser capaz de controlar su flotabilidad cambiando el contenido de gas en sus vesículas, por lo que se hunde o flota realizando una migración vertical.
Walsh et al. (2006) señalaron que Trichodesmium es un fijador de nitrógeno tropical de origen caribeño. Nuestros datos sugieren que el agua tropical superficial del Caribe modificada, que se encuentra en la capa de mezcla, fue la fuente de Trichodesmium spp., incluyendo la comunidad de fitoplancton encontrada sobre la región del Cañón de Campeche. La circulación ciclónica y anticiclónica observada bajo la termoclina generó un frente térmico geostrófico subsuperficial, donde la conspicua presencia de tricomas de Trichodesmium fue registrada con las más altas concentraciones.
El fitoplancton asociado a Trichodesmium spp. estuvo dominado por especies oceánicas tropicales, principalmente dinoflagelados y particularmente especies de Ceratium. Taylor et al. (2008) señalaron que los dinoflagelados son un grupo protista dominante y que muestra su mayor diversidad en regiones oceánicas tropicales, particularmente miembros de cuernos largos como el género Ceratium. Las 16 especies de Ceratium identificadas en este estudio han sido previamente reportadas por Licea et al. (2004) para el sur del Golfo de México.
Se requiere profundizar en la investigación para explorar mejor el impacto de las abundancias de Trichodesmium en la productividad primaria y en los ciclos biogeoquímicos del nitrógeno en el sur del Golfo de México.
Agradecimientos
Este trabajo fue financiado por CONACYT (proyecto G27777–B), por la Universidad Nacional Autónoma de México (UNAM) y por la Universidad Autónoma Metropolitana–Xochimilco (UAM–X). Agradecemos al capitán y a la tripulación del B/O Justo Sierra su ayuda, y a J Castro por mejorar sustancialmente las figuras.
References
Antoine JW, Bryant WR. 1968. Major transition zones of the Gulf of Mexico: DeSoto and Campeche Canyons. AAPG Bull. 52. Doi: 10.1306/5D25C505–16C1–11D7–8645000102C1865D. [ Links ]
Capone DG, Zehr JP, Paerl HW, Bergman B, Carpenter EJ. 1997. Trichodesmium, a globally significant marine cyanobacterium. Science 276: 1221–1229. [ Links ]
Carpenter EJ. 1983. Nitrogen fixation by marine Oscillatoria (Trichodesmium) in the world's ocean. In: Carpenter EJ, Capone DG (eds.), Nitrogen in the Marine Environment. Academic Press, New York, pp. 65–103. [ Links ]
Carpenter EJ, Subramanian A, Capone DG. 2004. Biomass and primary productivity of the cyanobacterium Trichodesmium spp. in the tropical N Atlantic Ocean. Deep–Sea Res. I 51: 173–203. [ Links ]
Clark KR, Gorley RN. 2006. PRIMER v6: User Manual/Tutorial. PRIMER–E, Plymouth, 256 pp. [ Links ]
Creager JS. 1958. A canyon like feature in the Bay of Campeche. Deep–Sea Res. 5: 169–172. [ Links ]
Escobar–Briones E, Estrada–Santillán EL, Legendre P. 2008. Macrofaunal density and biomass in the Campeche Canyon, southwestern Gulf of Mexico. Tropical Studies in Oceanography. The Deep Gulf of Mexico Benthos Program. Deep–Sea Res. II 55(24–26): 2679–2685. [ Links ]
Fofonoff NP, Millard Jr RC. 1983. Algorithms for computation of fundamental properties of seawater. UNESCO Tech. Pap. Mar. Sci.. 44: 53 pp. [ Links ]
Franks PJS. 1992. Sink or swim: Accumulation of biomass at fronts. Mar. Ecol. Prog. Ser. 82: 1–12. [ Links ]
