










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkCiencias marinas
Print version ISSN 0185-3880
Cienc. mar vol.35 n.3 Ensenada Sep. 2009
Artículos de investigación
Blue shrimp (Litopenaeus stylirostris) catch quotas as a management tool in the Upper Gulf of California*
La cuota de captura de camarón azul (Litopenaeus stylirostris) como instrumento de gestión en el Alto Golfo de California
AR García–Juárez1, 2*, G Rodríguez–Dominguez3, DB Lluch–Cota2
1 Instituto Nacional de Pesca, Centro Regional de Investigación Pesquera en Ensenada, Carretera Tijuana–Ensenada Km 97.5, El Sauzal de Rodríguez, CP 22760, Baja California, México.
2 Centro de Investigaciones Biológicas del Noroeste, Mar Bermejo No. 175, Col. Palo de Santa Rita, La Paz, CP 23090, Baja California Sur, México. * E–mail: argarcia@cibnor.mx
3 Facultad de Ciencias del Mar, Universidad Autónoma de Sinaloa, Paseo Claussen s/n, Col. Los Pinos, Mazatlán, Sinaloa, México.
Recibido en mayo de 2009.
Aceptado en agosto de 2009.
Abstract
Within the buffer zone of the Upper Gulf of California and Colorado River Delta Biosphere Reserve (established June 1993), the shrimp fishery is managed as a common and rational use resource, trying to minize the impact of fishing effort on both the resource and ecosystem. An analysis was thus made of potential catch quotas of Litopenaeus stylirostris using a dynamic model to estimate expected shrimp yield and future biomass that includes fishing season as a factor and catch per unit effort (CPUE) as abundance index. The CPUE of L. stylirostris declined in the 1990/91 season and tended to recover in 1995/96. The parameters of the biomass dynamic model were K = 21,323 t (±700 t, P < 0.05) and r = 1.18 (±0.1, P < 0.05). The estimated catchability was 0.00032 with a standard deviation of 0.25. Biomass was projected under three catch quota scenarios in the study area. Given the usefulness of establishing an annual catch limit for available biomass in the management of this resource, based on the scenarios analyzed, the adoption of quotas of 2200 and 2400 t for one fishing season is recommended, after which the effectiveness of this strategy should be assessed.
Key words: Upper Gulf of California, catch quota, Litopenaeus stylirostris.
Resumen
Dentro de la zona de amortiguamiento de la Reserva de la Biosfera del Alto Golfo de California y Delta del Río Colorado (establecida en junio de 1993), la pesquería de camarón se administra como un recurso de uso común y racional, tratando de que el esfuerzo pesquero impacte lo menos posible tanto al recurso como al ecosistema. Se analizaron posibles cuotas de captura estimando el rendimiento esperado y la biomasa futura de camarón mediante un modelo dinámico que usó la temporada de pesca como factor (denominado periodo), y la captura por unidad de esfuerzo (CPUE) como índice de abundancia. La CPUE de L. stylirostris declinó en la temporada 1990/91, tendiendo a la recuperación en 1995/96. Los parámetros del modelo dinámico de biomasa fueron K = 21,323 t (±700 t, P < 0.05) y r = 1.18 (±0.1, P < 0.05). La capturabilidad estimada fue de 0.00032 con una desviación estándar de 0.25. Se proyectó la biomasa con tres escenarios de cuota de captura dentro de la zona de estudio. Dada la utilidad de establecer un límite anual a la captura de biomasa disponible para la administración del recurso, y de acuerdo a los escenarios analizados, se propone la adopción de las cuotas de 2200 y 2400 t por una temporada de captura, y evaluar su efectividad al término del mismo.
Palabras clave: Alto Golfo de California, cuota de captura, Litopenaeus stylirostris.
Introducción
En el Pacífico mexicano se explotan cuatro especies de camarones peneidos: camarón azul Litopenaeus stylirostris (Stimpson 1874), camarón blanco L. vannamei (Boone 1931), camarón café Farfantepenaeus californiensis (Holmes 1900) y camarón cristalino o rojo F. brevirostris (Kingsley 1878); que aportan 70% de la producción nacional de camarón capturado. Sin embargo, por la distribución de estas especies, en el Alto Golfo de California (AGC) sólo se encuentran dos de ellas: camarón azul y camarón café (Rodríguez de la Cruz 1981).
