










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkCiencias marinas
versão impressa ISSN 0185-3880
Cienc. mar vol.34 no.1 Ensenada Mar. 2008
Artículos
Peces sin agua: Validación y aplicación de δ18O en los otolitos de Totoaba macdonaldi
Fish without water: Validation and application of δ18O in Totoaba macdonaldi otoliths
K Rowell1*, C True2, KW Flessa1, DL Dettman1
1 Department of Geosciences, University of Arizona, Tucson, Arizona 85721, USA.
2 Universidad Autónoma de Baja California, Ensenada, Baja California, México.
* Corresponding author's current address: Department of Biology, University of Washington, 528 Kincaid Hall, Box 351800, Seattle, WA 98195-1800, USA. E-mail: rowellk@u.washington.edu.
Recibido en abril de 2007.
Aceptado en enero de 2008.
Resumen
La geoquímica de otolitos de peces es una herramienta muy valiosa para reconstruir sus condiciones ambientales, migraciones y ciclos de vida. En este estudio se validó la relación entre la temperatura, la composición de los isótopos de oxígeno del agua (δ18O) y del otolito de Totoaba macdonaldi, una especie en peligro de extinción, tanto en individuos cultivados en condiciones controladas como en individuos encontrados en el medio natural. Este tipo de validación es muy importante para la reconstrucción del hábitat. Al comparar los valores de δ18O en la porción natal de los otolitos de especímenes de totoaba modernos y previos a la construcción de presas (~1000-4500 años AP), se examinó la hipótesis de que la totoaba usaba el estero del Río Colorado como sitio de crianza antes de que el río dejara de fluir debido a la sobreexplotación del agua. Los resultados indican que es posible predecir los valores de δ18O de los otolitos tanto en condiciones de laboratorio como en el medio natural. Los otolitos de las totoabas antes de la construcción de presas tienen valores natales de δ18O significativamente menores que los valores predecibles, lo que indica que estas diferencias en los valores de δ18O son el resultado de un cambio en los valores de δ18O en el agua, el cual a su vez es el resultado de la reducción del flujo del Río Colorado hacia la zona de crianza de la totoaba antes de la construcción de presas. Se concluye que el flujo del Río Colorado fue un componente principal del hábitat de crianza de la totoaba antes de la construcción de presas y la reducción del flujo del río. Estos resultados indican que es apropiado usar los otolitos para reconstruir información ecológica importante y el ciclo de vida de la totoaba, lo que resulta relevante para la investigación sobre esta especie en peligro de extinción.
Palabras clave: otolitos, isótopos de oxígeno, estuario del Río Colorado, totoaba, estuario.
Abstract
The geochemistry of fish otoliths is a valuable tool for reconstructing environmental conditions, migrations and life histories. In this study we validate the relationship between temperature, the water oxygen isotope composition (δ18O) and otolith δ18O for the endangered Totoaba macdonaldi, raised under controlled aquaculture conditions and in the field. This type of validation is instrumental for habitat reconstruction. By comparing δ18O values in the natal portion of totoaba otoliths from modern and predam specimens ~1000—4500 yr BP, we test the hypothesis that the totoaba used the Colorado River estuary as a nursery site before the river was over-allocated. We found that otolith δ18O could be predicted in a laboratory setting as well as in the wild. Totoaba otoliths from before river diversion had drastically lower natal δ18O values than predicted values, indicating that these differences in δ18O values are the result of a change in the water δ18O, a consequence of diverting the isotopically negative Colorado River flow from the totoaba's nursery grounds. We conclude that the Colorado River flow was a major component of the totoaba's nursery habitat before river diversions. These results are pertinent to ongoing research on this endangered fish, using otoliths to piece together important ecological and life history information.
Key words: otoliths, oxygen isotopes, Colorado River estuary, totoaba, estuary.
Introducción
La determinación de información ecológica básica para muchas especies marinas es una tarea difícil (Dayton et al. 1998). Los ecosistemas marinos han sido fuertemente influenciados por el impacto antropogénico y la pesca industrial desde mucho antes de que comenzaran los monitoreos ecológicos. En el caso de la Totoaba macdonaldi (Gilbert 1891) (Teleostei: Sciaenidae), una especie declarada en peligro de extinción, el intensivo esfuerzo de pesca sobre la población adulta (de 1930 a 1975), en combinación con la desviación de casi todo el flujo del Río Colorado (1963) que vertía sus aguas al único hábitat de desove y crianza de esta especie resultó ser crítico. A pesar de la conexión correlativa entre la declinación de la pesquería y las alteraciones al estuario del Río Colorado, no hay evidencia empírica de que la totoaba utilizaba este hábitat estuarino salobre como criadero (Flanagan y Hendrickson 1976, Cisneros-Mata et al. 1995, Lercari y Chávez 2007), ni tampoco se ha estudiado la influencia de la modificación del hábitat en la declinación de esta pesquería (Avila-Serrano et al. 2006). En este estudio se realizaron calibraciones en el laboratorio de las razones de los isótopos ambientales de oxígeno y la temperatura, y la razón de los isótopos de oxígeno en los otolitos (piedras del oído) para poder determinar si los juveniles de la totoaba habitaban el agua salobre que existía en el estero debido al vertido del Río Colorado. Esta reconstrucción ambiental del hábitat de los juveniles previo a la construcción de presas es el primer paso para entender la relación entre la totoaba y el Río Colorado.