Furnas MJ, Smayda TJ. 1987. Inputs of subthermocline waters and nitrate onto the Campeche Bank. Cont. Shelf Res. 7: 161–175. [ Links ]
Grimshaw R, Broutman D, He X, Sun P. 1994. Analytical and numerical study of a barotropic eddy on a topographic slope. J. Phys. Oceanogr. 24: 1587–1607. [ Links ]
Gutiérrez–de Velasco G, Winant CD. 1996. Seasonal patterns of wind stress and wind stress curl over the Gulf of Mexico. J. Geophys. Res. 101(C8): 18127–18140. [ Links ]
Hernández–Becerril DU, García–Reséndiz JA, Salas–de León DA, Monreal–Gómez MA, Signoret–Poillon M, Aldeco–Ramírez J. 2008. Nanoplankton fraction in the phytoplankton structure in the southern Gulf of Mexico (April 2000). Cienc. Mar. 34(1): 77–90. [ Links ]
Hood RR, Coles VJ, Capone DG. 2004. Modeling the distribution of Trichodesmium and nitrogen fixation in the Atlantic Ocean. J. Geophys. Res. 109, C06006. Doi:10.1029/2002JC001753. [ Links ]
Hood RR, Laws EA, Armstrong RA, Bates NR, Brown CW, Carlson CA, Chai F, Doney SC, Falkowski PG, Feely RA, Friedrichs MAM, Landry MR, Moore JK, Nelson DM, Richardson TL, Salihoglu B, Schartau M, Toole DA, Wiggert JD. 2006. Pelagic functional group modeling: Progress, challenges and prospects. Deep–Sea Res. II 53: 459–512. Doi:10.1016/j.dsr2.2206.01.025. [ Links ]
Hsueh Y, O'Brien JJ. 1971. Steady coastal upwelling induced by an along–shore current. J. Phys. Oceanogr. 1: 180–186. [ Links ]
Karl DM, Letelier R, Hebel D, Tupas L, Dore J, Christian J, Winn C. 1995. Ecosystem changes in the North Pacific subtropical gyre attributed to the 1991/92 El Niño. Nature 373: 230–234. [ Links ]
Lenes JM, Darrow BP, Cattrall C, Heil CA, Callahan M, Vargo GA, Byrne RH, Prospero JM, Bates DE, Fanning KA, Walsh JJ. 2001. Iron fertilization and the Trichodesmium response on the West Florida Shelf. Limnol. Oceanogr. 46: 1261–1277. [ Links ]
Letelier RM, Karl DM. 1998. Trichodesmium spp. physiology and nutrient fluxes in the North Pacific subtropical gyre. Aquat. Microb. Ecol. 15: 265–276. [ Links ]
Licea S, Zamudio ME, Luna R, Soto J. 2004. Free–living dinoflagellates in the southern Gulf of Mexico: Report of data (1979–2002). Phycol. Res. 52: 419–428. [ Links ]
Mann KH, Lazier JR. 1991. Dynamics of Marine Ecosystems. Biological–Physical Interactions in the Oceans. Blackwell, Oxford, 466 pp. [ Links ]
Mojica–Ramírez E. 2003. Variabilidad de la biomasa zooplanctónica en la región del Cañón de Campeche durante la campaña PROMEBIO–I (5 al 21 de agosto 1999). B.Sc. dissertation, Facultad de Ciencias, Universidad Nacional Autónoma de México, 50 pp. [ Links ]
Nof D. 1999. Strange encounters of eddies with walls. J. Mar. Res. 57: 739–761. [ Links ]
Nowlin Jr WD. 1972. Winter circulation patterns and property distributions. In: Capurro LRA, Reid JL (eds.), Contributions on the Physical Oceanography of the Gulf of Mexico. Vol. II. Gulf Publishing Co., Houston, pp. 3–52. [ Links ]
Nowlin Jr WD, McLellan HJ. 1967. A characterization of the Gulf of Mexico waters in winter. J. Mar. Res. 25: 29–59. [ Links ]
Oey LY, Zhang HC. 2004. The generation of subsurface cyclones and currents through eddy–slope interaction. Cont. Shelf Res. 24: 2109–2131. [ Links ]
Ramírez–Cruz V. 2005. Distribución de la biomasa zooplanctónica en la región del Cañón de Campeche en fin de temporada de Nortes. B.Sc. dissertation, Facultad de Ciencias, Universidad Nacional Autónoma de Mexico, 81 pp. [ Links ]