La pesca industrial del camarón en el AGC se inició a principios de los años cincuenta. No obstante, las estadísticas históricas acerca de la pesca de este recurso datan a partir de 1956 (Avalos–Hernández 1974). La Reserva de la Biósfera del Alto Golfo de California y Delta del Río Colorado fue establecida por decreto presidencial el 10 de junio de 1993. Los objetivos explícitos de esta reserva son: (1) conservar los ecosistemas del desierto de Sonora, el AGC y el Delta del Río Colorado; (2) dar protección permanente a especies únicas como la totoaba, la vaquita marina, el pupo del desierto y una variedad de especies de aves; (3) regular las actividades productivas para la protección de recursos naturales; (4) promover actividades económicas alternativas que incrementen el nivel de vida de la población residente; (5) hacer investigación científica y educación ambiental en la región; y (6) recuperar y preservar la flora y fauna, así como la calidad del medio ambiente (Diario Oficial de la Federación, 10 de junio de 1993).
En la zona existen dos actividades que merecen especial atención: la explotación de los recursos y la conservación del ecosistema. El Instituto Nacional de Pesca tiene un programa de manejo para el recurso camarón en el Pacífico Mexicano, en el cual no se han integrado las cuotas de captura como una recomendación de administración a la pesquería, sin embargo, si considera objetivos ambientales, económicos y sociales. Los datos de captura por unidad de esfuerzo (CPUE) y la tendencia de la captura en el período 1976–1998 revelan dos periodos en la pesquería, de manera similar a lo encontrado para F. californiensis (Morales–Bojórquez y López–Martínez 1999 en Morales–Bojórquez et al. 2001): una disminución en las captura de 1976/77 a 1990/91 y un aumento de 1991/92 a 1997/98.
Al igual que lo realizado por Morales–Bojórquez et al. (2001), los datos de captura y esfuerzo fueron analizados con un modelo dinámico de biomasa, que es mejor que utilizar un modelo estructurados por edades como el análisis de población virtual (Pope y Sheperd 1985 en Morales–Bojórquez et al. 2001), sobre todo cuando la información sobre la estructura de edades de las capturas es pobre o no existe. Los modelos dinámicos de biomasa son las técnicas de evaluación del stock más usadas comúnmente en pesquerías tropicales, atún y peces con afinidad templada. Otra ventaja para considerar los modelos dinámicos de biomasa es que pueden utilizar cualquier índice de abundancia de la pesquería. Para tasas de captura, la capturabilidad es referida como el coeficiente q, mientras que para datos de crucero de investigación q es el sesgo relativo del crucero. En algunas circunstancias, los modelos dinámicos de biomasa pueden proveer estimaciones más exactas y precisas que las obtenidas con enfoques más complejos (Morales–Bojórquez et al. 2001).
En camarones peneidos se han realizado estudios para determinar si existe una relación parentela–progenie (stock reproductor–reculta: SRR) en sus poblaciones. En general se han encontrado dos tendencias: una SRR casi lineal o ninguna SRR. Sin embargo, se ha concluido que no existe evidencia que demuestre que el reclutamiento dependa del stock reproductor. Se considera que la sobrepesca del reclutamiento es poco factible, ya que se supone que la captura de reclutas o adultos se realiza cuando se encuentran en la fase plana de la curva teórica asintótica de SRR. Sin embargo, para el camarón blanco del Golfo de México se ha demostrado que existe una sobrepesca de reclutamiento (Gracia 1996). Si se considera que no existe SRR, entonces hay que capturar cuanto sea posible. Con esto en mente, se puede mantener un esfuerzo en constante aumento y sobrepasar un límite de equilibrio (sin stock reproductor no hay reclutamiento) causando la poco aceptada sobrepesca del reclutamiento.