Los otolitos de peces se han convertido en un recurso popular para la reconstrucción de sus ambientes y ciclos de vida (Kalish 1991, Thorrold et al. 1997, Patterson 1998, Campana y Thorrold 2001, Weidman y Millner 2001, Rowell et al. 2005). Las razones de los isótopos estables de oxígeno conforme se incrementa el CaCO3 en los otolitos con el crecimiento, pueden proporcionar un registro cronológico de los parámetros ambientales asociados con los diferentes estadios del ciclo de vida de los peces proporcionando una visión de los ambientes antiguos y modernos donde se desarrollan sus poblaciones y su interacción con estos ambientes (Patterson 1998, Campana y Thorrold 2001). Las variaciones de δ18O (O18/O16 en comparación con un estándar) en los otolitos han sido utilizadas para detectar cambios climáticos (Patterson 1998) y migraciones anádromas (Dufour et al. 1992), y para discriminar entre hábitats salobres, oceánicos y de agua dulce (Rowell et al. 2005). Para los peces que permanecen en ambientes meramente marinos, los valores de δ18O en otolitos presentan una relación predecible con la temperatura (ver Kalish 1991, Thorrold et al. 1997, Weidman y Millner 2000). La discrepancia entre las relaciones publicadas entre la temperatura del agua, δ18O del agua y δ18O del otolito sugiere que éstas pueden ser específicas de cada especie. Aunque las relaciones son similares, es difícil elegir una de ellas para las reconstrucciones basadas en los isótopos de oxígeno sin examinar el taxón empleado en cada estudio.
En este estudio se probó la hipótesis de que los criaderos de T. macdonaldi, en peligro de extinción, se han visto alterados significativamente debido a la desviación de las aguas del Río Colorado. Para ello se examinó si las relaciones publicadas sobre la variación causada por la temperatura en el valor de δ18O de los otolitos puede predecir los valores de δ18O observados en los otolitos de las totoabas en condiciones controladas y en el medio natural. Al confirmar la predictibilidad de los valores de δ18O en los otolitos in situ en las condiciones actuales, se podrán interpretar los valores de δ18O de la porción natal de los otolitos de la totoaba en el pasado, cuando el estuario del Río Colorado no estaba controlado por la regulación de sus aguas río arriba. Este estudio tiene dos objetivos principales (fig. 1): (1) demostrar que se pueden utilizar las relaciones publicadas entre temperatura y δ18O de los otolitos para interpretar los valores natales de δ18O en los otolitos de la totoaba a partir de peces que vivieron antes de la regulación de las aguas río arriba (ca. 1000-4500 años AP; Rowell 2006), y (2) determinar si el flujo del Río Colorado ha sido un componente principal del hábitat de crianza de la totoaba. Este análisis retrospectivo del hábitat de crianza histórico de T. macdonaldi en peligro de extinción puede proporcionar directrices para la recuperación de esta especie.
Antecedentes
La totoaba es un depredador marino de gran tamaño (hasta 2 m de largo) y longevo (Flanagan y Hendrickson 1976, Román-Rodríguez y Hammann 1997). Es endémica del Alto Golfo de California y desova exclusivamente en la boca del Río Colorado durante la primavera (Román-Rodríguez y Hammann 1997). En 1976, después de años de pesca intensiva en su hábitat de desove y poco después de que los flujos fluviales disminuyeron a raíz de su desvío río arriba (1965), la totoaba fue declarada como especie en peligro de extinción en México y los Estados Unidos (Flanagan y Hendrickson 1976, Cisneros-Mata et al. 1995).
El éxito en los esfuerzos para conservar la totoaba ha sido limitado, a pesar de que sus áreas de desove y crianza han estado protegidas desde 1993 (Cisneros-Mata et al. 1995). Durante los años noventa, como parte de un programa de repoblación, la totoaba fue cultivada exitosamente en cautiverio a partir de unos reproductores silvestres iniciales. Los peces fueron capturados y trasladados a la Universidad Autónoma de Baja California (UABC), en Ensenada (México), donde se mantuvieron en cautiverio en condiciones controladas, simulando un fotoperiodo y una temperatura naturales para inducir la maduración y el desove (True et al. 1997). A pesar de la protección de su hábitat, su veda permanente y los esfuerzos de repoblación, hay poca evidencia de que la especie se esté recuperando. Otros factores que pueden estar amenazando a la totoaba son las alteraciones significativas que ha sufrido su hábitat de crianza (Flanagan y Hendrickson 1976, Cisneros-Mata et al. 1995). Hasta ahora existe poca evidencia empírica del uso histórico por la totoaba del hábitat estuarino generado por el Río Colorado.
