










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkCiencias marinas
versão impressa ISSN 0185-3880
Cienc. mar vol.34 no.3 Ensenada Set. 2008
Artículos
Dinámica del carbono y nitrógeno orgánico disueltos en el estuario del Duero, Portugal
Dissolved organic carbon and nitrogen dynamics in the Douro River estuary, Portugal
C Magalhães1*, C Teixeira1,2, R Teixeira1, A Machado1, I Azevedo1,2, AA Bordalo1,2
1 Centre of Marine and Environmental Research (CIIMAR), University of Porto, Porto, Portugal. *E-mail: c_magalhaes@yahoo.com, cmag@icbas.up.pt
2 Laboratory of Hydrobiology, Institute of Biomedical Sciences (ICBAS-UP), University of Porto, Porto, Portugal.
Recibido en febrero de 2008.
Aceptado en agosto de 2008.
Resumen
El carbono orgánico disuelto (COD) y el nitrógeno orgánico disuelto (NOD) son compuestos metabólicamente importantes en los sistemas estuarinos. Su disponibilidad es influenciada por aportes de materia orgánica (internos y externos) y por transformaciones microbianas internas. En este trabajo se evaluaron las dinámicas espaciales y temporales del COD y NOD en el estuario del Río Duero. Los muestreos se realizaron mensualmente entre octubre de 2005 y diciembre de 2006 a lo largo del gradiente salino del estuario, a tres profundidades distintas. El COD y NOD fueron calculados mediante la substracción de los respectivos componentes inorgánicos del CDT (carbono disuelto total) y el NDT (nitrógeno disuelto total). El carbono inorgánico disuelto (CID) en el estuario del Duero mantiene una relación lineal positiva con la salinidad, indicando un aporte de CID al estuario por parte de las aguas costeras. Este patrón de distribución a lo largo del gradiente salino fue también observado para el material particulado total, ya que se observó una tendencia decreciente corriente arriba. En contraste, los resultados mostraron un comportamiento no conservativo del COD durante gran parte de los meses, en su mayoría asociado al aporte de COD de origen antropogénico en las porciones media y inferior del estuario. Las concentraciones de NOD representan 52-88% del NDT. A lo largo del transecto las relaciones entre nitrógeno inorgánico disuelto (NID) y NOD con la salinidad mostraron un comportamiento conservativo, lo que indica una fuerte influencia del Río Duero en las concentraciones de NID y NOD del estuario. No obstante, se han registrado aportes ocasionales de NOD de origen antropogénico. Las exportaciones globales de COD y NOD de la cuenca hidrográfica del Duero a las aguas costeras fueron estimadas en 420 kg C m-2 año-1 y 125 kg N m-2 año-1, respectivamente, las cuales son elevadas al compararse con los resultados de modelos de exportacion global de COD y NOD para la misma cuenca.
Palabras clave: carbono orgánico disuelto, estuario del Duero, nitrógenio orgánico disuelto.
Abstract
Dissolved organic carbon (DOC) and dissolved organic nitrogen (DON) are metabolically important in estuaries. Their availability is influenced by organic matter inputs (internal and external) as well as by internal microbial transformations within the system. In this study, spatial and seasonal dynamics of DOC and DON were evaluated in the Douro River estuary (Portugal). Monthly surveys were performed from October 2005 to December 2006 along a salinity gradient at three different depths. DOC and DON were calculated by subtracting the respective inorganic components (dissolved inorganic carbon, DIC, and dissolved inorganic nitrogen, DIN) from total dissolved carbon (TDC) and total dissolved nitrogen (TDN). In the Douro River estuary, DIC exhibits a linear and positive distribution as a function of salinity, indicating an input of DIC into the estuary from coastal waters. This pattern of distribution along the salinity gradient was also observed for total particulate matter, since a decreasing trend upstream was observed. In contrast, the results showed a general nonconservative behaviour of DOC along the estuarine transect during most months. This typical deviation from conservative mixing was mainly associated with sewage DOC sources into the lower and middle estuary. On average, DON concentrations represented 52-88% of the TDN. Along the transect, relationships between DIN and DON with salinity were generally conservative, decreasing towards the mouth, highlighting the fluvial source of such nitrogen forms; however, occasional DON sewage loads were registered. Estimated global Douro River watershed exports of DOC and DON to coastal waters yielded values of 420 kg C m-2 yr-1 and 125 kg N m-2 yr-1, respectively, which are high when compared with previously predicted levels from global models of DOC and DON export.