El objetivo del presente estudio fue proyectar la biomasa del camarón azul L. stylirostris de la zona del AGC y estimar cuotas de captura bajo tres escenarios, utilizando como índice de abundancia la captura por unidad de esfuerzo.
Material y métodos
Los datos de captura anual (toneladas, peso vivo) y la CPUE (toneladas por barco) fueron obtenidos de los registros oficiales provenientes de la flota camaronera de arrastre de Puerto Peñasco y el Golfo de Santa Clara, Sonora y San Felipe, Baja California de 1987/88 a 2006/07 (tabla 1). Se analizó la información considerando los periodos anuales de la temporada de pesca, aproximadamente de septiembre a mayo (Rodríguez de la Cruz 1976 and Magallón–Barajas 1987 in Morales–Bojórquez et al. 2001) aunque en algunas temporadas se acortó este periodo. La captura total se separó por especies con la información de las plantas procesadoras de camarón en los puertos mencionados y para este estudio se utilizó sólo la captura de camarón azul en peso vivo.

Modelo
Al igual que Morales–Bojórquez et al. (2001), para describir la situación general de la pesquería de camarón se analizó la CPUE mediante el modelo dinámico de biomasa de Schaefer (Hilborn y Walters 1992) el cual relaciona la captura total y el esfuerzo de pesca. El ajuste de una curva parabólica de esta relación permite obtener una estimación del rendimiento máximo sostenible y el esfuerzo óptimo de la pesquería. Este modelo no supone condiciones de equilibrio, lo que representa una ventaja en la evaluación de organismos de ciclo de vida corto como el camarón azul. La expresión determinística del modelo es:

donde t = tiempo (temporada de pesca), Bt = biomasa de la población al tiempo t, Bt+1 = biomasa de la población al tiempo t + 1, r = tasa intrínseca del crecimiento de la población, K = tamaño de la biomasa virgen o capacidad de carga, Ct = captura al tiempo t.
Estimación de los errores de observación y proceso
Se supone que el indicador de abundancia relativa It está medido con error; en consecuencia, el índice es estimado como:

donde q es la capturabilidad, Bt es la biomasa de la población al tiempo t y vt es el error de observación suponiendo vt ≈ N(0, σv) e  = qBt (Punt 1992, Punt y Hilborn 1996). Este error de observación se estima mediante:
= qBt (Punt 1992, Punt y Hilborn 1996). Este error de observación se estima mediante:

Cuando se usó el error de observación se probó una hipótesis sobre la población, H: La variabilidad en la población fue causada por la incertidumbre acerca del verdadero valor del índice It.
La estimación de los parámetros rO, KO y δO, dado el índice It (el subíndice O se refiere al error de observación) fue realizada usando la siguiente función de verosimilitud (–1 nL):
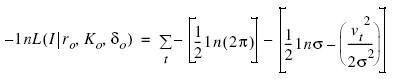
En esta expresión δ es una constante llamada deplete, que fue usada para estimar la biomasa durante la temporada de pesca con que inicia el análisis (Punt 1992) y σ es la desviación estándar del error de proceso, que fue estimada usando la siguiente solución analítica (Morales–Bojórquez et al. 2001), definida como:

En la serie de tiempo se considera que ha habido una evolución en los sistemas de pesca, así como en la experiencia de los pescadores, lo cual influye sobre el coeficiente de capturabilidad y del poder de pesca de la flota. Por consiguiente, es de esperar que el valor de capturabilidad cambie a través del tiempo. Para representar este patrón de cambio se hicieron las siguientes suposiciones: (a) el indicador de abundancia relativa tiene una distribución log normal; (b) el indicador de abundancia relativa estimado puede ser expresado como = qBtexp(vt), donde vt es el error de observación; (c) si el indicador de abundancia relativa It corresponde a cada año, entonces la capturabilidad para cada año puede ser expresada como qt = It/Bt, donde el subíndice t representa el tiempo; (d) el valor de capturabilidad de cada año bajo esta condición es variable, lo cual debe ser una condición obligada de las modificaciones en el esfuerzo que cambiaron la CPUE; y (e) el modelo no trabaja con los valores de capturabilidad de cada año, sino con un promedio dentro de toda la escala de tiempo en la cual se realizó el análisis, y por consiguiente, la expresión que representa el valor esperado de capturabilidad es estimado como:

Dado que se supuso una distribución log normal del indicador de abundancia relativa, entonces el promedio debe ser recalculado por su respectivo valor exponencial. La expresión muestra que la capturabilidad tiene una solución algebraica explícita que deriva de todos los valores de capturabilidad estimados para cada temporada de pesca, lo cual es un intento por valorar su variabilidad.