Sitio de estudio
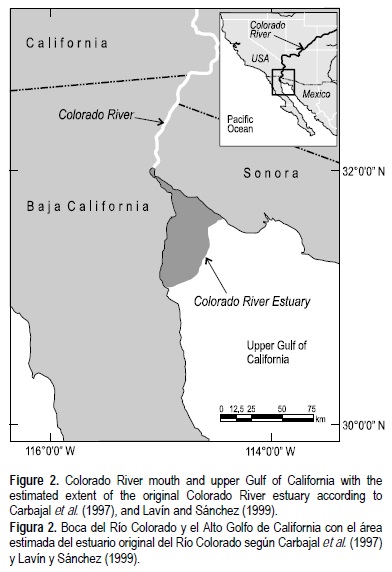
El Río Colorado ejercía una gran influencia sobre el Alto Golfo de California. Tenía flujos grandes y variables que mantenían un estuario que cubría ~4000 km2 de la parte más interna del Golfo de California (Lavín y Sánchez 1999) y una zona de mezcla que se extendía hasta ~65 km de la boca del río (Rodriguez et al. 2001) (fig. 2). Durante el último siglo el Río Colorado fue represado, desviado y sobreasignado; hoy en día, apenas fluye al Golfo de California. Los efectos de haber desviado el agua dulce han sido patentes conforme el Golfo de California se ha vuelto más salino (Lavín et al. 1998). Aun así, los efectos ecológicos a causa de la eliminación de los flujos de agua dulce siguen sin dilucidarse.
Material y métodos
Se utilizaron tres juegos de otolitos: doce otolitos de totoabas juveniles cultivadas en el laboratorio de la UABC, cinco de totoabas silvestres que vivieron antes de la construcción de presas en el Río Colorado, y cinco de totoabas silvestres modernas. Los otolitos usados para la validación en el laboratorio fueron tomados de peces desovados y criados en condiciones controladas y conocidas. Las razones de los isótopos de oxígeno de los otolitos de las totoabas cultivadas en el laboratorio fueron comparadas con los valores pronosticados mediante tres relaciones entre temperatura y δ18O del agua y δ18O de la aragonita del otolito, publicadas por Thorrold et al. (1997), Weidman y Millner (2000) y Kalish (1991). Para probar si los valores de δ18O de los otolitos pueden ser pronosticados en el campo, se compararon los valores natales de δ18O de otolitos de totoabas silvestres de tiempos tanto prehistóricos como modernos, y las diferencias se interpretaron en términos de diferencias ambientales.
Los otolitos de totoabas anteriores al represado del río se obtuvieron de dos conchales diferentes en la costa del Alto Golfo de California. Cuatro especímenes pertenecientes a la Colección de Vertebrados Marinos del Instituto Oceanográfico Scripps (EUA) fueron recolectados en 1955, cerca de San Felipe, Baja California (México), en un conchal fechado (por 14C) por una concha marina (1885 años AP) y tres edades de carbón vegetal (1065-1220 años AP) (ver Goodfriend y Flessa (1997) para mayor información). El quinto otolito fue donado por el Instituto Tecnológico y de Estudios Superiores de Monterrey, campus Guaymas (Mexico), a la Colección de Peces de la Universidad de Arizona (EUA), y fue obtenido de la superficie de un conchal en Bahía San Jorge, Sonora (México). Este espécimen fue fechado directamente en 4580 ± 130 14C años AP. Rowell (2006) ha realizado una descripción más completa de estos otolitos, los métodos de fechado y los perfiles de δ18O para todo su ciclo de vida.
Los otolitos posteriores al represado del río fueron extraidos de individuos de T. macdonaldi como parte de un estudio de monitoreo de la población realizado en 1987 y 1990, y empleados para estudiar la edad y crecimiento de las totoabas (Román-Rodríguez y Hammann 1997). Estos especímenes fueron posteriormente donados por el Instituto del Medio Ambiente y el Desarrollo Sustentable del Estado de Sonora a la Colección de Peces de la Universidad de Arizona. En este trabajo se utilizó la porción natal de estos otolitos, que representó los años de 1963 a 1976.
Análisis de δ18O en los otolitos
La proporción o razón de isótopos de oxígeno (18O/16O) en la aragonita depositada en los otolitos está en cuasi equilibrio con la del medio acuático (Kalish 1991, Thorrold et al. 1997, Weidman y Millner 2000). Un cambio en esta razón (expresado como δ18O) es resultado de un cambio en la temperatura ambiente o en el δ18O del agua en que habita el animal. Cambios en el origen del agua (e.g., tierra adentro vs marina) también pueden ocasionar cambios en los valores de δ18O; sin embargo, si un pez es restringido a condiciones marinas, el ciclo de δ18O en el otolito permanecerá dentro de los límites controlados por la temperatura estacional. El ciclo de δ18O en los otolitos de peces que habitan ambientes estuarinos salobres estará controlado por la temperatura, pero sus valores serán más negativos que los de un registro marino típico debido a la adición de agua de río isotópicamente negativa al estuario (Ingram et al. 1996, Dettman et al. 2004). Por tanto, el δ18O en otolitos puede ser utilizado para documentar la temperatura del agua, así como la presencia y cantidad de aguas isotópicamente distintas (Dettman et al. 2004, Jamieson et al. 2004, Rowell et al. 2005).
