










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkCiencias marinas
versão impressa ISSN 0185-3880
Cienc. mar vol.34 no.1 Ensenada Mar. 2008
Artículos
Un patrón de periodicidad regular en las capturas mensuales de jibia (Dosidicus gigas) a lo largo de la costa de Chile (2002-2005)
A regular pattern of periodicity in the monthly catch of jumbo squid (Dosidicus gigas) along the Chilean coast (2002-2005)
MJ Zúñiga1, LA Cubillos1*, C Ibáñez1, 2
1 Laboratorio Evaluación de Poblaciones Marinas (EPOMAR), Departamento de Oceanografía, Facultad de Ciencias Naturales y Oceanográficas, Universidad de Concepción, Casilla 160-C, Concepción, Chile. * E-mail: lucubillos@udec.cl
2 Instituto de Ecología y Biodiversidad, Departamento de Ciencias Ecológicas, Facultad de Ciencias, Universidad de Chile, Casilla 653, Santiago, Chile.
Recibido en abril de 2007.
Aceptado en diciembre de 2007.
Resumen
Se detectó un patrón de periodicidad estacional en los datos mensuales de captura de Dosidicus gigas a lo largo de la costa chilena, durante el más reciente periodo de gran abundancia de la especie en la región. Los datos de capturas mensuales, que cubren de 2002 a 2005, fueron agrupados en tres grandes regiones, donde la tendencia de cada serie de tiempo fue eliminada utilizando un suavizador "loess". Se calcularon los residuales entre la curva original y la tendencia "loess", y éstos se analizaron utilizando técnicas de autocorrelación y correlación cruzada con diferentes retrasos de tiempo, así como a través del ajuste de un modelo trigonométrico para detectar oscilaciones estacionales. Se observó un patrón coherente en los residuales mensuales de corto plazo, revelando un proceso similar subyacente en los datos de captura, con un ciclo de 6 meses. Se postula que las fluctuaciones estacionales e interanuales observadas en los registros de captura podrían estar relacionadas con el éxito reproductivo de la especie a través de la ocurrencia de dos ciclos reproductivos por año, y de aquí la generación de dos cohortes por año en promedio.
Palabras clave: Dosidicus gigas, datos de captura, patrón estacional, Chile.
Abstract
A regular seasonal pattern of periodicity was detected in monthly catch data for Dosidicus gigas during the most recent period of high abundance of the species off Chile. Monthly catch data covering from 2002 to 2005 were grouped into three large zones, and the trend of the time series was removed by using loess smoother. The residuals between the original and the loess trend curve were computed and analyzed using autocorrelation and cross-correlation at different lags, and a trigonometric model was fitted to detect seasonal oscillation. A coherent pattern in the short-term monthly residuals was observed in the three time series, revealing a similar underlying process in the catch data, with a six-month cycle. We postulate that the seasonal and interannual fluctuations observed in the catch records could be related to the reproductive success of the species through the average occurrence of two reproductive cycles per year, and hence the generation of two cohorts per year.
Key words: Dosidicus gigas, catch data, seasonal pattern, Chile.
Introducción
La jibia Dosidicus gigas (Orbigny 1835) es uno de los calamares omastréfidos más abundantes en el océano Pacífico Oriental, y se ha descrito como un componente importante del ecosistema tropical, particularmente en el sistema de surgencia de Perú y en el Golfo de California, dónde exhibe la mayor abundancia relativa (Anderson y Rodhouse 2001, Argüelles et al. 2001, Markaida y Sosa-Nishizaki 2003, Nigmatullin et al. 2001, Diaz-Uribe et al. 2006, Bazzino et al. 2007).