De los dos estimadores se calcularon dos importantes parámetros de manejo. el máximo rendimiento sostenido, MRS = rK/4, y la biomasa con la cual se puede alcanzar el MRS (BMRS), BMRS = K/2. La tasa óptima de explotación HR es HR = r/2 (Punt y Japp 1994).
Intervalos de confianza
Los intervalos de confianza de r y K fueron estimados para el modelo que mejor se ajustó a los datos (error de proceso o error de observación). Para estimar los intervalos de confianza se utilizó el perfil de verosimilitud (Venzon y Moolgavkor 1988 y Hilborn y Mangel 1997 en Morales–Bojórquez et al. 2001), ya que éste puede ser usado para determinar los intervalos de confianza de los parámetros conjunta o individualmente. Los intervalos de confianza para n parámetros fueron estimados basados en una distribución χ2 con m grados de libertad (Zar 1974 en Morales–Bojórquez et al. 2001). Para un sólo parámetro p, el intervalo de confianza (IC) es definido como todos los valores que satisfacen la siguiente condición (Polacheck et al. 1993 en Morales–Bojórquez et al. 2001).

donde L(Y/pest) es el logaritmo de la verosimilitud del mejor valor de p y χ1,1–a2 es el valor de la distribución chi cuadrada con un grado de libertad a un nivel de confianza 1 – α. Así, el intervalo de confianza al 95% para p abarca todos los valores de p, que es dos veces la diferencia entre el logaritmo de la verosimilitud y el logaritmo de la verosimilitud del mejor estimado de p, que es menor de 3.84 (Kimura 1981, Polacheck et al. 1993 and Morales–Bojórquez 1999 en Morales–Bojórquez et al.2001).
Resultados
Las mayores capturas se obtuvieron en la temporada 1988/ 89; después cayeron y permanecieron a la baja durante casi cuatro temporadas de pesca, y aunque se observó una recuperación de la captura, ya no se recobraron los niveles de 1987/88 and 1988/89 (tabla 1). En los dos últimos años del período estudiado las capturas oscilaron entre 2500 y 3000 t (tabla 1). Sin embargo, al analizar la tendencia de CPUE se observó que este índice se ha recuperado positivamente, lo cual permite suponer una recuperación en la abundancia si se considera que CPUE es proporcional a la abundancia. Los valores de la temporada 1999/2000 provocan que la información presente un aparente patrón de ciclos, el cual debería comprobarse posteriormente (tabla 1).
El ajuste del modelo dinámico de biomasa a los datos de CPUE muestra que es posible identificar dos periodos. el primero de 1988/89 a 1993/94 con un ajuste adecuado; y el segundo de 1994/95 a 2006/07, que no fue descrito de forma adecuada por el modelo (fig. 1) mostrando gran variabilidad de la CPUE. Los parámetros del modelo dinámico de biomasa fueron K = 21,323 t (con intervalo de confianza de ±700 t, P < 0.05) (fig. 2), y r = 1.18 (con intervalo de confianza de ±0.1, P < 0.05).


El modelo ajustado a los datos de CPUE arrojó un MRS de6325 t con una biomasa que permitiría obtenerlo (BMRS) de cerca de 10,662 t. De esta manera la tasa de explotación óptima corresponde a un valor del 0.59 de la biomasa disponible. De acuerdo con los datos históricos (tabla 1), es posible alcanzar el valor de MRS; sin embargo, considerando los datos de las temporadas más recientes es posible que la captura sea de cerca de 2500 t. En todo caso, se debe considerar que se resolvió un modelo que supone errores de medición en la CPUE.