Cuando se conocen el δ18O del agua y la temperatura, se pueden realizar predicciones del δ18O de la aragonita de los otolitos. Aunque varios estudios documentan la variación en el δ18O del otolito generada por la temperatura, cada relación publicada difiere en la influencia relativa que tiene la temperatura sobre la fraccionación de δ18O para otolitos. Se compararon los promedios observados de la porción natal de 12 otolitos con tres valores pronosticados usando las siguientes relaciones, donde T es el promedio de la temperatura del agua (registrada en °C), δ18Ootolito es el promedio de δ18O del otolito expresado como Vienna Pee Dee Belemnite (VPDB), y δ18Oagua es el promedio de δ18O del agua expresado como Vienna Standard Mean Ocean Water (VSMOW):

Thorrold et al. (1997), con base en la gurrubata Micropogonias undulatus cultivada a temperaturas entre 18°C y 25°C.

Weidman y Millner (2000), con base en el bacalao del Atlántico Gadus morhua cultivado a temperaturas entre 2.5°C y 10.5°C.

Kalish (1991), con base en el salmón austrialiano Arripis trutta cultivado a temperaturas entre 13°C y 22°C.
Validación en el laboratorio
En vista de que las totoabas están protegidas federalmente, no se permiten realizar experimentos que requieran manipular los parámetros ambientales y puedan poner en riesgo a los individuos. Por tanto, simplemente se monitorearon las condiciones del agua y se tomaron muestras de agua para el análisis isotópico. Se recolectaron huevos fértiles de totoaba de tanques de reproductores, y se almacenaron en tanques cónicos de 2200 L a una temperatura controlada de 25°C y con flujo de agua de mar estéril (del Pacífico local) a una tasa de 3 L min-1. La alimentación inició cuando las larvas mostraron una boca y un estómago funcional (~4-5 días después de la eclosión). Al día 22 los juveniles en etapa temprana fueron transferidos a tanques más grandes (8000 L) para permitir un crecimiento adecuado. Durante esta etapa se mantuvieron las condiciones de cultivo, aunque el flujo de agua de mar se incrementó a 33 L min-1. La temperatura en ambos tanques se mantuvo entre 24°C y 27°C, simulando el ambiente natural de los alevines en primavera y verano en el norte del Golfo de California (True et al. 1997, 2001). La temperatura se registró cada 30 min con un termógrafo Hobo. Para una descripción más detallada de las condiciones de cultivo larval, ver los trabajos de True et al. (1997, 2001).
Se recolectaron tres juegos de muestras de agua de los tanques de cultivo durante un periodo de dos meses. El δ18O del agua se midió mediante un espectrómetro de masas de doble entrada (Delta-S, Thermo Finnegan, Brema, Alemania) equipado con una unidad automática para balancear CO2-H2O. La estandarización se basó en patrones internos referenciados a VSMOW y Vienna Standard Light Antarctic Precipitation (VSLAP). La precisión para δ18O fue mayor a ± 0.08%.
Se extrajeron los otolitos sagitales (n = 12) de los alevines de totoabas de edad conocida (31-60 días) y se lavaron ultrasónicamente en agua destilada. Ya que los otolitos de los juveniles eran demasiado chicos para obtener un corte delgado (~2 mm), se utilizó un molinillo para exponer una sección transversal dorsoventral. Se tomaron dos muestras de cada otolito: una de la porción natal del otolito y otra del borde exterior del mismo. Se utilizó un microtaladro y una broca de 0.3 mm de diámetro para recolectar las muestras de 30-60 μg. Las muestras se calentaron a 180°C en un horno de vacío durante una hora para eliminar el material volátil. Todas las razones de oxígeno en los valores de carbonato se expresan en relación con el estándar VPDB, y se midieron en el Laboratorio de Isótopos Ambientales del Departamento de Geociencias de la Universidad de Arizona. Los carbonatos fueron analizados usando un espectrómetro de masas Finnigan MAT 252 equipado con un dispositivo automático de muestreo de carbonatos Kiel-III. Las muestras pulverizadas fueron tratadas al vacío con ácido fosfórico deshidratado a 70°C. La medición de la razón isotópica se calibró mediante mediciones repetidas de NBS-19 y NBS-18, y la precisión fue de ± 0.1%.
El valor medio del otolito fue emparejado con el promedio pronosticado usando la temperatura media del agua para el periodo de tiempo que representa cada muestra. Como se conoce el tiempo de desove (porción natal) y muerte (perímetro), se puede estimar el intervalo de tiempo que representa cada muestra (0.3 mm de material), suponiendo un crecimiento constante y midiendo el diámetro del otolito. Se utilizó una prueba t por pares para comparar el promedio pronosticado, usando cada una de las ecuaciones para cada otolito, con el promedio observado.
