










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCiencias marinas
versión impresa ISSN 0185-3880
Cienc. mar vol.33 no.3 Ensenada sep. 2007
Notas de investigación
Patrones de caza y éxito de captura de una población de águila pescadora (Pandion haliaetus) en Bahía Magdalena, Baja California Sur, México
Hunting patterns and success of an osprey (Pandion haliaetus) population at Magdalena Bay, Baja California Sur, Mexico
A Castellanos-Vera*, E Rivera
Centro de Investigaciones Biológicas del Noroeste, SC, Apartado postal No. 128, La Paz CP 23000, Baja California Sur, México, * E-mail: arcas04@cibnor.mx.
Recibido en marzo de 2007;
Aceptado en julio de 2007
Resumen
A finales del otoño de 2004 se estudió el efecto de la condición de la superficie del agua, la velocidad del viento y el estado de la marea en los patrones de caza o vuelo (aleteo, planeo y cernido) sobre la probabilidad de captura de la población de águila pescadora (Pandion haliaetus) de Bahía Magdalena, Baja California Sur, México. Mediante seguimientos focales se obtuvo el número de cacerías, el éxito de captura, el número de lanzamientos, la duración de la cacería y la proporción de tiempo de la cacería invertida en cada patrón de caza. El éxito de captura fue de 61%, con 2.4 lanzamientos en promedio para capturar una presa. La cacería duró, en promedio, 450 segundos, invirtiendo 50% de este tiempo en aleteo, 41% en planeo y 7% en vuelo cernido. El promedio de lanzamientos para capturar una presa en las diferentes condiciones de superficie del agua, velocidad del viento y de marea, varió significativamente. La probabilidad de captura de una presa disminuyó al incrementarse la velocidad del viento y al deteriorarse la condición de la superficie del agua; sin embargo, en ambos casos la duración de la cacería fue similar. Las águilas pescadoras en Bahía Magdalena disminuyen la proporción de aleteo y aumentan el planeo cuando la velocidad del viento se incrementa y la condición de la superficie del agua pasa de calmada a agitada. El cambio de la marea de bajamar a pleamar propició un incremento en la probabilidad de captura y en el número de lanzamientos por cacería, pero la duración de la cacería no tuvo cambios significativos. Nuestros resultados fueron comparados con estudios de otras regiones.
Palabras clave: Pandion haliaetus, águila pescadora, éxito de captura, patrones de caza, Baja California Sur.
Abstract
In late autumn 2004 we studied the effect of water surface condition, wind speed, and tidal status on the hunting patterns (flapping, gliding, hovering) and hunting success of the osprey population at Magdalena Bay, Baja California Sur, Mexico. Through focal sightings we obtained the number of hunting dives, hunting length, and the proportion of hunting length invested in each pattern. Hunting success was 61%. Ospreys required an average of 2.4 dives to catch a prey. Hunting lasted 450 seconds on average, with about 50% of this time invested in flapping, 41% in gliding, and 7% in hovering. The average number of dives to catch a prey varied according to the different water surface conditions, wind speed, and tidal status. The probability of catching a prey decreased when wind speed increased and the condition of the water surface deteriorated; however, hunting length was similar in both cases. Ospreys at Magdalena Bay decrease the proportion of flapping and increase gliding when wind speed increases and the condition of the water surface changes from calm to choppy. The change from low to high tide favored an increase in the probability of catching a prey and in the number of dives, but hunting length was not affected. We compared our results with studies from other regions.
Key words: Pandion haliaetus, osprey, hunting success, hunting patterns, Baja California Sur.
Introducción
La temperatura, la velocidad del viento, el estado de la marea, la intensidad luminosa y la lluvia, juegan un papel fundamental en el comportamiento y éxito de caza de las aves piscívoras (Lambert 1943, Dunn 1973, Ueoka y Koplin 1973, Grubb 1977, Stinson 1978, Szaro 1978, Swenson 1979, Russell y Burtt 1979, Stinson et al. 1987, Machmer e Ydenberg 1990, Phelps 1993). Sin embargo, existen discrepancias sobre la importancia de estas variables sobre el éxito de caza.