Los valores estimados de cuota de captura mostraron que utilizar una cuota de 2220 t por temporada mantendría los niveles de biomasa por encima de las 6000 t, mientras que una cuota de 2400 t los mantendría cerca de las 6000 t (fig. 3).

Considerando estos dos escenarios de cuotas de captura la biomasa se mantendría por debajo del nivel de referencia del rendimiento estimado de 6325 t, aunque también se encontraría por debajo de su nivel de referencia (10,662 t). Se observó que estas cuotas de captura no permiten que la biomasa se incrementé a lo largo del tiempo, e incluso una cuota de captura de 2470 t provocaría una caída de la biomasa de la población (fig. 3). Las cuotas analizadas se basan en las capturas históricas registradas, pero el análisis hace evidente que existe un riesgo al mantener el nivel óptimo de la biomasa. Ante estos resultados la cuota de captura no debe exceder las 2400 t y este valor debiese disminuir para que exista un incremento en la biomasa de la especie, esto con el fin de utilizar una estrategia de cuota de captura para la pesquería del camarón azul en el AGC.
Discusión
Este trabajo presenta la primera estimación de biomasa para camarón azul en el AGC. Nuestros resultados muestran que la capacidad de carga del sistema para L. stylirostris es K = 21,323 t, mientras que la tasa intrínseca de crecimiento de la población es de r = 1.18. Con la estimación de estos parámetros además fue posible calcular que el máximo rendimiento sostenido puede ser de 6325 t. Por consiguiente, para obtener este rendimiento es necesario mantener una biomasa mínima de 10,662 t, la cual es entendida como la biomasa que nos llevaría a alcanzar el rendimiento esperado (BMRS). Con esta información pudimos calcular la tasa óptima de explotación, la cual se basa en el valor estimado de la tasa intrínseca de crecimiento de la población (0.59).
El nivel óptimo de rendimiento de 6325 t a partir de una BMRS de 10,662 t representa el primer punto de referencia biológico para la especie. Sin embargo, nosotros consideramos valores precautorios de captura sobre una base histórica de datos, de tal forma que nuestra propuesta muestra como se puede mantener el volumen de las capturas a largo plazo (fig. 3). Esto puede a su vez beneficiar el mantenimiento de la biomasa total del recurso. Los valores usados para la simulación fueron menores a la tasa óptima de explotación estimada, ya que ese valor sería el correspondiente a la extracción de 6325 t por temporada de pesca.
Debido al decreto de área natural protegida que ostenta la zona del AGC es imperativo proceder con un enfoque precautorio. La serie de datos de la pesquería de camarón muestra una gran variabilidad en la CPUE. Las capturas han disminuido a casi la mitad en las últimas 12 temporadas de pesca, lo cual implica que se deben utilizar criterios racionales para la explotación del recurso. El primer criterio es el establecimiento de una estrategia de manejo que límite las capturas, la cual podría consistir en una tasa de explotación constante (Hilborn y Walters 1992). En este trabajo, esta tasa de explotación constante se estimó en 0.59, lo que implicaría extraer 59% de la biomasa disponible cada año. De adoptar esta estrategia, se debe también determinar el tiempo que tomaría definir el éxito de esta medida administrativa, así como considerar procedimientos alternativos que permitan una estimación más precisa de la biomasa, principalmente mediante la investigación mediante cruceros y con arrastres experimentales dentro de la zona de estudio.