Validación en el medio natural
Considerando que la validación de la relación entre temperatura, δ18Oagua y δ18Ootolito es más valiosa cuando se puede utilizar para determinar las condiciones ambientales de las totoabas silvestres, se analizó si las ecuaciones utilizadas pueden predecir el 818O en otolitos natales de totoabas silvestres capturadas en condiciones meramente marinas. Se pronosticó el promedio y el intervalo de los valores de δ18Ootolito para la porción natal de los otolitos usando promedios semanales de la temperatura de 1964 a 1999 para el periodo de desove de la totoaba (de marzo a junio) (Rowell et al. 2005). Se supuso que para el Alto Golfo de California, δ18Oagua era similar a la media del agua oceánica, 0.0% para la totoaba moderna. Las estimaciones actuales de δ18Oagua en el Alto Golfo de California son más positivas que tal valor (~ +0.6%) a causa de las altas tasas de evaporación y medio siglo de poca o nula entrada de agua del Río Colorado al estuario (Dettman et al. 2004, Rowell et al. 2005). Durante la evaporación, el O16 se consume con mayor facilididad, lo que ocasiona el enriquecimiento del agua restante con O18. El aporte de agua isotópicamente negativa del Río Colorado (~ -12%) compensaría este enriquecimiento de la evaporación (Dettman et al. 2004). Dado que se conoce el tiempo de captura se puede retrocalcular el año que representa la porción natal: de 1963 a 1976, poco tiempo después de que se construyeron las últimas presas importantes del Río Colorado. Se supone, de manera conservadora, que durante ese tiempo la evaporación (y la falta de flujo fluvial) aún no tenía una influencia tan importante sobre el δ18Oagua en el Alto Golfo de California como la tiene actualmente.
Para determinar si el hábitat de crianza se ha visto alterado significativamente por las derivaciones río arriba, se compararon los intervalos pronosticados de δ18O de los valores natales para la totoaba in situ con los promedios natales de los otolitos (n = 5) de totoabas capturadas mediante un permiso federal en 1987 y 1990 (Román-Rodríguez y Hammann 1997). Se recolectaron muestras del carbonato de los otolitos usando el mismo método utilizado con los peces criados en el laboratorio. Los valores de δ18O observados para las totoabas silvestres modernas deberían de estar dentro del rango de la variación generada por la temperatura durante la temporada de desove. La confirmación de que una relación generada por la temperatura puede predecir el δ18Ootolito en totoabas silvestres permite determinar si las totoabas prehistóricas vivían en un hábitat isotópicamente distinto: uno dominado por la influencia de los flujos de agua isotópicamente negativa del Río Colorado.
Se compararon estos mismos valores pronosticados con los promedios natales observados de los otolitos de las totoabas prehistóricas (n = 5). Los otolitos prehistóricos representan peces que se desarrollaron en condiciones no perturbadas, un periodo cuando la alteración antropogénica del hábitat era mínima en comparación con la actual. Se utilizó el promedio de los dos valores más natales de δ18O en otolitos para cada espécimen; por tanto, cualquier diferencia en el δ18O de la aragonita entre los otolitos anteriores y posteriores a las derivaciones del agua del Río Colorado, secretados a la misma temperatura, es el resultado de un cambio en la influencia relativa de aguas isotópicamente diferentes.
Resultados
Validación en el laboratorio
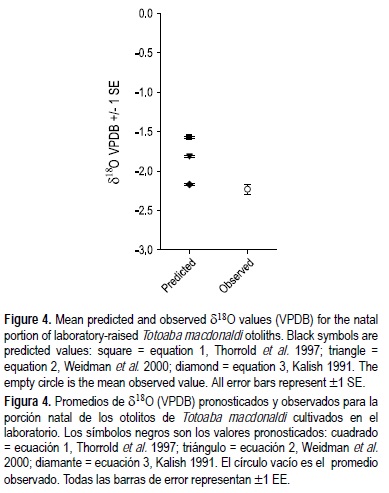
Se encontró que es posible predecir el δ18O de los otolitos en condiciones conocidas. Durante el periodo de crecimiento, los valores de δ18Oagua fueron cada vez más negativos con el tiempo (las muestras dieron -0.5%, -0.6% y -0.7%) y la temperatura se mantuvo relativamente constante (25.3°C ± 0.01 EE). Los valores de δ18Ootolito observados para las totoabas cultivadas en el laboratorio oscilaron entre -1.86% y -2.85%, y el promedio de δ18Ootolito fue de -2.2% ± 0.7 EE (tabla 1). Los promedios para los otolitos de peces con mayor tiempo de crecimiento fueron más negativos; o sea, los peces más viejos presentaban valores de δ18Ootolito más negativos (fig. 3). El intervalo entre los valores pronosticados de δ18Ootolito fue menor a uno por mil (-1.5% a -2.17%), y el promedio pronosticado de δ18Ootolito para las ecuaciones 1, 2 y 3 fue de -1.54%, -1.78% y -2.14%, respectivamente. Los valores observados de δ18Ootolito difirieron significativamente de los valores pronosticados por las ecuaciones 1 (t pareada = -9.98, P < 0.0001) y 2 (t pareada = -6.32, P < 0.0001), pero no de los valores pronosticados por la ecuación 3 (t pareada = -0.927, P = 0.374; fig. 4). El promedio de las diferencias entre los valores pronosticados y los observados fue de 0.66% para la ecuación 1, 0.42% para la ecuación 2 y 0.06% para la ecuación 3.

Validación en el medio natural
Los valores observados de δ18Ootolito fueron -0.84% ± 0.1 EE para las totoabas modernas y -2.99% ± 0.6 EE para las antiguas (tabla 2). Los valores natales pronosticados de δ18Ootolito para los peces silvestres que se desarrollan en aguas marinas y a temperaturas características de su temporada de desove (~16-27°C) varió de +1.5% a -2.11%. Los promedios natales pronosticados de δ18Ootolito para las ecuaciones 1, 2 y 3 fueron -0.05%, -0.25% y -0.18%, respectivamente. Los valores observados para las totoabas silvestres modernas estuvieron dentro del intervalo de los valores pronosticados de δ18Ootolito, pero los valores natales antiguos fueron más de 2% más negativos que los valores modernos observados y más de 0.8% más negativos que el menor valor pronosticado de δ18Ootolito (fig. 5).