Key words: dissolved organic nitrogen, dissolved organic carbon, Douro River estuary.
Introducción
La materia orgánica disuelta (MOD) es un componente principal de la materia orgánica transportada a la zona costera por los ríos (Meybeck 1982, Ludwig et al. 1996); influye en las tramas alimenticias, controla la disponibilidad de nutrientes y metales disueltos, y afecta las propiedades ópticas de los sistemas acuáticos (Findlay y Sinsabaugh 2003). Además de controlar procesos a nivel ecosistema, la MOD también es importante desde una perspectiva biogeoquímca regional y global, ya que constituye una importante ruta para el transporte de carbono (C) y nitrógeno (N) de la tierra al mar. Estos componentes son metabólicamente importantes en los estuarios y su disponibilidad está influenciada por los aportes de materia orgánica (internos y externos) y las transformaciones microbianas que allí ocurren. Los principales mecanismos de producción de MOD in situ incluyen su exudado por el fitoplancton, su excreción por organismos proto y metazoarios pastoreadores, y la degradación viral de las células de fitoplancton y bacterias (Nagata 2000). La potencial utilización o remineralización de los sustratos de origen orgánico está determinada primordialmente por la razón C:N (Goldman et al. 1987). Otro factor importante que afecta la dinámica de la MOD en los sistemas estuarinos es la presión antropogénica mediante, por ejemplo, la construcción de presas que modifican las cantidades de agua dulce, sedimento y materia orgánica e inorgánica, que llegan a los estuarios y sus zonas costeras adyacentes, así como el momento en que lo hacen (Hopkinson y Vallino 1995). La descarga de aguas negras, tratadas o sin tratar, a los estuarios también afecta su metabolismo al incrementar los nutrientes alóctonos, o bien, los aportes de materia orgánica, respectivamente. Además, en varios ecosistemas estuarinos se ha evidenciado la importancia de la hidrología para retener estos elementos (Nixon et al. 1996).
Para comprender mejor estos procesos se requieren datos sobre nitrógeno orgánico disuelto (NOD) y carbono orgánico disuelto (COD), los cuales no han sido registrados previa ni rutinariamente en el estuario del Río Duero. Este río drena la principal cuenca hidrológica de la Península Ibérica, cuyos 98,000 km2 son compartidos en diferente proporción entre Portugal (20%) y España (80%). Más de 50 grandes presas han sido construidas para irrigación y generación de energía eléctrica sobre su cauce, principalmente en los últimos 50 años, dando como resultado la regulación de su caudal. El estuario del Duero se encuentra limitado hacia el río por la última de estas presas, ubicada a 21.6 km de la boca. Ésta determina el flujo de agua dulce hacia el estuario (Vieira y Bordalo 2000) y representa la mayor fuente de biomasa fitoplanctónica para el mismo (Bordalo y Vieira 2005). El flujo del río presenta una gran variabilidad interanual, con diferencias considerables entre años lluviosos y secos. Se ha reportado una disminución en el flujo anual debida al cambio climático global, pero también debida al mayor almacenamiento de agua para la generación de energía hidroeléctrica, la agricultura y el consumo doméstico (Bordalo y Vieira 2005). El estuario del Duero se encuentra en una de las zonas más pobladas de Portugal, en cuyas inmediaciones se encuentran tres importantes ciudades (Porto, Vila-Nova-de-Gaia y Gondomar), con un total de 700 000 habitantes que ejercen una importante presión antropogénica sobre el estuario en términos de drenaje urbano y aguas negras, terrenos ganados al estuario y construcciones costeras. En total ocho plantas de tratamiento de aguas negras descargan al estuario. En este trabajo se investigaron las fuentes y la distribución del COD y el NOD a lo largo del gradiente salino del estuario del Duero.
Métodos
Area de estudio y muéstreos
El estuario del Duero tiene un régimen mesomareal semidiurno, con una profundidad media de 8.2 m y una amplitud mareal media de 2.8 m. El tiempo de residencia del agua puede llegar hasta 14 días durante las mareas muertas de verano, mientras que durante eventos de altas descargas, el estuario se vacía completamente en un solo ciclo de mareas evitando la intrusión del agua de mar durante el flujo (Vieira y Bordalo 2000). El estuario se encuentra verticalmente estratificado aún en condiciones de bajo flujo del río (Vieira y Bordalo 2000). Las temperaturas de la columna de agua fluctuan entre 7°C en invierno y 22°C en verano, y la salinidad en la parte baja del estuario varía entre 0 y 35.