Por ejemplo, estudios realizados en poblaciones de águila pescadora (Pandion haliaetus), un depredador diurno que se alimenta exclusivamente de peces, indican que los patrones de caza y el éxito de captura se ven afectados significativamente debido a cambios en la velocidad del viento (Grubb 1977, Stinson 1978, Machmer e Ydenberg 1990). Por otro lado, Grubb (1977) y Machmer e Ydenberg (1990) coinciden en que el estado de la superficie del agua, influenciado por la velocidad del viento, influye en los patrones de caza y el éxito de captura. Además, Grubb (1977) y Stinson (1978) sugieren que la intensidad de la luz es otro factor que influye significativamente en el éxito de caza, ya que la visualización de las presas puede depender de la intensidad lumínica. Además de estas variables, Ueoka y Koplin (1973) sugieren que el estado de la marea puede afectar la duración de la cacería y el éxito de captura del águila pescadora, debido al comportamiento que presentan los peces por el movimiento de las mareas. En tanto, otros estudios sugieren que el éxito de captura se debe no a las condiciones ambientales, sino principalmente a la experiencia y habilidad que tienen los individuos para cazar, así como a las características de las presas (gobernadas por factores como el hábitat, localización y disponibilidad) (Szaro 1978, Swenson 1979).
Los estudios muestran divergencias de resultados en cuanto al efecto de las variables ambientales en los patrones de caza y éxito de captura del águila pescadora. Es posible que dichas diferencias se deban a la variabilidad geográfica de las poblaciones analizadas o a que la mayor parte de los estudios han sido realizados en poblaciones de águila pescadora migratorias, con diferentes requerimientos biológicos y ecológicos.
El águila pescadora, una especie de amplia distribución en la Península de Baja California, tiene poblaciones muy abundantes en las lagunas Ojo de Liebre-Guerrero Negro (Castellanos et al. 2001, Castellanos y Ortega-Rubio 1995) y San Ignacio (Danemann y Guzmán-Poo 1992). En Bahía Magdalena su población ha estado creciendo en años recientes, estimándose entre 80-100 parejas (A Castellanos, datos no publicados). Los nidos están distribuidos en las islas y en tierra firme, sobre postes de la red eléctrica en el poblado de Puerto Adolfo López Mateos, de la carretera y caminos de la zona (A Castellanos obs. pers.). Las águilas pescadoras se desplazan desde sus sitios de posadero (principalmente los nidos) hasta las aguas de la bahía para cazar, internándose en ella desde unos cuantos metros a varios kilómetros. Sin embargo, se desconoce la energía destinada a la captura de presas así como el efecto de las variables ambientales sobre el éxito de caza. Por lo anterior, el presente trabajo se enfocó en documentar el efecto de la velocidad del viento, el estado de la superficie del agua y de la marea, en los patrones de caza y el éxito de captura de una población residente de águila pescadora en Baja California Sur (BCS), México, a fin de ampliar y profundizar el conocimiento de la ecología de esta especie en regiones poco estudiadas, y en especial para generar información que permita entender las diferencias o similitudes entre las poblaciones residentes y las migratorias en cuanto a los patrones de caza y el éxito de captura.
Área de estudio y métodos
El estudio se realizó en Puerto Adolfo López Mateos y la playa Santa Elenita, ubicados en Bahía Magdalena, en la costa oeste de BCS, México, entre 24°15'-25°20' N y 111°30'-112°15' O. Bahía de Magdalena es uno de los humedales más importantes de la Península de Baja California, por su amplia variedad de especies y abundantes poblaciones de aves acuáticas residentes y migratorias (ver Massey y Palacios 1994, Carmona y Danemann 1998, Zárate-Ovando et al. 2006). Esta bahía se extiende a lo largo de 250 km de costa y su superficie abarca cerca de 1875 km2 de aguas profundas, canales, manglares y dunas; incluye además tres islas: Santo Domingo, Magdalena y Santa Margarita (Zárate-Ovando et al. 2006) (fig. 1).