Como la mayoría de los cefalópodos conocidos, las poblaciones de omastréfidos muestran una gran variabilidad en su abundancia (Anderson y Rodhouse 2001, Boyle y Boletzky 1996, Taipe et al. 2001) debido principalmente a sus características biológicas tales como rápido crecimiento, maduración temprana, vida corta, alta capacidad migratoria, y complejos patrones de reclutamiento (Boyle y Boletzky 1996). Estas características han permitido a las poblaciones de omastréfidos habitar en zonas caracterizadas por regimenes oceanográficos amplios en aguas oceánicas y costeras, así como en aguas tropicales y templadas (Anderson y Rodhouse 2001). En el Golfo de California el crecimiento y la mortalidad natural de D. gigas varía entre las estaciones del año, y las cohortes muestran una gran variabilidad interanual en la mortalidad por pesca, en la tasa de explotación anual, en el reclutamiento y en la abundancia media (Nevárez-Martínez et al. 2006, Bazzino et al. 2007). Para aguas peruanas y chilenas se ha postulado que dos cohortes de D. gigas podrían explicar los cambios significativos que se observan en la estructura de longitudes entre estaciones del año (Argüelles et al. 2001, Tafur et al. 2001, Chong et al. 2005, Ibáñez y Cubillos 2007).
Frente de Chile, aunque con poca frecuencia, D. gigas muestra episodios de gran abundancia, por lo que su pesquería ha sido clasificada por Fernández y Vásquez (1995) como efímera. Cabe mencionar que en Chile no existe una pesquería orientada hacia D. gigas, y el último periodo de gran abundancia ha sido reportado a partir de la fauna acompañante de las pesquerías demersales y pelágicas tradicionales que operan tanto en aguas costeras como oceánicas. Según Rocha y Vega (2003), entre 1960 y 1972, se obtuvo una captura máxima anual de 3000 t (en 1964), mientras que de 1973 a 1990 sólo se registraron capturas entre 1 y 9 t. Entre 1992 y 1994 ocurrió un periodo de gran abundancia (Rocha y Vega 2003), y recientemente desde 2002 se ha presentado un nuevo periodo de gran abundancia de D. gigas frente a las costas de Chile, alcanzando capturas totales de 15,000 t en 2003 y de 248,000 t en 2005. Los datos de captura pueden ser cuestionables como representativos de la abundancia ya que varios factores influyen sobre la captura. Sin embargo, dado que en Chile no hay una pesquería orientada a la jibia, los datos de esfuerzo de pesca no están disponibles para obtener la captura por unidad de esfuerzo. La especie ha sido capturada principalmente como fauna acompañante por las flotas artesanal e industrial que pescan otros recursos pesqueros pelágicos y demersales. De hecho, como D. gigas es una especie de vida corta (Argüelles et al. 2001, Nigmatullin et al. 2001, Markaida et al. 2004), se asume que los pescadores pueden detectar rápidamente cohortes abundantes o pobremente reclutadas y, por consiguiente, éstas pueden verse reflejadas bastante bien en la captura.
El reciente incremento en la captura de D. gigas frente a Chile, ocurrido entre 2002 y 2005, podría asociarse al éxito reproductivo de la especie en aguas locales, o a invasiones irregulares masivas hacia el sur desde el rango central de distribución de la especie, tal como lo sugiere Nigmatullin et al. (2001). Según estos autores, tales invasiones podrían ser debidas a migraciones alimentarias activas durante años de gran abundancia. En este trabajo se analizó la periodicidad mensual en la captura registrada a lo largo de la costa chilena. El objetivo es inferir un patrón regular que podría relacionarse con las características biológicas de la especie, tales como el éxito reproductivo o migraciones regulares.
Materiales y métodos
Se usaron datos mensuales de captura que cubren de enero de 2002 a diciembre de 2005, y que fueron obtenidos de las estadísticas de captura oficiales del Servicio Nacional de Pesca de Chile. Las series de tiempo de los datos de captura se agruparon en tres zonas: zona norte (18°20'-25°20' S), zona central (25°20'-33°00' S), y zona sur (33°00' S-43°00' S) (fig. 1). Las series de tiempo que aquí se analizan cubren sólo 48 meses, tiempo considerado suficiente para detectar una periodicidad regular mensual. Además, antes de 2002 los datos de la captura eran nulos, por consiguiente la serie de tiempo que aquí se usó está representando el reciente incremento en la aparición de la jibia frente a Chile.