Los datos presentados en este estudio se obtuvieron en el marco de un programa más amplio de muestreos realizado por contrato con Servidos Municipalizados de Águas e Saneamiento do Porto, diseñado para evaluar el nivel de contaminación fecal de las aguas del estuario. Entre octubre de 2005 y diciembre de 2006 se visitaron mensualmente ocho estaciones a lo largo del gradiente salino del estuario, durante la bajamar, cubriendo descargas fluviales contrastantes (fig. 1). Se obtuvieron perfiles verticales de temperatura, conductividad, salinidad, oxígeno disuelto, pH y turbidez, mediante un CTD YSI 6600. Simultáneamente se recolectaron muestras de agua a tres profundidades (superficie, media agua y fondo) con una botella Van Dorn para evaluar nutrientes (nitrato, nitrito, amonio), materia particulada total (MPT), materia orgánica particulada (MOP), carbono disuelto total (CDT), nitrógeno disuelto total (NDT), y bacterias fecales. Estas muestras fueron almacenadas en frascos lavados con ácido, refrigeradas en hieleras y transportadas en oscuridad al laboratorio para ser procesadas menos de una hora después de su recolección.

Análisis de coliformes fecales
La determinación de coliformes fecales en las muestras de agua estuarina se realizó de acuerdo con el estándar ISO 9308/ 1 (ISO 1990). Las bacterias se concentraron en filtros estériles de membrana de 0.45 µm, los cuales fueron colocados en cajas de Petri con medio de cultivo mFC (Merck 10426) e incubados a 44.5°C. Después de incubarlas por 24 h se contaron las colonias de coliformes fecales y los datos se expresaron como unidades formadoras de colonias (UFC) por 100 mL de muestra de agua estuarina.
Procedimientos analíticos
Para la evaluación de la MPT y la MOP se prefiltraron las muestras de agua del estuario a través de filtros de fibra de vidrio GF/F pre-combustionados (Whatman), y filtros secados a 105°C y luego incinerados a 500°C (APHA 1992). Las determinaciones de CDT, carbono inorgánico disuelto (CID) y NDT se realizaron mediante un analizador Shimadzu Instruments TOC-VCSN acoplado a una unidad medidora de nitrógeno total TNM-1, Shimadzu. El CDT se midió por oxidación catalítica a alta temperatura seguida por la detección infrarroja no dispersiva del CO2. Para determinar el CID se acidificaron de manera automática las muestras con HCL 2M al 1.5% y se rociaron con gas transportador (aire purificado) para sólo convertir el carbono inorgánico a CO2. El COD se determinó mediante la diferencia entre CDT y CID. El NDT se descompuso térmicamente en un tubo de combustión, y el óxido nítrico resultante fue detectado mediante quimioluminiscencia. Se analizaron de tres a cinco replicas de CDT, CID y NDT, logrando varianzas del coeficiente menores a 2%. Al principio de cada corrida analítica para CDT y CID se generaron curvas estándar de ftalato ácido de potasio (C8H5KO4) de alto grado y carbonato ácido de sodio (NaHCO3) más carbonato de sodio (Na2CO3), respectivamente. Para el NDT se generaron curvas adicionales de nitrato de potasio (KNO3) de alto grado. Los límites de detección para CDT, CID y NDT fueron, respectivamente, 50 µg L-1, 4 µg L-1 and 0.1 mg L-1. El NOD se calculó como la diferencia entre NDT y nitrógeno inorgánico disuelto (NID: NH4 + NOX). Nitrito y amonio fueron analizados siguiendo los métodos descritos por Grasshoff et al. (1983), mientras que el nitrato se cuantificó mediante una adaptación de la técnica de reducción por cadmio esponjoso (Jones 1984), substrayendo el valor del nitrito del total. Todos los análisis del nitrógeno inorgánico se realizaron por triplicado y el coeficiente de variación estuvo en el rango de 0.1% a 8% dependiendo de la concentración del nutriente en particular.
Análisis de los datos
El análisis estadístico se realizó a un nivel de confianza del 95% (P < 0.05). Se probó la normalidad de los datos usando la prueba de Kolmogorov-Smirnov y su homocedasticidad con la de Leven (Zar 1996). Se realizó un análisis de regresión lineal (Zar 1996) para establecer relaciones del COD y el CID con el nitrógeno (NOD y NID) y otros parámetros ambientales medidos.