Para analizar el efecto de tres variables ambientales (velocidad del viento, estado de la marea y de la superficie del agua) en los patrones de la caza y en el éxito de captura del águila pescadora, se realizaron observaciones en el campo durante 16 días, de octubre a diciembre de 2004 (etapa no reproductiva del águila pescadora). Las observaciones se hicieron diariamente en bloques de 2 h cada uno, entre las 09:00 y 17:00, invirtiendo un total de 96 h, desde dos puntos sobre la línea de costa (Santa Elenita y López Mateos) separados 7 km entre sí, que permitían un campo de visión de aproximadamente 2 km2 de superficie del mar, es decir, bahía adentro donde se alimentaban las águilas pescadoras. Desde estos puntos, mediante seguimientos focales (seguimiento de un individuo con la ayuda de binoculares y de un telescopio) de las águilas pescadoras que ingresaban en el campo de visión del observador, se documentó la secuencia y duración de los episodios de aleteo, planeo y vuelo cernido, así como el número de lanzamientos por cacería (suma de lanzamientos exitosos y fallidos). Se denominó lanzamiento exitoso cuando el ave capturó una presa, y lanzamiento fallido aquel en que el ave tuviera o no contacto con el agua no capturó presa alguna. Dado que las aves no estaban marcadas, para minimizar el seguimiento de los mismos individuos se utilizaron dos sitios de observación dentro de la misma micro-región con condiciones ambientales similares.
El trabajo de campo se desarrolló por tres personas. Una persona se encargada de cronometrar el tiempo empleado en cada uno de los patrones de caza. La segunda persona realizaba el seguimiento focal, el cual daba inicio cuando algún águila pescadora entraba en el campo de visión del observador y terminaba cuando el ave obtenía una presa, se posaba o salía del campo de visión (Ver Machmer e Ydenberg 1990). La tercera persona anotaba los datos dictados por los otros dos observadores y registraba el estado de las superficie del agua, la altura de la marea y la velocidad del viento cada 20 minutos.
El estado de la superficie del agua se clasificó en calmada, rizada o agitada. La altura de la marea se determinó utilizando el calendario de mareas del Centro de Investigación Científica y Educación Superior de Ensenada (CICES; González et al. 2004). La altura de la marea se clasificó como: pleamar cuando el agua estaba >150 cm por encima del nivel de referencia, media cuando el agua estaba <150 y >50 cm encima del nivel de referencia, y bajamar cuando el agua estaba <50 cm encima del nivel de referencia. La velocidad del viento se determinó con un anemómetro digital y se clasificó en ocho intervalos: 0 (0-0.9 m s-1), 1 (1-1.9 m s-1), 2 (2-2.9 m s-1), 3 (3-3.9 m s-1), 4 (4-4.9 m s-1), 5 (5-5.9 m s-1), 6 (6-6.9 m s-1) y 7 (> 7 m s-1).
Para el análisis de datos sólo se emplearon los seguimientos focales en los que el ave realizó al menos un lanzamiento, y a los cuales se denominó cacerías (C). Se excluyeron los casos en que se detectó un águila pescadora en el campo visual del observador que no efectuó ningún lanzamiento. De las cacerías, se obtuvo el número de cacerías exitosas (CE) (cacería en la que el ave capturó un pez), la duración en segundos de cada cacería (TC) y el número de lanzamientos por cacería.