Salas–de León DA, Monreal–Gómez MA, Signoret M, Aldeco J. 2004. Anticyclonic–cyclonic eddies and their impact on near–surface chlorophyll stocks and oxygen supersaturation over the Campeche Canyon, Gulf of Mexico. J. Geophys. Res. 109, C05012. Doi: 10.1029/2002JC001614. [ Links ]
Schroeder WW, Berner L, Nowlin WD. 1974. The oceanic waters of the Gulf of Mexico and Yucatan Strait during July 1969. Bull. Mar. Sci. 24: 1–19. [ Links ]
Shi C, Nof D. 1993. The splitting of eddies along boundaries. J. Mar. Res. 51:771–795. [ Links ]
Smith IV DC, O'Brien JJ. 1983. The interaction of a two–layer isolated mesoscale eddy with topography. J. Phys. Oceanogr. 13: 1681–1697. [ Links ]
Sournia A. 1968. La Cyanophycée Oscillatoria (= Trichodesmium) dans le plancton marin: Taxinomie et observations dans le canal de Mozambique. Nova Hedwigia 15(1): 1–12, pl. 1–2. [ Links ]
Steidinger KA, Tangen K. 1997. Dinoflagellates. In: Tomas CR (ed.), Identifying Marine Phytoplankton. Academic Press, pp. 387–584. [ Links ]
Subramaniam AC, Brown W, Hood RR, Carpenter EJ, Capone DG. 2002. Detecting Trichodesmium blooms in SeaWiFS imagery. Deep–Sea Res. II 49: 107–121. [ Links ]
Taylor FJR, Hoppenrath M, Saldarriaga JF. 2008. Dinoflagellate diversity and distribution. Biodiv. Conserv. 17: 407–418. [ Links ]
Tomas CR. 1997. Identifying Marine Phytoplankton. Academic Press, San Diego, 858 pp. [ Links ]
Tyrrell T, Marañon ER, Poulton AJ, Bowie AR, Harbour DS, Woodward M. 2003. Large–scale latitudinal distribution of Trichodesmium spp. in the Atlantic Ocean. J. Plankton Res. 25: 405–416. [ Links ]
Vázquez–de la Cerda AM, Reid RO, DiMarco SF, Jochens AE. 2005. Bay of Campeche circulation: An update. In: Sturges A, Lugo–Fernandez A (eds.), Circulation in the Gulf of Mexico: Observations and Models. Geophys. Monogr. Ser. 161. American Geophysical Union, Washington DC, pp. 279–294. [ Links ]
Vidal VMV, Vidal FV, Hernández AF, Meza E, Zambrano L. 1994. Winter water mass distribution in the western Gulf of Mexico affected by colliding anticyclonic ring. J. Oceanogr. 50: 559–588. [ Links ]
Villareal TA, Carpenter EJ. 1990. Diel buoyancy regulation in the marine diazotrophic cyanobacterium Trichodesmium thiebautii. Limnol. Oceanogr. 35: 1832–1837. [ Links ]
Walsh JJ, Jolliff JK, Darrow BP, Lenes JM, Milroy SP, Remsen A, Dieterle DA, Carder KL, Chen FR, Vargo GA, Weisberg RH, Fanning KA, Müller–Karger FE, Shinn E, Steidinger KA, Heil CA, Tomas CR, Prospero JS, Lee TN, Kirkpatrick GJ, Whitledge TE, Stockwell DA, Villareal TA, Jochens AE, Bontempi PS. 2006. Red tides in the Gulf of Mexico: Where, when, and why? J. Geophys. Res. 111, C11003. Doi:10.1029/2004JC002813. [ Links ]
Westberry TK, Siegel DA. 2006. Spatial and temporal distribution of Trichodesmium blooms in the world's oceans. Global Biogeochem. Cycles 20, GB4016. Doi: 10.1029/2005GB002673. [ Links ]
Westberry TK, Siegel DA, Subramanian A. 2005. An improved bio–optical model for the remote sensing of Trichodesmium blooms. J. Geophys. Res. 110, C06012. Doi:10.1029/2004JC002517. [ Links ]
Wood EJF. 1971. Phytoplankton distribution in the Caribbean Region. In: UNESCO (ed.), Coloquio sobre Investigaciones y Recursos del Mar Caribe y Regiones Adyacentes. CICAR, pp. 399–410. [ Links ]
* DESCARGAR VERSIÓN BILINGÜE (INGLÉS–ESPAÑOL) EN FORMATO PDF