La representación más simple de un patrón en abundancia o productividad es un incremento o disminución temporal (Spencer y Collie 1997). Para evaluar la tendencia en los datos de la captura, se aplicó un suavizador "loess" (Cleveland 1979) a cada conjunto de datos usando una ventana suavizadora (span = 0.8, grado = 1) a la longitud de la base de datos. Se utilizó el paquete "gam" (Hastie y Tibshirani 1990) para lenguaje y software R (Ihaka y Gentleman 1996) para ajustar la tendencia "loess" a los datos. Los residuos entre el dato original y la tendencia suavizada fueron definidos aquí como de corto plazo, es decir la variación mensual. Después de quitar la tendencia global a los datos de captura, los residuos entre los datos originales y suavizados se analizaron para detectar periodicidad y asociación entre la serie de tiempo. Se aplicó la función de autocorrelación (ACF) a los residuos para detectar la estructura correlacionada de la serie de tiempo en diferentes retrasos (el máximo retraso usado fue de 36 meses). También se aplicó la función de la correlación cruzada para detectar una probable dependencia entre las series de tiempo. Se utilizó el paquete "stats" para R (Hihaka y Gentleman 1996) para calcular las funciones de autocorrelación y correlación cruzada.
Finalmente, la periodicidad regular se evaluó utilizando un modelo trigonométrico de acuerdo con Astudillo y Caddy (1988) y Arancibia et al. (1995), i.e.

donde Yt es la captura en el mes t (t = 1 para enero de 2002, ..., t = 48 para diciembre de 2005), αt, es la tendencia "loess" de cada serie de tiempo, A es la amplitud del ciclo, n y  son constantes relacionadas con la fase. Los parámetros A, n y fueron estimados por regresión no lineal en el paquete "nls" de Venables y Ripley (2002) para el lenguaje y software R (Ihaka y Gentleman 1996).
son constantes relacionadas con la fase. Los parámetros A, n y fueron estimados por regresión no lineal en el paquete "nls" de Venables y Ripley (2002) para el lenguaje y software R (Ihaka y Gentleman 1996).
Resultados
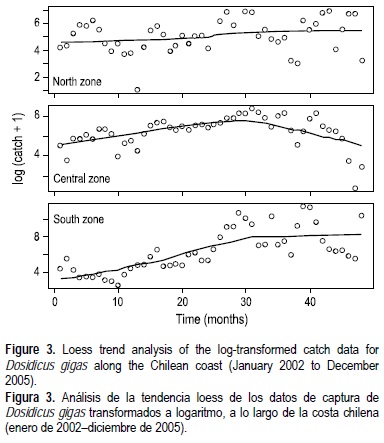
En el periodo de 2002 a 2005 los registros de captura de D. gigas revelaron cambios concurrentes a lo largo de la costa chilena (fig. 2). Según el nivel de captura, las capturas más importantes ocurrieron en la zona sur y central durante 2004 y 2005, y las capturas más bajas fueron registradas en la zona norte. A pesar de la magnitud de la captura en esas tres zonas, los datos de captura transformados a logaritmo mostraron una serie de tiempo estacionaria en la zona norte, junto con un incremento monótono de 2002 a 2004 en los datos de la captura de la zona centro, y un incremento sigmoideo en los datos de captura de la zona sur. Las tendencias fueron bien detectadas mediante la técnica "loess" para cada serie de tiempo (tabla 1). Frente al norte de Chile se observó una tendencia positiva. En cambio, en la zona centro se observó también una tendencia positiva al comienzo, pero después de mayo de 2004 (mes 29) la captura mensual tendió a declinar. En la zona sur se obtuvo una tendencia sigmoidea desde enero de 2002 a diciembre de 2005 (fig. 3).