La proporción de tiempo de la cacería invertida en aleteo (PA), planeo (PP) y vuelo cernido (PS), se calculó dividiendo el tiempo invertido en cada uno de los patrones de caza entre la duración de cada cacería (PA = TIA/TC, PP = TIP/TC, PS = TIS/TC). Asimismo, para cada variable ambiental se calculó la probabilidad de captura (P) mediante la formula P = CE/C. El éxito de captura para la población se calculó dividiendo el número total de cacerías exitosas entre el número total de cacerías (ECP = CET/CT). De las cacerías exitosas se obtuvo el promedio de lanzamientos por cacería necesarios para capturar una presa.
El promedio de lanzamientos por cacería, las proporciones de tiempo invertidas en cada patrón de caza, así como la duración total de la cacería (tiempo de cacería transformado a logaritmo de base 10), se compararon con las condiciones ambientales mediante análisis de varianza de una vía. Dado que los datos de patrones de caza constituyen proporciones de tiempo, se hizo una transformación al arco seno de sus valores para normalizarlos. Asimismo se utilizó el análisis de varianza de una vía para determinar la asociación entre la probabilidad de captura y las condiciones de la superficie del agua, los estados de la marea y la velocidad del viento. La velocidad del viento y el estado de la superficie del mar mostraron una correlación alta (r = 0.83, n = 142, P < 0.001), por ello, no se efectuó un análisis de correlación múltiple de ambas variables con las patrones de caza y el éxito de captura, ya que tal análisis no proporcionaría mayor explicación que la obtenida únicamente con la velocidad del viento.
Resultados
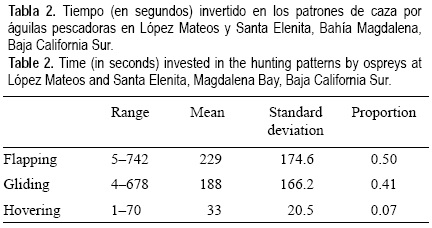
Se observaron un total de 142 cacerías, de las cuales 87 fueron exitosas y 55 no exitosas, lo que significa un éxito de captura de 61%. En promedio cada ave utilizó 2.4 lanzamientos para capturar una presa; el máximo de lanzamientos fue 7 y el mínimo 1 (tabla 1). Los individuos de águila pescadora invirtieron en promedio 450 s durante la cacería, con un mínimo de 25 s y un máximo de 919 s, de los cuales 50% fueron de aleto, 41% de planeo y 7% de vuelo cernido (tabla 2).
El promedio de lanzamientos requeridos para capturar un pez varió significativamente entre las diversas condiciones de la superficie del agua (F2,139 = 17.506, P < 0.05) (tabla 3). La probabilidad de captura de una presa fue significativamente más alta con la superficie del mar calmada que cuando ésta estaba rizada o agitada (F2,139 = 17.42, P < 0.0001) (tabla 3).
La duración de la cacería fue similar en las tres condiciones de la superficie del agua (F2 139 = 0.098, P > 0.05) (tabla 3). No obstante se detectaron diferencias significativas en la proporción de tiempo que el ave emplea en los patrones de caza, disminuyendo el aleteo (F2,139 = 18.54, P < 0.05) (tabla 3) e incrementándose el planeo (F2,139= 11.97495, P < 0.05) y el vuelo cernido (F2,139= 3.095592, P < 0.05) cuando la condición del agua cambió de calmada a agitada.
El promedio de lanzamientos que se requirieron para obtener una presa se incrementó de 1.9 a 3.5 (F7,134 = 5.385144, P < 0.05) (tabla 4) cuando la velocidad del viento pasó de 0 a 7m s-1. La probabilidad de captura fue relativamente alta y estable con vientos entre 0 y 3 m s-1; sin embargo, cuando la velocidad del viento sobrepasó los 4 m s-1, la probabilidad de captura decreció significativamente (F7,134 = 5.1288, P < 0.0001) (tabla 4).