Discusión
Cuando se conoce la temperatura y el δ18O del agua, es posible predecir el δ18O del carbonato de los otolitos de la totoaba tanto en el laboratorio como en el medio natural. Los valores observados del isótopo de oxígeno en los otolitos de los peces cultivados coincidieron con los valores pronosticados por la ecuación 3 (Kalish 1991), y en el medio natural, las tres relaciones pronosticaron exitosamente los valores natales para los otolitos modernos en un hábitat marino. La escala de la resolución (estacional) usada para modelar las condiciones del medio natural fue mayor que en el laboratorio y es posible que por ello todas las ecuaciones hayan podido predecir el δ18O natal en otolitos de peces silvestres modernos. El intervalo de ~11°C de temperaturas posibles durante la temporada de desove de la totoaba (marzo a junio), se traduce en un intervalo de valores de δ18O > 3.5%. Los valores natales de δ18O en los otolitos de las totoabas prehistóricas fueron significativamente diferentes de los de los otolitos modernos y de los pronosticados para condiciones totalmente marinas. Esta diferencia indica que los peces previos a la construcción de presas se criaron en un ambiente significativamente diferente. Esta compensación fuertemente negativa evidencia que los peces antiguos se desarrollaban en zonas con influencia fluvial.
En general, los valores de δ18Ootolito para las totoabas cultivadas en el laboratorio fueron más negativos que los valores pronosticados. La compensación negativa entre los valores observados y los pronosticados puede ser resultado de varios factores. Uno podría ser un efecto cinético específico de la especie, en el que durante el crecimiento el 16O se incorpora en el carbonato en una razón mayor a la determinada por la fraccionación lenta en equilibrio. A pesar del pequeño tamaño de muestra, se presentó una relación negativa entre la tasa de crecimiento de la totoaba (mm día-1) y el promedio de δ18O (F1,12 = 4.39, P < 0.0001, r2 = 0.24); sin embargo, Thorrold et al. (1997) no encontraron ninguna relación entre la tasa de crecimiento y δ18Ootolito en la especie M. undulatus. Aunque la fraccionación de δ18O puede verse o no afectada por el crecimiento del otolito, seguramente la temperatura debe tener algún efecto. Dado que el crecimiento de los peces tiende a ser más rápido a temperaturas cálidas, el promedio de δ18O de un otolito puede estar cargado hacia condiciones más cálidas, dando como resultado mayor depósito de material carbonatado a temperaturas mayores y valores de δ18O más negativos. En otras palabras, el δ18O de otolitos simplemente puede estar sesgado hacia las temperaturas óptimas para el crecimiento. Las ecuaciones que actualmente predicen δ18Ootolito no consideran la influencia de la temperatura en la tasa de crecimiento.
Otra posible explicación para la compensación negativa entre los valores observados y los pronosticados es que nuestro muestreo de los otolitos se haya realizado a mayor resolución que el de δ18Oagua y, por tanto, no se pudo caracterizar de manera precisa la química del agua que afecta la predicción. Las muestras representaron, en promedio, de 9 a 10 días de crecimiento. Las muestras de agua se tomaron tres veces a lo largo de 62 días. Los valores de δ18Oagua variaron más de lo anticipado debido a una serie de tormentas invernales y al hecho de que el suministro de agua para los tanques ocurre donde el escurrimiento local hacia el mar podría haber influenciado la señal marina, potencialmente causando una variación a corto plazo sesgada hacia valores de δ18Oagua más negativos. Consecuentemente, nuestro régimen de muestreo del agua pudo haber sido insuficiente para caracterizar los valores de δ18Oagua durante los momentos en que se precipitaba el carbonato del otolito; sin embargo, los valores de δ18Ootolito sí siguieron la misma tendencia hacia valores más negativos observados en las muestras de δ18Oagua durante el periodo de crecimiento.
Los valores natales de δ18O observados en los otolitos de las totoabas silvestres modernas se encuentran dentro del intervalo pronosticado para cada ecuación. Si se supone que estos peces desovaron a la temperatura media de la temporada de desove, hay una compensación negativa entre los valores natales observados y los pronosticados. Claro, es posible que estos individuos hayan desovado más tarde en la temporada. Usando las ecuaciones 1, 2 y 3, se puede retrocalcular a partir de los valores natales observados de los otolitos silvestres modernos, lo que sugiere que la porción natal de los otolitos se desarrolló en aguas a 18-19°C, temperaturas características de finales de abril y principios de mayo. En favor de ello, existe evidencia de que la temporada de desove de la totoaba se ha desplazado hacia finales de la primavera y que se ha visto truncada significativamente por el esfuerzo de pesca selectivo durante las agregaciones reproductivas a principios de la temporada (Flannagan y Hendrickson 1976). Flannagan y Hendrickson (1976) indicaron que antes de que la totoaba fuera protegida, la mayor parte de las capturas se realizaban en los últimos dos días de la temporada de pesca (30 de marzo al 1 de abril). Las temperaturas cálidas usadas durante el cultivo para el desove de las totoabas y la crianza de las larvas también indican que el desove y el éxito del reclutamiento de larvas se pueden ver favorecidos por estas temperaturas más cálidas.