Resultados
La descarga media mensual del río durante el periodo de estudio (octubre de 2005 a diciembre de 2006) varió entre 39 y 1450 m3 s-1, con un promedio de 432 m3 s-1. Las descargas mínimas ocurrieron durante el verano y a principios del otoño (junio-octubre), mientras que otoño, invierno y primavera se caracterizaron por altas descargas (noviembre-mayo), con una descarga máxima registrada el 26 de noviembre de 2006 durante el flujo de mareas, cuando la descarga horaria del río alcanzó los 7500 m3 s-1 (fig. 2). La estructura longitudinal de la salinidad correspondiente a un influjo bajo, intermedio y alto del río durante el reflujo de marea se presenta en la figura 3a-c.

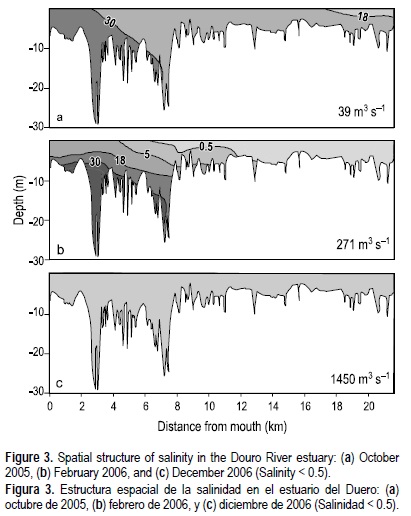
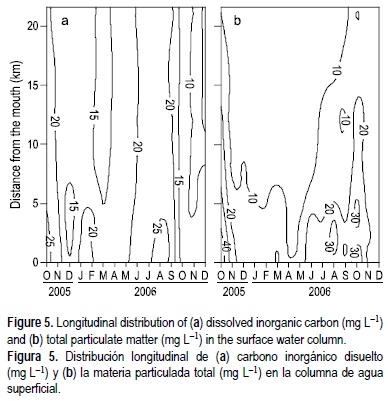
En la tabla 1 se presentan los promedios, intervalos y errores estándar de algunas variables selectas obtenidas durante este estudio. La concentración del CDT varió de 12.9 a 28.2 mg L-1 y estuvo compuesta por una fracción mayor de CID (8.4-25.5 mg L-1) que la de COD (1.7-4.5 mg L-1). El CID en el estuario del Duero mostró un comportamiento muy conservativo a lo largo del gradiente salino (figs. 4, 5a). Este mismo patrón de distribución también se observó para la MPT y sus fracciones orgánica (MOP) e inorgánica (fig. 5b); es decir, se observó una tendencia a incrementar hacia la boca del estuario, corriente abajo. Durante los meses de flujo bajo todo el estuario estuvo dominado por agua de mar y salobre, y se encontraron concentraciones elevadas de CID y MPT, mientras que en momentos de flujo alto del río se observaron valores menores de estas variables (fig. 5).

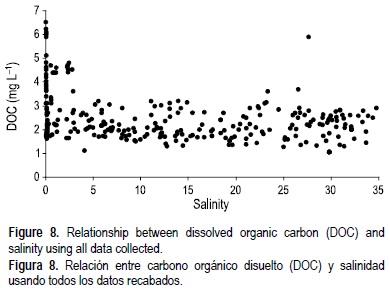
En este estudio las mayores concentraciones de COD (noviembre y diciembre de 2006; fig. 6) corresponden a situaciones de alto influjo del río y flujo de mareas (fig. 2), lo que causó que las concentraciones de COD se incrementaran abruptamente dentro del estuario. La curva de mezcla generada con todos los datos de COD muestra un comportamiento general no conservativo (figs. 7, 8). Las curvas de mezcla independientes para cada mes de COD también mostraron típicamente desviaciones de la mezcla conservativa con excepción de las de enero, mayo, junio y julio de 2006. En esos meses se observó un gradiente del COD similar al de la salinidad, siendo el extremo del agua dulce la mayor fuente de COD para el estuario (fig. 9a). En el COD se observó un comportamiento no conservativo en situaciones de alto influjo fluvial (noviembre y diciembre de 2006) en las que toda la columna de agua estaba bien mezclada, el agua dulce dominaba todo el estuario, y las concentraciones de COD no variaban a lo largo del estuario (fig. 9c). En condiciones de influjo fluvial bajo e intermedio el comportamiento no conservativo del COD se veía relacionado con los aportes de COD de los vertidos de drenaje en la parte baja y media del estuario (fig. 9b).