La duración total de la cacería se incrementó con la velocidad del viento y al deteriorarse la condición de la superficie del agua; sin embargo las diferencias no son estadísticamente significativas (F7,134= 0.81254, P > 0.05) (tabla 4). Los patrones de caza variaron con la velocidad del viento; las águilas pescadoras aletearon en mayor proporción cuando la velocidad del viento fue baja (de 0 a 3 m s-1), disminuyendo cuando ésta superó los 4 m s-1 (F2,139 = 18.27013, P <0.0001) en tanto que el planeo aumentó significativamente (F2,139 = 11.92003, P < 0.001) y el vuelo cernido ligeramente (F2,139 = 3.1185, P < 0.05) (tabla 4).
El número promedio de lanzamientos por cacería varió en cada uno de los estados de la marea, alcanzando su valor más alto en marea baja (F2,139 = 8.446882, P < 0.001). La probabilidad de captura se incrementó (0.53, 0.54 y 0.73) cuando la marea cambió de bajamar a pleamar (F2,139 = 2.7523, P = 0.034) (tabla 5). Cuando el estado de la marea cambió de bajamar a pleamar, la duración de la cacería disminuyó, sin embargo la diferencia no fue estadísticamente significativa (F2,139 = 2.017, P > 0.05) (tabla 5).
Discusión
El éxito de captura de la población de águila pescadora en Bahía Magdalena (61%) fue menor que el reportado para las poblaciones migratorias localizadas en la costa occidental de Canadá (89.34%; Lambert 1943), en el Lago Cascade, Idaho, EUA (69%; Phelps 1993) y en los manglares del sureste de Brasil (71%; Silva y Olmos 2002). No obstante, resultó más eficiente que la población de Lago George, Florida (24%; Grubb 1977).
Nosotros encontramos que el número de lanzamientos requeridos por los individuos para capturar a una presa, fue mayor que el de las poblaciones localizadas en los EUA y occidente de Canadá, aunque, la duración de la cacería resultó menor (ver Ueoka y Koplin 1973, Szaro 1978, Swenson 1978, Phelps 1993). Esto sugiere que la población de águila pescadora de Bahía Magdalena fue más eficiente ya que el éxito de captura resultó alto, en una cacería relativamente corta. Estas diferencias de eficiencia pueden deberse a factores como la disponibilidad de presas y de su comportamiento alimenticio (Swenson 1979) o a la experiencia y edad de las águilas pescadoras (Szaro 1978), entre otras razones.
Nuestros resultados muestran que cuando la condición de la superficie del agua cambia de calmada a agitada, la proporción de aleteo disminuye significativamente y la de planeo se incrementa, en tanto que la de vuelo cernido, permanece estable. Las águilas pescadoras ajustan sus patrones de caza con la velocidad del viento; así, en Bahía Magdalena el aleteo disminuye y el planeo aumenta cuando la velocidad del viento se incrementa. Estos hallazgos son consistentes con los realizados en poblaciones del occidente de Canadá (Machmer e Ydenberg 1990). Sin embargo, la proporción de tiempo que el ave invierte en el vuelo cernido en Bahía Magdalena, es comparativamente mayor que la registrada por Machmer e Ydenberg (1990).
La variabilidad del tiempo empleado en cada patrón de caza, al ser éstos actividades excluyentes, puede ser una forma en que el ave compensa el gasto de energía (Machmer e Ydenberg 1990), de acuerdo a las condiciones ambientales prevalecientes. De esta manera, las águilas aprovechan las corrientes ascendentes de aire caliente generadas por convección de la atmósfera, o el choque de las corrientes de aire con los riscos o montañas, ya que el aleteo es el patrón más costoso en energía pues requiere de gran esfuerzo de los músculos pectorales (Welty y Baptista 1990, Navarro y Benítez 1995). Dado que en Bahía Magdalena las águilas pescadoras requirieron en promedio de un mayor número de lanzamientos para cada cacería, la proporción de vuelo cernido, es mayor, pues cada lanzamiento implica una fracción de este tipo de vuelo.