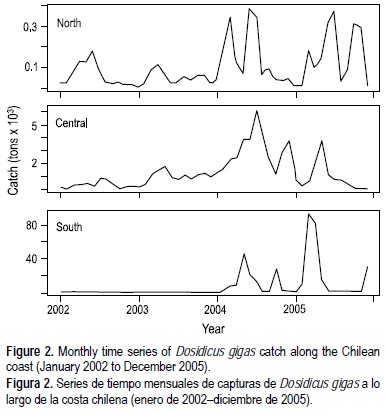
La variación mensual, por lo que se refiere a los residuos entre los datos originales de captura y la tendencia loess (transformados a logaritmo), muestra cambios concurrentes (fig. 4). Aunque hubo diferencias en los residuos de algunos meses, particularmente al final de la serie de tiempo (i.e. 2004 a 2005), se detectó un patrón regular de fluctuación mensual al aplicar la función de autocorrelación. De hecho, la función de autocorrelación reveló una correlación significativa en un retraso de 1 mes, y cambios cada seis meses aproximadamente (fig. 5). La función de correlación cruzada mostró una dependencia entre las tres zonas, con una correlación cruzada significativa sin retraso (lag = 0) entre las zonas norte y centro, y retrasos de 1 y 2 meses entre las zonas norte y sur, y en retrasos de 1, 2 y 3 meses entre las zonas centro y sur (fig. 6).



La periodicidad en la conducta de la variación mensual, evaluada con el método de regresión no lineal periódico, reveló amplitud y fase similares para las tres zonas (tabla 2). Los parámetros del modelo trigonométrico ajustado a cada serie de tiempo fueron significativos (P < 0.01), y los modelos describieron bien los cambios observados en los datos de captura mensuales (fig. 7). Sin embargo, las periodicidades mensuales fueron más similares entre las zonas norte y centro que con la zona sur. De hecho, las zonas norte y centro presentaron una respuesta retardada con respeto a la zona sur, pero en las tres zonas se observa un proceso subyacente similar en los datos de la captura de D. gigas con un ciclo de 6 meses (fig. 8).


Discusión
Se ha observado una fuerte variabilidad interanual en las series de datos de capturas históricas de D. gigas en el Golfo de California (Bazzino et al. 2007), así como también en aguas peruanas (Tafur et al. 2001, Waluda et al. 2006), mientras que en Chile las capturas de D. gigas han sido más bien esporádicas (Rocha y Vega 2003). Algunos autores sugieren que las causas de la variabilidad interanual en las capturas de D. gigas podrían estar relacionadas con cambios en las condiciones ambientales (Névarez-Martínez et al. 2002, Markaida 2006, Waluda et al. 2006), o con cambios en migraciones alimentarias activas, patrones reproductivos o con el éxito del reclutamiento (Ehrhardt et al. 1983, Nigmatullin et al. 2001). A pesar de la variabilidad interanual en los datos de captura, se ha observado un claro patrón estacional de D. gigas en el Golfo de California (Markaida 2006) y frente a Perú (Waluda et al. 2006). En este trabajo se encontro también una periodicidad estacional en la variación mensual de la captura de D. gigas a lo largo de la costa chilena, incluso en la presencia de tendencias y magnitudes interanuales diferentes de la captura entre las zonas. De hecho, las diferencias en la magnitud y tendencia en los datos de la captura están obscureciendo el patrón estacional aquí encontrado a través de los residuos mensuales de captura de D. gigas. En las tres zonas, la captura de esta especie tiende a ser mayor durante abril a julio (otoño del hemisferio del sur) y menor durante octubre a diciembre, lo que configura un ciclo de 6 meses como fue detectado en la función de autocorrelación. Este modelo estacional podría estar relacionado con las características biológicas de la especie, particularmente con procesos biológicos regulares que ocurren dentro de un ciclo anual. De esta manera, los procesos estacionales más relevantes podrían estar relacionados con probables migraciones alimentarias activas estacionales (Nigmatullin et al. 2001), y/o con el ciclo reproductivo y el patrón de reclutamiento estacional subsiguiente (González y Chong 2006, Díaz-Uribe et al. 2007).