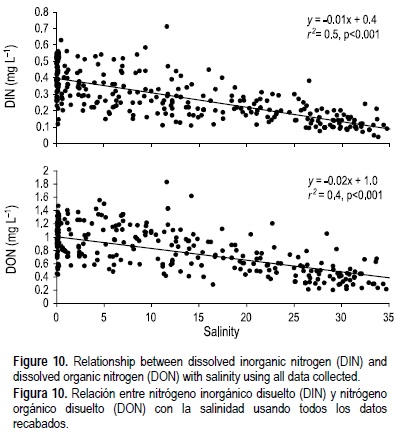
Se observó una clara estacionalidad en las concentraciones de NID y NOD en el estuario, y ambas formas de nitrógeno mostraron el mismo patrón de variación (fig. 6c, d). Las concentraciones medias máximas de NID y NOD ocurrieron cuando el caudal del río era mayor (tabla 1, fig. 2), y cuando el agua dulce del Duero ocupaba casi todo el estuario (fig. 3c), mientras que los valores de estas variables fueron menores al reducirse las descargas del río (tabla 1, fig. 2), coincidiendo con una cuña de agua salada que penetraba en el estuario (fig. 3a). El aporte de N al estuario del Duero estuvo dominado por la fracción orgánica ya que las concentraciones medias de NOD (0.4-1.2 mg L-1) siempre fueron mayores que las de NID (0.2-0.5 mg L-1). A lo largo del transecto estuarino la relación entre NID y NOD, y la salinidad, fue generalmente conservativa (fig. 10). Sin embargo, en abril, mayo, y agosto de 2006 se observó un comportamiento no conservativo relacionado con las cargas de NOD y NID del drenaje de aguas negras, debido a que tales compuestos aumentaban con el incremento de las coliformes fecales en las partes media y baja del estuario (en la figura 11 se muestra un ejemplo para abril de 2006).

Discusión
Hidrología
El principal forzamiento para la circulación del agua en el estuario del Duero fue el influjo de agua dulce. La descarga media mensual del Río Duero en el largo plazo (1985-2005) es 455 m3 s-1 (Bordalo y Vieira 2005), no obstante el flujo puede aumentar de 0 a >900 m3 s-1 en cuestión de minutos, según el régimen de descargas de la presa río arriba. Por ello, el promedio mensual de descarga fluvial del periodo de estudio representa un año regular en lo que concierne al flujo del río.
En el estuario del Duero la cuña de agua salada se ubica en función del estado de la marea y la descarga de agua dulce controlada por la presa. Aunque bajo un alto influjo fluvial toda la columna de agua se encontró bien mezclada y dominada por el agua dulce, con descargas por debajo del umbral de 800 m3 s-1 se presenta estratificación vertical (Vieira y Bordalo 2000). En este último caso los contornos de salinidad mostraron una cuña de agua salada penetrando durante el flujo de marea, misma que se conservaba dentro de los límites del estero durante el reflujo.
Dinámica del carbono y nitrógeno disueltos
No existe información acerca del COD y el NOD en el estuario del Duero. En la literatura sólo se encuentran disponibles valores de COD basados en un solo crucero (Abril et al. 2002), por lo que este estudio constituye la primera evaluación comprehensiva de la biogeoquímica de estos grupos de compuestos en el sistema. Aquí se presentan mediciones mensuales hechas a lo largo del gradiente salino entre la boca del río y el extremo superior del estuario, esto a fin de entender la dinámica estuarina de los componentes claves de la materia orgánica disuelta.
El CID en el estuario del Duero mostró una relación monotónica positiva con la salinidad, lo que indica un aporte de CID al estuario desde las aguas costeras adyacentes. Este patrón de distribución a lo largo del gradiente salino también se observó para la MPT, cuya dinámica en los estuarios depende del transporte de sedimentos de los ríos y de sedimentos suspendidos a la deriva localmente, asociados a un máximo de turbidez o a la resuspensión de sedimentos del lecho marino (Uncles et al. 2000). La dinámica de la MPT en el estuario del Duero puede ser explicada por el alto nivel de retención de sedimentos por la red de más de 50 grandes presas río arriba que retienen la materia suspendida del agua descargada hacia el estuario, pero también por el vertido de aguas negras en las partes media y baja del estuario y por las mayores tasas de resuspensión de sedimentos causadas por las mayores velocidades de la corriente en la parte baja del mismo (Azevedo et al. 2006, Azevedo et al. 2008). No obstante, las cargas de MPT al estuario del Duero son relativamente bajas en comparación con otros sistemas estuarinos europeos tales como los estuarios del Sado, la Gironda y Scheldt (Cabecadas et al. 1999).