El promedio de lanzamientos requeridos para capturar un pez se incrementa cuando la superficie del agua cambia de calmada a agitada. Asimismo, observamos que la probabilidad de captura disminuye, mientras que la duración de la cacería aumenta al deteriorarse la condición de la superficie del agua de calmada a agitada y al incrementarse la velocidad del viento. Estos hallazgos son consistentes con los de estudios de poblaciones más norteñas (ver Grubb 1977, Machmer e Ydenberg 1990). Es posible que tanto el número de lanzamientos requeridos para capturar una presa como la probabilidad de captura respondan (el primero aumenta y la segunda disminuye) a la reducción de la visualización que tiene el ave sobre su presa, dado que el viento agita el agua e incrementa su turbidez (ver Grubb 1977, Machmer e Ydenberg 1990). En Bahía Magdalena el número de lanzamientos es casi el doble de lo reportado para la población de águila pescadora del occidente de Canadá. Esto puede deberse a que durante el período de estudio, el patrón de viento diario se caracterizó por el incremento de la velocidad conforme avanzaba el día y por un viento intenso (>4 m s-1) durante 66% del día, y en consecuencia, de una superficie del agua agitada. En el occidente de Canadá se registraron vientos muy intensos (>7 m s-1) durante 5% del día (Machmer e Ydenberg 1990).
Un estudio realizado en Florida sugiere que el éxito de captura del águila pescadora se incrementa cuando disminuye la profundidad del agua por la marea baja, ya que los peces presentan mayor vulnerabilidad (Szaro 1978). En este trabajo, sin embargo, el número de lanzamientos fue menor y la probabilidad de captura se mantuvo alta durante la pleamar. Estas divergencias pueden deberse a las diferentes condiciones físicas y geográficas entre los sitios de estudio, que a su vez pueden influir en el comportamiento del depredador y de la presa, ya que la migración vertical y horizontal de los peces puede estar determinada por diferentes condiciones locales como el desplazamiento de las mareas (Maes et al. 1999). Además, en nuestra área de estudio la marea baja dejaba aguas muy someras, lo cual podría reflejarse en menos presas disponibles y, por ello, en una probabilidad de captura relativamente baja en comparación a la observada en otras latitudes.
Los vientos intensos y aguas agitadas que prevalecen en más de la mitad del día en Bahía Magdalena, parecen ser relativamente adversos para las águilas pescadoras (depredador diurno, piscívoro) ya que pueden implicar un mayor gasto energético para atrapar una presa (la probabilidad de captura disminuye y el número de lanzamientos se incrementa con la velocidad del viento). Tales condiciones son posiblemente compensadas en cierta medida por la elevada incidencia (37%) de cacerías exitosas en pleamar (que ocurrieron principalmente en las mañanas), cuando el viento fue menos intenso que en el resto del día y en consecuencia la probabilidad de captura fue mayor. Esto explica en parte las altas ocurrencias de captura en pleamar, una diferencia con respecto a otras poblaciones estudiadas. Todos estos ajustes en los patrones de caza y de captura parecen responder a la necesidad de las águilas pescadoras de lograr la captura de una presa al menor costo energético posible en las condiciones ecológicas de Bahía Magdalena.
Agradecimientos
Agradecemos al Centro de Investigaciones Biológicas de Noroeste SC y al Proyecto 2002-C01-0844 SEMARNAT-CONACYT de A Ortega-Rubio el apoyo financiero para realizar este trabajo; a A Cozar y P Támez su ayuda en el trabajo de campo, a C Arguelles su valioso y excelente apoyo técnico, y a J Rivera la edición de la figura. Asimismo agradecemos a H de la Cueva y a dos revisores anónimos sus críticas y sugerencias académicas al manuscrito.