El modelo resuelto supone que la CPUE está medida con error, así que la estimación del MRS tiene implícita esa variación. La tendencia de la CPUE no evidenció un fuerte contraste en los datos y el ajuste de la serie presentó una variabilidad con patrones de cambio indefinidos, lo que permite plantear una hipótesis que relacione los cambios en el poder de pesca. Este problema es común en evaluación de stocks con modelos de biomasa dinámicos, ya que su principal suposición es que la capturabilidad permanece constante en el tiempo (McAllister y Kirkwood 1998 en Morales–Bojórquez et al. 2001). La primera suposición sobre los datos de CPUE puede no ser razonable por diversas causas. Por ejemplo, como se indicó antes, la capturabilidad puede incrementarse temporalmente conforme los pescadores conocen mejor el recurso y cómo explotarlo, lo cual puede ser también una función lineal que dependa del tamaño del stock (Atran y Loesch 1995 y Tanaka 1997 en Morales–Bojórquez et al. 2001). Se analizaron 20 temporadas de pesca y la variación en la capturabilidad puede ser la mayor fuente de error en la evaluación del stock que se basa en CPUE con capturabilidad constante (Ricker 1975, Hilborn y Walters 1992 y Ye y Mohammed 1999 en Morales–Bojórquez et al. 2001).
Las opiniones divergentes pueden estar parcialmente relacionadas con el tamaño de la serie de tiempo disponible, y con las posibles variaciones anuales de corto plazo (o ruido) que deben diferenciarse de las variaciones de largo plazo. Este hecho puede confundirse con variaciones relacionadas al esfuerzo de pesca, sesgando completamente la relación de la CPUE–esfuerzo y la evaluación resultante. Hay suficientes indicadores para mostrar que la producción de camarón varía de año en año con respecto a los factores climáticos de gran escala (Barrett y Ralph 1977, Lluch–Belda 1977, Da Silva 1986, Del Valle–Lucero 1989, Solana y Arreguín–Sánchez 1993 y Sheridan 1996 en Morales–Bojórquez et al. 2001). El modelo propuesto supone que el indicador de la pesquería (CPUE) tiene una distribución log normal y está medido con error. La influencia de la variabilidad ambiental y demás fenómenos climáticos es medida indirectamente cuando se utilizan estimadores con error de proceso, sin que necesariamente el error derivado tenga un significado explícito en términos ambientales (Morales–Bojórquez 1999, Morales–Bojórquez et al. 2001). En todo caso, el error de proceso considera que existen fuentes de variación ajenas a las mediciones del índice de la pesquería y de los parámetros estimados, que pueden ofrecer una mayor capacidad de ajuste del modelo a los datos. Utilizamos un modelo global en el que no se consideran los efectos del ambiente, pero éstos no se deben de perder de vista ya que en los hechos tienen una influencia importante sobre el recurso y sus variaciones de abundancia.
Los mecanismos son complejos e involucran varios parámetros como fecundidad, reclutamiento, crecimiento, supervivencia y capturabilidad (García y Le Reste 1981 en Morales–Bojórquez et al. 2001). Nuestro modelo considera una sola hipótesis para la estimación de parámetros, la hipótesis es el error de observación, la cual supone que la fuente del error se encuentra en los datos de la CPUE. Los efectos asociados de la variabilidad en la demografía de la especie (fecundidad, reclutamiento, crecimiento y supervivencia), así como el de la CPUE (capturabilidad) implican fuentes de variación que influyen en el error del índice de la pesquería. Por consiguiente, los estimados de rendimiento y tasa intrínseca de crecimiento de la población, así como de las cantidades de manejo derivadas de los parámetros del modelo, consideran un error de observación asociado únicamente a la CPUE.
Aparentemente, las variaciones en la biomasa y CPUE estimada con el estimador de error de proceso pueden sugerir una hipótesis biológica de efecto del ambiente en los cambios de la población y las tendencias de la CPUE. Específicamente, se consideró que eventos extraordinarios de escala anual, como El Niño, pueden causar cambios biológicos que podrían observarse en la captura (Valenzuela–Quiñones 1998, Lluch–Cota et al. 1999 y López–Martínez et al. 2000 en Morales–Bojórquez et al. 2001). Esta relación podría ser definida utilizando un análisis con variabilidad ambiental que explicara los principales procesos biológicos que pueden afectar el rendimiento de la pesquería de camarón (Morales–Bojórquez et al. 2001).