Por lo que se refiere al ciclo reproductivo, Nesis (1970, 1983) ha sugerido que el desove ocurre durante la primavera y el verano en el hemisferio del sur. Clark y Paliza (2000) postularon la existencia de dos periodo de desove dentro de un año para D. gigas en el hemisferio del sur, con uno de los picos en otoño (abril-mayo) y otro secundario en primavera entre octubre-noviembre. En Perú, Tafur et al. (2001) también sugirieron que el pico de desove principal tiende ocurrir entre octubre y enero, con un pico secundario entre julio y agosto. Tafur et al. (2001) también señalaron que un índice de reclutamiento mostró un pico en invierno y uno secundario en verano (hemisferio sur). En el Golfo de California Ehrhardt et al. (1983) concluyeron que el stock de D. gigas está compuesto por cohortes múltiples que migran separadamente en algunas ocasiones, pero el reclutamiento a las zonas de pesca principales tiende a ocurrir alrededor de mayo cada año. Díaz-Uribe et al. (2007) apoyaron la hipótesis de cohortes múltiples porque encontraron una gran variabilidad en el índice de madurez, lo que indica maduración asincrónica de los oocitos en la gónada de la hembra. Aunque se han observado especimenes maduros a lo largo del año, en el Golfo de California el máximo de madurez parece prevalecer de mayo a octubre (Bazzino et al. 2007). En cambio, Morales-Bojórquez et al. (2001) comunicaron la existencia de tres cohortes diferentes frente a la costa de Guaymas, Sonora, estimando la capturabilidad a cada longitud de D. gigas. En aguas chilenas, recientemente González y Chong (2006) analizaron la maduración de especimenes de D. gigas y postularon la presencia de por lo menos dos cohortes de la especie para explicar las diferencias en la maduración de los especimenes observada entre invierno y primavera.
Nosotros encontramos un patrón regular en la periodicidad estacional de la captura de D. gigas a lo largo de la costa chilena. En correspondencia con los dos periodos de desove que se han informado para la especie podría suponerse que la fluctuación estacional e interanual observada en los registros de captura podría relacionarse con el éxito reproductivo a través de la ocurrencia media de dos ciclos reproductivos por año, y de aquí la generación de dos cohortes por año independientemente de la magnitud de este patrón de reclutamiento. La existencia de dos cohortes por año también ha sido inferida de los datos de frecuencia de longitud de Chong et al. (2005) e Ibáñez y Cubillos (2007). Probablemente una de las cohortes domina en la abundancia y produce el patrón estacional observado en los datos de la captura.
Por lo que se refiere a las migraciones alimentarias activas estacionales de D. gigas a lo largo de la costa chilena, Nigmatullin et al. (2001) han postulado que esta especie extiende su distribución horizontal desde el Pacífico oriental central hacia el norte (Alaska) y sur de Chile durante periodos de gran abundancia. Nuestros resultados hacen pensar en una respuesta retardada en los residuos de la captura a lo largo de la costa chilena. De hecho, teniendo en cuenta la función de correlación cruzada, solamente hay una respuesta retardada entre las zonas con la zona sur, mismo que también es evidente en los resultados obtenidos con el modelo de periodicidad trigonométrico. Si las migraciones estacionales activas son el principal proceso subyacente en los datos de captura, entonces el proceso podría empezar en la zona sur y ser observado uno a tres meses después en las zonas centro y norte. Este argumento es contradictorio con lo postulado por Nigmatullin et al. (2001), y exige investigar más las probables migraciones a lo largo de la costa chilena. Por mientras, nosotros postulamos que el proceso estacional regular subyacente en los datos de captura parece más relacionado con la existencia de dos periodos reproductivos y con el patrón de reclutamiento subsiguiente que consiste en la generación de dos cohortes por año en promedio.
Agradecimientos
Agradecemos a Hugo Arancibia y Milton Pedraza por su colaboración y ayuda en la revisión de los resultados de nuestra investigación. Gracias también a M Gálvez por proveer los datos de captura utilizados en este trabajo. Estamos agradecidos de los revisores anónimos cuyos comentarios fueron esenciales para mejorar este trabajo.