En este estudio las concentraciones de COD se encontraron en un rango menor en comparación con otros sistemas estuarinos (c.f. Abril et al. 2002, Goni et al. 2003, Harrison et al. 2005). Dado que el COD es un componente mayor de la materia orgánica fluvial importada hacia los estuarios (Spitzy e Ittekkot 1991) usualmente disminuye linealmente conforme se incrementa la salinidad. Sin embargo, esto no necesariamente implica un comportamiento conservativo ya que esto podría deberse a la existencia de fuentes y sumideros simultáneos que resultarían en cambios netos casi nulos (Moran et al. 1999, Raymond y Bauer 2000, Abril et al. 2002, Goni et al. 2003). El comportamiento general no conservativo del COD observado en el estuario del Duero sugiere la existencia de fuentes que podrían variar temporal y espacialmente dependiendo de los aportes externos (descarga fluvial, pulsos de descarga de aguas negras) y o internos. Los resultados mostraron aportes de COD de drenaje en las partes baja y media del estuario que coincidieron en buena medida con los conteos de coliformes fecales (indicadoras de descargas de aguas negras) y con las concentraciones de COD observadas (fig. 9c). La contaminación inducida por el hombre en las partes baja y media del estuario del Duero ya había sido descrita por Bordalo (2003), y previamente se había encontrado que incrementaba la respiración del sistema contribuyendo a la heterotrofia observada todo el año en el metabolismo pelágico de las porciones baja y media del estuario del Duero (Azevedo et al. 2006).
La mayoría de los estudios en sistemas estuarinos están enfocados en la bioquímica del nitrógeno inorgánico (NID, nitrato + nitrito + amonio), mientras que los datos sobre NOD son más bien escasos. Sin embargo, éste representa una reserva importante de N reducido, ya que una porción considerable de los aportes de N a los ecosistemas costeros es en forma orgánica (e.g. Meybeck 1982, Seitzinger y Sanders 1997, Lewis et al. 1999, Seitzinger et al. 2002). En este estudio tanto la forma inorgánica como la orgánica mostraron el mismo patrón de variación, lo que denota una fuente y un comportamiento común. Se observó una influencia fluvial preponderante en las concentraciones de NID y NOD en las aguas del estuario del Duero. Los aportes de N al estuario del Duero estuvieron dominados por la fracción orgánica; en promedio, las concentraciones de NOD representaron de 52% a 88% del NDT. Mientras que éste no es siempre el caso (Mortazavi et al. 2000), el NOD es frecuentemente más abundante que el NID en los sistemas estuarinos (Eyre y Pont 2003), particularmente en sistemas que reciben aguas negras no tratadas, debido al tiempo necesario para la transformación biológica de las formas orgánicas. Las cargas antropogénicas de NOD y NID al estuario del Duero no fueron tan evidentes como las observadas para el COD, ya que el comportamiento conservativo de estos compuestos dominó en la mayoría de los muestreos. Sin embargo, en abril, mayo y agosto de 2006 se registraron cargas de NOD y NID provenientes de aguas negras (fig. 11). Dado que el N es el recurso limitante de nutrientes primordial para la producción algal y microbiana en muchos ambientes marinos y algunos de agua dulce, los aumentos en los aportes de NOD pueden alterar notablemente tales ecosistemas (e.g., Ryther y Dunstan 1971, Vitousek y Howarth 1991). Por ello resulta de considerable interés investigar los efectos de la contaminación inducida por el hombre en forma de sobrecargas de NOD a la producción, biomasa y composición específica del fitoplancton, particularmente en el estuario del Duero donde la presión antropogénica es evidente.