La capacidad de predecir el δ18O del otolito en condiciones ambientales conocidas nos permite determinar si los otolitos de las totoabas que vivían en el pasado (condiciones desconocidas) se desarrollaron en un hábitat isotópicamente diferente. Encontramos que los valores natales de δ18Ootolito de las totoabas prehistóricas fueron considerablemente menores que los valores pronosticados al igual que los valores obtenidos para los especímenes modernos. Aun cuando la porción natal del otolito se hubiera desarrollado durante las mayores temperaturas del año (31°C), los valores pronosticados no serían iguales que los registrados por los peces antiguos. Esto es evidencia directa de que antes de la alteración antropogénica del flujo del río las totoabas desovaban en un ambiente muy diferente: uno con un valor de δ18Oagua significativamente menor, característico de estuarios influenciados por ríos (Rowell et al. 2005).
Al igual que para varias comunidades marinas, existen pocos datos sobre el ecosistema del Alto Golfo de California antes de las perturbaciones antropogénicas, lo que dificulta la validación de las condiciones naturales (Dayton et al. 1998, Jackson et al. 2001). Desafortunadamente son los científicos los que tienen que aportar evidencias de que las alteraciones antropogénicas (sobrepesca, regulación de ríos, desarrollo costero, contaminación) han tenido efectos negativos sobre el ecosistema marino (Dayton 1998). Las técnicas que usan indicadores naturales para documentar las condiciones ambientales naturales (antes de las perturbaciones) y la ecología de los organismos son de inestimable valor para el manejo adecuado de los océanos. Se ha confirmado aquí que el δ18O de los otolitos de totoabas puede resolver algunas de estas lagunas del conocimiento. También se demostró que el δ18O tiene una relación predecible con la temperatura del agua del medio si se conoce el δ18O del agua. Por medio de estas herramientas para investigar el impacto de la reducción del flujo del Río Colorado hacia el Golfo de California, se encontró que la zona de crianza de T. macdonaldi, una especie en peligro de extinción, ha sido profundamente alterada y modificada de ser un estuario salobre a convertirse en uno salino. El éxito en la recuperación de la totoaba probablemente requerirá un flujo estacional apropiado de agua del Río Colorado hacia su histórico estuario.
Agradecimientos
Reconocemos y agradecemos a LM López-Acuña el suministro de las dietas para las larvas, así como a G Sandoval-Garibaldi e I Monay su apoyo en la crianza de las larvas, los muestreos de agua y la recolección de otolitos. Los otolitos fueron donados por la Universidad Autónoma de Baja California (Ensenada) al Museo de Historia Natural de la Universidad de Arizona (UAZ2005-07 al 18). Los otolitos recolectados en 1987 y 1990 (UAZ-S2005-01 al 05) fueron donados por el Instituto del Medio Ambiente y el Desarrollo Sustentable del Estado de Sonora a la Colección de Peces de la Universidad de Arizona. El espécimen antiguo (UAZ-S2005-06) fue donado por el Tecnológico de Monterrey, campus Guaymas. Todos los especímenes donados fueron importados bajo el permiso CITES 06US119621/9 a la Universidad de Arizona. Los especímenes utilizados en este estudio fueron proporcionados por P Reinthal (Colección de Vertebrados de la Universidad de Arizona), LT Findley y MJ Román. Los comentarios y las sugerencias atinadas de JJ Tewksbury y C Cintra-Buenrostro contribuyeron al éxito de este proyecto. Se recibió apoyo financiero de National Science Foundation (EAR-0125025 y INT-0323139), T&E Inc., Southern Arizona Environmental Management, y Marshall Foundation. También agradecemos los comentarios editoriales de dos revisores y S Shephard.