Sobre la base de la variabilidad previamente expuesta dentro de la zona de Reserva del AGC es factible una captura permisible de 2200 t, la cual de forma extraordinaria podría incrementarse hasta 2400 t. También sería de utilidad para la administración del recurso el establecimiento de una tasa de explotación constante del 0.59 de la biomasa disponible cada año, estrategia que se debería adoptar al menos por una temporada para evaluar su efectividad. Para precisar la biomasa disponible se recomienda realizar arrastres experimentales dentro de la zona de reserva del AGC con barcos de investigación del Instituto Nacional de Pesca.
Agradecimientos
Este trabajo fue realizado con financiamiento del Instituto Nacional de Pesca. El Consejo Nacional de Ciencia y Tecnología otorgó beca de posgrado al primer autor (CONACYT 190129).
Referencias
Avalos–Hernández M. 1974. Desarrollo histórico de las pesquerías de camarón (Penaeus sp.) y totoaba (Cynoscion macdonaldi) en la parte norte del Golfo de California. B.Sc. dissertation, Escuela Superior de Ciencias Marinas, Universidad Autónoma de Baja California, Ensenada, 37 pp. [ Links ]
Gracia A. 1996. White shrimp (Penaeus setiferus) recruitment overfishing. Mar. Freshwat. Res. 47. 59–65. [ Links ]
Hilborn R, Walters C. 1992. Quantitative Fisheries Stock Assessment. Choice, Dynamics and Uncertainty. Chapman and Hall, New York, 570 pp. [ Links ]
Hilborn R, Mangel M. 1997. The Ecological Detective. Confronting Models with Data. Monographs in Population Biology, Princeton Academic Press, 315 pp. [ Links ]
López–Martínez J, Lluch–Cota DB, Arreguín–Sánchez F, Hernández–Vázquez S. 2000. Population dynamics of brown shrimp (Farfantepenaeus californiensis) in the Gulf of California as related to El Niño events. In. Wooster WS, LeBlond PH, Allen RL, Leaman BM, Low LL, Tillman MF (eds.), Beyond El Niño. A Conference on Pacific Climate Variability and Marine Ecosystems Impacts, from the Tropics to the Arctic. La Jolla, California, 23–26 March, 98 pp. [ Links ]
Morales–Bojórquez E. 1999. Análisis del error de proceso y de observación en la estimación del reclutamiento de la sardina del Pacífico, Sardinops sagax caeruleus (Girard), en la costa suroeste de la Península de Baja California, México. Cienc. Mar. 25(4). 597–608. [ Links ]
Morales–Bojórquez E, López–Martínez J. 1999. The brown shrimp fishery in the Gulf of California, Mexico. CalCOFI Rep. 40. 28. [ Links ]
Morales–Bojórquez E, López–Martínez J, Hernández–Vázquez S. 2001. Dynamic catch–effort model for the brown shrimp Farfantepenaeus californiensis (Holmes) from the Gulf of California, Mexico. Cienc. Mar. 27(1). 105–124. [ Links ]
Polacheck T, Hilborn R, Punt A. 1993. Fitting surplus production models. Comparing methods and measuring uncertainty. Can. J. Fish. Aquat. Sci. 50. 2597–2607. [ Links ]
Punt A. 1992. Selecting management methodologies for marine resources, with an illustration for southern African hake. S. Afr. J. Mar. Sci. 12. 943–958. [ Links ]
Punt A, Hilborn R. 1996. Biomass dynamic models. User's manual. FAO Comput. Inf. Ser. (Fish.) No. 10. 62 pp. [ Links ]
Punt A, Japp DW. 1994. Stock assessment of the Kingklip (Genypterus capensis) resource off South Africa. S. Afr. J. Mar. Sci. 14. 133–149. [ Links ]
Rodríguez de la Cruz MC. 1981. Aspectos pesqueros del camarón de altamar en el Pacífico mexicano. Cienc. Pesq. 1(2). 1–19. [ Links ]
Venzon DJ, Moolgavkor SH. 1988. A method for computing profile likelihood–based confidence intervals. Appl. Stat. 37. 87–94. [ Links ]
* DESCARGAR VERSIÓN BILINGÜE (INGLÉS–ESPAÑOL) EN FORMATO PDF