Referencias
Anderson CIH, Rodhouse PG. 2001. Life cycles, oceanography and variability: Ommastrephid squid in variable oceanographic environments. Fish. Res. 54: 133-143. [ Links ]
Arancibia H, Alarcón R, Cubillos L, Arcos D. 1995. A landing forecast for horse mackerel, Trachurus symmetricus murphyi (Nichols, 1920) off central Chile. Sci. Mar. 59: 113-117. [ Links ]
Argüelles J, Rodhouse PG, Villegas P, Castillo G. 2001. Age, growth and population structure of the jumbo flying squid Dosidicus gigas in Peruvian waters. Fish. Res. 54: 51-61. [ Links ]
Astudillo A, Caddy JF. 1988. Periodicidad de los desembarcos de merluza (Merluccius merluccius) y salmonete (Mullus sp.) en la isla de Mallorca. In: Wyatt T, Larrañeta MG (eds.), International Symposium on Long-Term Changes in Marine Fish Populations, Vigo, 1986, pp. 221-234. [ Links ]
Bazzino G, Salinas-Zavala C, Markaida U. 2007. Variability in the population structure of jumbo squid (Dosidicus gigas) in Santa Rosalía, central Gulf of California. Cienc. Mar. 33: 173-186. [ Links ]
Boyle P, Boletzky S. 1996. Cephalopod populations: Definitions and dynamics. Philos. Trans. R. Soc. Lond. B 351: 985-1002. [ Links ]
Chong J, Oyarzún C, Galleguillos R, Tarifeño E, Sepúlveda RD, Ibáñez CM. 2005. Parámetros biológico-pesqueros de la jibia, Dosidicus gigas (Orbigny, 1835) (Cephalopoda: Ommastrephidae) frente a la costa de Chile central (29°S y 40°S) durante el período 1993-1994. Gayana (Concepción) 69: 319-328. [ Links ]
Clarke R, Paliza O. 2000. The Humboldt Current squid Dosidicus gigas (Orbigny, 1835). Rev. Biol. Mar. Oceanogr. 35: 1-39. [ Links ]
Cleveland WS. 1979. Robust locally weighted regression and soothing scatterplots. J. Am. Stat. Assoc. 74: 829-836. [ Links ]
Díaz-Uribe JG, Hernández-Herrera A, Morales-Bojórquez E. 2006. Histological validation of the gonadal maturation stages of female jumbo squid (Dosidicus gigas) in the Gulf of California, Mexico. Cienc. Mar. 32: 23-31. [ Links ]
Ehrhardt NM, Jacquemin P, García F, González G, López J, Ortiz J, Solís A. 1983. On the fishery and biology of the giant squid Dosidicus gigas in the Gulf of California, Mexico. In: Caddy JF (ed.), Advanced in Assessment of World Cephalopod Resources. FAO Fish. Tech. Pap. 231, Rome, pp. 306-340. [ Links ]
Fernández F, Vásquez JA. 1995. La jibia gigante Dosidicus gigas (Orbigny, 1835) en Chile: Análisis de una pesquería efímera. Estud. Oceanol. 14: 17-21. [ Links ]
González P, Chong J. 2006. Biología reproductiva de Dosidicus gigas D'Orbigny 1835 (Cephalopoda, Ommastrephidae) en la zona norte-centro de Chile. Gayana 70: 65-72. [ Links ]
Hastie T, Tibshirani R. 1990. Generalized Additive Models. Chapman and Hall, New York, 559 pp. [ Links ]
Ibáñez CM, Cubillos LA. 2007. Seasonal variation in the length structure and reproductive condition of jumbo squid Dosidicus gigas (d'Orbigny, 1835) off central-south Chile. Sci. Mar. 71: 123-128. [ Links ]
Ihaka R, Gentleman R. 1996. R: A language for data analysis and graphics. J. Comp. Graph. Stat. 5: 299-314. [ Links ]