Referencias
Avila-Serrano GE, Flessa KW, Téllez-Duarte MA, Cintra-Buenrostro CE. 2006. Distribution of the intertidal macrofauna of the Colorado River Delta, northern Gulf of California, Mexico. Cienc. Mar. 32: 649-661. [ Links ]
Campana SE, Thorrold SR. 2001. Otoliths, increments and elements: Keys to a comprehensive understanding of fish populations. Can. J. Fish. Aquat. Sci. 58: 30-38. [ Links ]
Carbajal N, Souza A, Durazo R. 1997. A numerical study of the ex-ROFI of the Colorado River. J. Mar. Sys. 12: 17-33. [ Links ]
Cisneros-Mata MA, Montemayor-López G, Román-Rodríguez MJ. 1995. Life history and conservation of Totoaba macdonaldi. Conserv. Biol. 9: 806-814. [ Links ]
Dayton PK. 1998. Reversal of the burden of proof in fisheries management. Science 279: 821-822. [ Links ]
Dayton PK, Tegner MJ, Edwards PB, Riser KL. 1998. Sliding baselines, ghosts and reduced expectations in kelp forest communities. Ecol. Appl. 8: 309-322. [ Links ]
Dettman D, Flessa KW, Roopnarine PD, Schone BR, Goodwin DH. 2004. The use of oxygen isotope variation in shells of estuarine mollusks as a quantitative record of seasonal and annual Colorado River discharged. Geochim. Cosmochim. Acta 68: 1253-1263. [ Links ]
Dufour V, Pierre C, Rancher J. 1992. Stable isotopes in fish otoliths discriminate between lagoonal and oceanic residents of Taiaro Atoll (Tuamotu Archipelago French Polynesia). Coral Reefs 17: 23-28. [ Links ]
Flanagan CA, Hendrickson JR. 1976. Observations on the commercial fishery and reproductive biology of the totoaba, Cynoscion macdonaldi, in the northern Gulf of California. Fish. Bull. 74: 531-544. [ Links ]
Gilbert CH. 1891. Scientific results of the exploration by the US Fish Commission steamer Albatross. No. XII. A preliminary report on the fishes collected by the steamer Albatross on the Pacific coast of North America during the year 1889, with descriptions of twelve new genera and ninety-two new species. Proc. US Natl. Mus. 13: 49-126. [ Links ]
Goodfriend GA, Flessa KW. 1997. Radiocarbon reservoir ages in the Gulf of California: Roles of upwelling and flow from the Colorado River. Radiocarbon 39: 139-148. [ Links ]
Ingram BL, Conrad ME, Ingle JE. 1996. Stable isotope and salinity systematics in estuarine waters and carbonates: San Francisco Bay. Geochim. Cosmochim. Acta 60: 455-467. [ Links ]
Jackson JBC, Kirby MX, Berger WH, Bjorndal KA, Botsford LW, Bourque BJ, Bradbury RH, Cooke R, Erlandson J, Estes JA, Hughes TP, Kidwell S, Lange CB, Lenihan HS, Pandolfi JM, Peterson CH, Steneck RS, Tegner MJ, Warner RR. 2001. Historical overfishing and the recent collapse of coastal ecosystems. Science 293: 629-638. [ Links ]
Jamieson RE, Schwarcz HP, Brattey J. 2004. Carbon isotopic records from the otoliths of Atlantic cod (Gadus morhua) from eastern Newfoundland, Canada. Fish. Res. 68: 83-97. [ Links ]
Kalish JM. 1991. Oxygen and carbon stable isotopes in the otoliths of wild and laboratory-reared Australian salmon (Arripis trutta). Mar. Biol. 110: 37-47. [ Links ]
Lavín MF, Sánchez S. 1999. On how the Colorado River affected the hydrography of the upper Gulf of California. Cont. Shelf Res. 19: 1545-1560. [ Links ]
Lavín MF, Godínez VM, Alvarez LG. 1998. Inverse-estuarine features of the upper Gulf of California. Estuar. Coast. Shelf Sci. 47: 769-795. [ Links ]
Lercari D, Chávez E. 2007. Possible causes related to historic stock depletion of the totoaba, Totoaba macdonaldi (Perciformes: Sciaenidae), endemic to the Gulf of California. Fish. Res. 86: 136-142. [ Links ]
Patterson WP. 1998. North American continental seasonality during the last millennium: High resolution analysis of sagittal otoliths. Palaeogeogr., Palaeoclimatol., Palaeoecol. 138: 271-303. [ Links ]
Rodriguez CA, Flessa KW, Dettman DL. 2001. Effects of upstream diversion of Colorado River water on the estuarine bivalve molluscMulinia coloradoensis. Conserv. Biol. 15: 249-258. [ Links ]
Román-Rodríguez MJ, Hammann MF. 1997. Age and growth of totoaba, Totoaba macdonaldi (Sciaenidae), in the upper Gulf of California. Fish. Bull. 95: 620-628. [ Links ]
Rowell K. 2006. Isotopic logs of the Sea of Cortez: Oxygen and carbon stable isotopes in otoliths from marine fish record the impact of diverting the Colorado River from the sea. Dissertation, University of Arizona. [ Links ]
Rowell K, Flessa KW, Dettman DL, Román M. 2005. The importance of Colorado River flow to nursery habitats of the Gulf corvina (Cynoscion othonopterus). Can. J. Fish. Aquat. Sci. 62: 2874-2885. [ Links ]
Thorrold SR, Campana SE, Jones CM, Swart PK. 1997. Factors determining δ13C δ18O fractionation in aragonitic otoliths of marine fish. Geochim. Cosmochim. Acta 61: 2990-2919. [ Links ]
True CD, Loera AS, Castro NC. 1997. Acquisition of broodstock of Totoaba macdonaldi: Field handling, decompression, and prophylaxis of an endangered species. Prog. Fish-Cult. 59: 246-248. [ Links ]
True CD, Castro-Castro N, Sandoval-Garibaldi G, Morales-Ortiz C. 2001. Reproducción controlada de Totoaba macdonaldi (Gilbert). VIII Congreso Nacional y II Simpósium Internacional sobre el Mar de Cortés. [ Links ]
Weidman CR, Millner R. 2000. High-resolution stable isotope records from North Atlantic cod. Fish. Res. 46: 327-342. [ Links ]
Nota
Traducido al español por Christine Harris.