Rererencias
Carmona R, Danemann GD. 1998. Distribución espaciotemporal de aves en la salina de Guerrero Negro, Baja California Sur, México. Cienc. Mar. 24: 3 89-408. [ Links ]
Castellanos A, Ortega-Rubio A. 1995. Artificial nesting sites and ospreys at Ojo de Liebre and Guerrero Negro lagoons, Baja California Sur, Mexico. J. Field Ornithol. 66: 117-127. [ Links ]
Castellanos A, Salinas F, Ortega-Rubio A. 2001. Inventory and conservation of breeding waterbirds at Ojo de Liebre and Guerrero Negro lagoons, Baja California Sur, Mexico. Cienc. Mar. 27: 351-373. [ Links ]
Danemann GD, Guzmán-Poo J. 1992. Notes on the birds of San Ignacio Lagoon, Baja California Sur, Mexico. West. Birds 23: 11-19. [ Links ]
Dunn EK. 1973. Changes in fishing ability of terns associated with wind speed and sea surface conditions. Nature 244: 520-521. [ Links ]
González JI, Soto R, Ochoa J. 2004. Predicción de mareas en México. Oceanografía Física, CICESE. Fecha de última modificación 10 de diciembre de 2003. Fecha de acceso 20 de septiembre de 2004. http://oceanografía.cicese.mx/predmar/predicción. [ Links ]
Grubb TC. 1977. Weather-dependent foraging in ospreys. Auk 94: 146-149. [ Links ]
Lambert GG. 1943. Predation efficiency of osprey. Can. Field-Nat. 57: 87-88. [ Links ]
Machmer MM, Ydenberg CR. 1990. Weather and osprey foraging energetics. Can. J. Zool. 68: 40-13. [ Links ]
Maes J, Pas J, Taillieu A, Van Damme PA, Ollevier F. 1999. Diet changes in the vertical distribution of juvenile fish in the Zeeschelde estuary. J. Fish Biol. 54: 1329-1333. [ Links ]
Massey WB, Palacios E. 1994. Avifauna of the wetlands of Baja California, Mexico: Current status. Stud. Avian Biol. 15: 45-57. [ Links ]
Navarro A, Benítez H. 1995. El Dominio en el Aire. Fondo de Cultura Económica, México DF, 211 pp. [ Links ]
Phelps JM. 1993. Factors affecting foraging and productivity of ospreys (Pandion haliaetus) at Cascade Reservoir, Idaho. Thesis, Boise State University, USA, 69 pp. [ Links ]
Russell BR, Burtt EH. 1979. Weather-dependent foraging of great blue herons (Ardea herodias). Auk 96: 628-630. [ Links ]
Silva SS, Olmos F. 2002. Osprey ecology in the mangroves of southeastern Brazil. J. Raptor Res. 36: 328-331 [ Links ]
Stinson CH. 1978. The influence of environmental conditions on aspects of the time budgets of breeding ospreys. Oecologia 36: 127-139. [ Links ]
Stinson CH, Lauthner J, Ray RT. 1987. The effect of weather conditions on the behavior of ospreys in northwestern Washington. Can. J. Zool. 65: 2116-2118. [ Links ]
Swenson JE. 1978. Prey and foraging behavior of ospreys on Yellowstone Lake, Wyoming. J. Wildl. Manage. 42: 87-90. [ Links ]
Swenson JE. 1979. The relationship between prey species ecology and dive success in ospreys. Auk 96: 408-412. [ Links ]
Szaro CR. 1978. Reproductive success and foraging behavior of the osprey at Seahorse Key, Florida. Wilson Bull. 90: 112-118. [ Links ]
Ueoka LM, Koplin RJ. 1973. Foraging behavior of ospreys in northwestern California. Raptor Res. 7: 32-38. [ Links ]
Welty JC, Baptista L. 1988. The Life of Birds. 4th ed. Saunders College Publ., New York, 581 pp. [ Links ]
Zárate-Ovando B, Palacios E, Reyes-Bonilla H, Amador E, Saad G. 2006. Waterbirds of the Lagoon Complex Magdalena Bay-Almejas, Baja California Sur, Mexico. Waterbirds 29: 350-364 [ Links ]