Markaida U. 2006. Food and feeding of jumbo squid Dosidicus gigas in the Gulf of California and adjacent waters after the 1997-98 El Niño event. Fish. Res. 79: 16-27. [ Links ]
Markaida U, Sosa-Nishizaki O. 2003. Food and feeding habits of jumbo squid Dosidicus gigas (Cephalopoda: Ommastrephidae) from the Gulf of California, Mexico. J. Mar. Biol. Assoc. UK 83: 1-16. [ Links ]
Markaida U, Quiñónez-Velásquez C, Sosa-Nishizaki O. 2004. Age, growth and maturation of jumbo squid Dosidicus gigas (Cephalopoda: Ommastrephidae) from the Gulf of California, Mexico. Fish. Res. 66: 31-47. [ Links ]
Morales-Bojórquez E, Martínez-Aguilar S, Arreguín-Sánchez F, Nevárez-Martínez MO. 2001. Estimations of catchability-at-length for the jumbo squid (Dosidicus gigas) fishery in the Gulf of California, Mexico. CalCOFI Rep. 42: 167-171. [ Links ]
Nesis KN. 1970. The biology of the giant squid of Peru and Chile, Dosidicus gigas. Okeanologiya 10: 140-152. [ Links ]
Nesis KN. 1983. Dosidicus gigas. In: Boyle PR (ed.), Cephalopod Life Cycles. Vol. I. Species Accounts. Academic Press, London, pp. 215-231. [ Links ]
Nevárez-Martínez MO, Rivera-Parra GI, Morales-Bojórquez E, López-Martínez J, Lluch-Belda DB, Miranda-Mier E, Cervantes-Valle C. 2002. The jumbo squid Dosidicus gigas fishery of the Gulf of California and its relation to environmental variability. In: Salinas S, Urban JH, Arntz WE (eds.), Extended Abstracts of the El Niño Symposium & Workshop. Invest. Mar. 30: 193-194. [ Links ]
Nevárez-Martínez MO, Méndez-Tenorio FJ, Cervantes-Valle C, López-Martínez J, Anguiano-Carrasco ML. 2006. Growth, mortality, recruitment, and yield of the jumbo squid (Dosidicus gigas) off Guaymas, Mexico. Fish. Res. 79: 38-47. [ Links ]
Nigmatullin CM, Nesis KN, Arkhipkin AI. 2001. A review of the biology of the jumbo squid Dosidicus gigas (Cephalopoda: Ommastrephidae). Fish. Res. 54: 9-19. [ Links ]
Rocha F, Vega VM. 2003. Overview of the cephalopod fisheries in Chilean waters. Fish. Res. 60: 151-159. [ Links ]
Rodhouse PG. 2001. Managing and forecasting squid fisheries in variable environments. Fish. Res. 54: 3-8. [ Links ]
Spencer PD, Collie JS. 1997. Patterns of population variability in marine fish stocks. Fish. Oceanogr. 6: 188-204. [ Links ]
Tafur R, Villegas P, Rabí M, Yamashiro C. 2001. Dynamics of maturation, seasonality of reproduction and spawning grounds of the jumbo squid Dosidicus gigas (Cephalopoda: Ommastrephidae) in Peruvian waters. Fish. Res. 54: 33-50. [ Links ]
Taipe A, Yamashiro C, Mariategui L, Rojas P, Roque C. 2001. Distribution and concentrations of jumbo flying squid (Dosidicus gigas) off the Peruvian coast between 1991 and 1999. Fish. Res. 54: 21-32. [ Links ]
Venables WN, Ripley BD. 2002. Modern Applied Statistics with S. 4th ed. Springer-Verlag, New York. [ Links ]
Waluda CM, Yamashiro C, Rodhouse PG. 2006. Influence of the ENSO cycle on the light-fishery for Dosidicus gigas in the Peru Current: An analysis of remotely sensed data. Fish. Res. 79: 56-63. [ Links ]