Balance anual de carbono y nitrógeno disuelto
La magnitud y estacionalidad del aporte de agua dulce al estuario del Duero se encuentra determinada principalmente por el patrón de flujo del Río Duero, el cual es manejado más de acuerdo con las necesidades de producción hidroeléctrica que con criterios ambientales (Azevedo et al. 2008). En este estudio se estimaron el NID y NOD exportados por el estuario del Duero a partir de sus concentraciones medias en la columna de agua y las descargas fluviales. Los cálculos integrados anualmente resultaron en una exportación hacia la zona costera de 1.2 × 107 kg año-1 de NOD, lo que corresponde a 125 kg año-1 de NOD exportado por kilómetro cuadrado de la cuenca del Duero (98,000 km2). Estos valores difieren de los predichos para el NOD exportado hacia aguas costeras por cuencas individuales alrededor del mundo calculados por el modelo global desarrollado por Harrison et al. (2005) para el NOD, el cual arrojó valores menores para la cuenca hidrológica del Río Duero (30-50 kg N km-2 año-1). En el caso del COD, aunque el río representa su fuente principal, este estudio reveló que existen fuentes antropogénicas adicionales en las partes media y baja del estuario. Al considerar la cuenca hidrológica del Duero, la exportación anual de COD hacia la zona costera se estimó en 4.2 × 107 kg año-1 ó 431 kg C km-2 año-1. En contraste con lo que sucede con el NOD, la exportación de COD por el estuario del Duero calculada en este estudio resultó similar al mayor intervalo de valores predichos por los modelos globales de exportación de COD de los ríos (Harrison et al. 2005), cuyas predicciones para el área geográfica del Duero fueron de 170-340 kg C km-2 año-1.
Agradecimientos
Agradecemos a L Lima su asistencia en el laboratorio. Este trabajo fue parcialmente financiado por Servicos Municipalizados de Águas e Saneamiento do Porto (SMAS) mediante un contrato asignado a AA Bordalo.
Traducido al español por Manuel Gardea-Ojeda.
Referencias
APHA, WWA, WEF. 1992. Standard Methods for the Examination of Water and Wastewater. 18th ed. American Public Health Association, Washington, DC, pp. 256-257. [ Links ]
Abril G, Nogueira M, Etcheber H, Cabeçadas G, Lemaire E, Brogueira MJ. 2002. Behaviour of organic carbon in nine contrasting European estuaries. Estuar. Coast. Shelf Sci. 54: 241-262. [ Links ]
Azevedo IC, Duarte PM, Bordalo AA. 2006. Pelagic metabolism of the Douro estuary (Portugal): Factors controlling primary production. Estuar. Coast. Shelf Sci. 69: 133-146. [ Links ]
Azevedo IC, Duarte PM, Bordalo AA. 2008. Understanding spatial and temporal dynamics of key environmental characteristics in a mesotidal Atlantic estuary (Douro, NW Portugal). Estuar. Coast. Shelf Sci. 76: 620-633. [ Links ]
Bordalo A. 2003. Microbiological water quality in urban coastal beaches: The influence of water dynamics and optimization of the sampling strategy. Water Res. 37: 3233-3241. [ Links ]
Bordalo A, Vieira M. 2005. Spatial variability of phytoplankton, bacteria and viruses in the mesotidal salt wedge Douro Estuary (Portugal). Estuar. Coast. Shelf Sci. 63: 143-154. [ Links ]
Cabeçadas G, Nogueira M, Brogueira MJ. 1999. Nutrient dynamics and productivity in three European estuaries. Mar. Pollut. Bull. 38: 1092-1096. [ Links ]
Eyre BD, Pont D. 2003. Intra- and inter-annual variability in the different forms of diffuse nitrogen and phosphorus delivered to seven sub-tropical east Australian estuaries. Estuar. Coast. Shelf Sci. 57: 137-148. [ Links ]
Findlay SEG, Sinsabaugh RL. 2003. Aquatic Ecosystems: Interactivity of Dissolved Organic Matter. Elsevier, New York, 512 pp. [ Links ]
Goldman JC, Caron DA, Dennett MR. 1987. Regulation of gross growth efficiency and ammonium regeneration in bacteria by substrate C:N ratio. Limnol. Oceanogr. 32: 1239-1252. [ Links ]
Goni MA, Teixeira MJ, Perkey DW. 2003. Sources and distribution of organic matter in a river-dominated estuary (Winyah Bay, SC, USA). Estuar. Coast. Shelf Sci. 57: 1023-1048. [ Links ]
Grasshoff K, Ehrhardt M, Kremling K. 1983. Methods of Seawater Analysis. Verlag Chemie, Weinheim. [ Links ]
Harrison JA, Nina C, Seitzinger SP. 2005. Global patterns and sources of dissolved organic matter export to the coastal zone: Results from a spatially explicit, global model. Global Biogeochem. Cycles 19: 1-16. [ Links ]
Hopkinson C, Vallino J. 1995. The relationships among Man's activities in watersheds and estuaries: A model of runoff effects on patterns of estuarine community metabolism. Estuaries 18: 598-621. [ Links ]
ISO. 1990. Water Quality: Detection and Enumeration of Coliform Bacteria, Thermotolerant Coliform Bacteria and Escherichia coli, ISO 9308/1. International Organization for Standardization, Switzerland. [ Links ]
Jones MN. 1984. Nitrate reduction by shaking with cadmium: Alternative to cadmium columns. Water Res. 18: 643-646. [ Links ]
Lewis WM, Melack JM, Mcdowell WH, Mcclain M, Richey JE. 1999. Nitrogen yields from undisturbed watersheds in the Americas. Biogeochemistry 46:149-162. [ Links ]
Ludwig W, Probst JL, Kempe S. 1996. Predicting the oceanic input of organic carbon by continental erosion. Global Biogeochem. Cycles 10: 23-41. [ Links ]
Meybeck M. 1982. Carbon, nitrogen, and phosphorus transport by world rivers. Am. J. Sci. 282 : 401-450. [ Links ]
Moran MA, Sheldon WM, Sheldon JE. 1999. Biodegradation of riverine dissolved organic carbon in five estuaries of the southeastern United States. Estuaries 22: 55-64. [ Links ]
Mortazavi B, Iverson R, Huang W, Lewis G, Caffrey JM. 2000. Nitrogen budget of Apalachicola Bay, a bar-built estuary in the northeastern Gulf of Mexico. Mar. Ecol. Prog. Ser. 195: 1-14. [ Links ]
Nagata T. 2000. Production mechanisms of dissolved organic matter. In: Kirchman DL (ed.), Microbial Ecology of the Oceans. Wiley-Liss, New York, pp. 121-152. [ Links ]
Nixon SW, Ammerman JW, Atkinson LP, Berounsky VM, Billen G, Boicourt WC, Boynton WR, Church TM, DiToro DM, Elmgren R, Garber JH, Giblin AE, Jahnke RA, Owens NJP, Pilson MEQ, Seitzinger SP. 1996. The fate of nitrogen and phosphorus at the land-sea margin of the North Atlantic Ocean. Biogeochemistry 35: 141-180. [ Links ]
Raymond PA, Bauer JE. 2000. Bacteria consumption of DOC during transport through a temperate estuary. Aquat. Microb. Ecol. 22: 1-2. [ Links ]
Ryther JH, Dunstan WM. 1971. Nitrogen, phosphorus and eutrophication in the coastal marine environment. Science 171: 1008-1013. [ Links ]
Seitzinger SP, Sanders RW. 1997. Contribution of dissolved organic nitrogen from rivers to estuarine eutrophication. Mar. Ecol. Prog. Ser. 159: 1-12. [ Links ]
Seitzinger SP, Sanders RW, Styles R. 2002. Bioavailability of DON from natural and anthropogenic sources to estuarine plankton. Limnol. Oceanogr. 47: 353-366. [ Links ]
Spitz A, Ittekkot V. 1991. Dissolved and particulate organic matter in rivers. In: Mantora RFC, Martin JM, Wallast R (eds.), Ocean Margin Processes in Global Change. Wiley, pp. 5-16. [ Links ]
Uncles RJ, Bloomer NJ, Frickers PE, Griffiths ML, Harris C, Howland RJM, Morris AW, Plummer DH, Tappin AD 2000. Seasonal variability, temperature, turbidity and suspended chlorophyll in the Tweed Estuary. Sci. Total Environ. 251: 115-124. [ Links ]
Vieira MEC, Bordalo AA. 2000. The Douro Estuary (Portugal): A mesotidal salt wedge. Oceanol. Acta 23: 585-594. [ Links ]
Vitousek PM, Howarth RW. 1991. Nitrogen limitation on land and in the sea: How can it occur? Biogeochemistry 13: 87-115. [ Links ]
Zar JH. 1996. Biostatistical Analysis. Prentice-Hall, Englewood Cliffs, New Jersey. [ Links ]
Notas
Traducido al español por Manuel Gardea-Ojeda.

