










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkCiencias marinas
versão impressa ISSN 0185-3880
Cienc. mar vol.33 no.3 Ensenada Set. 2007
Nota de investigación
La "extinción de Stilostomella" durante el Pleistoceno medio frente a la costa norte de Chile (25°S)
Mid-Pleistocene "Stilostomella extinction event" off northern Chile (25°S)
S Núñez-Ricardo1,2, M Marchant3, B Hayward4, D Hebbeln5
1 Graduate Program in Oceanography, Department of Oceanography, University of Concepción, Casilla 160-C, Concepción, Chile. E-mail: snunez@udec.cl.
2 Center for Oceanographic Research in the eastern South Pacific, Chile (COPAS).
3 Department of Zoology, University of Concepción, P.O. Box 160-C, Concepción, Chile.
4 GeoMarine Research, 49 Swainston Rd, St Johns, Auckland, New Zealand.
5 Research Center Ocean Margins, Bremen University, D-28334 Bremen, Germany.
Recibido en diciembre de 2006;
Aceptado en mayo de 2007
Resumen
Usando muestras de sedimentos del testigo GeoB 3388-1 de la dorsal de Iquique, norte de Chile (25°S), reportamos la última fecha de aparición (UFA) de siete especies del grupo extinto Stilostomella. La extinción de Stilostomella, el más grande recambio de foraminíferos bentónicos de aguas profundas, ha sido documentada en trabajos previos para el Pacífico sudoriental (PSO) y es ampliada por nuestro trabajo como evidencia cualitativa de este evento. La extinción de Stilostomella involucra la desaparición de especies elongadas de las familias Nodosariidae, Pleurostomellidae y Stilostomellidae. La UFA de Myllostomella sp. Hayward 2002 (518 cm, 0.79 ma) del testigo GeoB 3388-1 es similar a un registro previo para el PSO, pero es anterior a reportes a nivel mundial (0.65-0.57 ma), por lo que se concluye que el momento de extinción en el PSO fue diferente al ocurrido en los océanos Atlántico y Pacífico sudoccidental. La UFA de Myllostomella sp. coincide con el incremento en δ13C y CaCO3, sugiriendo cambios en la disponibilidad de nutrientes y la circulación del PSO durante la Transición del Pleistoceno Medio. El mecanismo causal preciso de la extinción del grupo Stilostomella (enfriamiento, incremento en la oxigenación de las aguas de fondo, cambios en el suministro de alimento) permanece sin resolverse.
Palabras clave: grupo Stilostomella, Pleistoceno, foraminíferos de aguas profundas, Pacífico sudoriental, Chile.
Abstract
Based on sediment samples from core site GeoB 3388-1 from the Iquique Ridge off northern Chile (25°S), we report the last appearance datum (LAD) of seven species that belong to the Stilostomella extinction group. The Stilostomella extinction was the last major turnover in deep-sea benthic foraminifera. It is documented in previous works for the southeastern Pacific (SEP) and is extended here by our results, giving further qualitative indication of this important extinction event. The Stilostomella extinction involves the disappearance of elongate species presenting cribrate (Chrysalogonium), slit lunate, hooded with two teeth (Pleurostomellidae), or secondarily toothed and necked (Stilostomellidae) apertures. The LAD of Myllostomella sp. Hayward 2002 (518 cm, 0.79 Ma) in core GeoB 3388-1 is remarkably consistent with a previous record for the SEP, but is prior to other previous worldwide reports (0.65-0.57 myr); thus, we conclude that the timing of the SEP extinction was different from that in the Atlantic Ocean and southwestern Pacific. Additionally, the LAD of Myllostomella sp. corresponds to an increase in both benthic δ13C and CaCO3 content, suggesting that a profound change in nutrient availability and circulation occurred in the SEP during the Mid-Pleistocene Transition. The precise mechanistic cause of the Stilostomella extinction (cooling, increased oxygenation of bottom waters, changes in food supply) is yet to be resolved.
Key words: Stilostomella group, Pleistocene, deep-sea benthic foraminifera, southeastern Pacific, Chile.
Introducción
Transición del Pleistoceno Medio
El cambio en la variabilidad climática durante el Cuaternario medio de un modo dominado por oblicuidad a uno dominado por excentricidad, fechado alrededor de 0.9 ma y conocido como la Revolución del Pleistoceno Medio (Berger y Jansen 1994) o la Transición del Pleistoceno Medio (TPM, Raymo et al. 1997), representa uno de los grandes enigmas de la investigación Cuaternaria (e.g., Ruddiman 2003). Se caracteriza por una prolongación e intensificación marcada de ciclos climáticos glaciales e interglaciales entre 900 y 650 ka. A pesar del cambio pronunciado en la respuesta del sistema de la Tierra evidenciado en los registros paleoclimáticos, las características de la frecuencia y amplitud de los parámetros orbitales que causan cambios climáticos globales a largo plazo, tales como la excentricidad (~100 ka), la oblicuidad (~41 ka) y la precesión (~21 y ~19 ka), no varían durante la TPM (Berger et al. 2005). Esto sugiere que la causa del cambio durante la TPM es interno más que externo al sistema climático global, y muestra el papel de los "feedbacks" (o mecanismos de retroalimentación) en los ciclos glaciales e interglaciales en la amplificación del forzamiento débil de la insolación (Imbrie et al. 1993). No obstante, aún no se entiende completamente ni existe una buena explicación del mecanismo de este cambio climático durante la TPM (ver revisión de Ruddiman 2003).
Extinción de foraminíferos bentónicos durante el Pleistoceno medio
La extinción durante la TPM de foraminíferos bentónicos de aguas profundas con conchas elongadas, cilindricas, uni-seriales y multiseriales, y con aberturas con carácteristicas muy específicas (e.g., pequeñas y redondas, dentadas, cribadas o en forma de media luna), tales como Orthomorphina, Pleurostomella, Siphonodosaria y Stilostomella, se conoce como la "extinción de Stilostomella" (por la desaparición de la familia Stilostomellidae durante ese tiempo; Weinholz y Lutze 1989) y hasta recientemente ha recibido poca atención científica. Este evento de extinción fue identificado por primera vez en el Océano Atlántico, donde se registró la desaparición de diez especies de foraminíferos bentónicos de seis géneros cerca de la inversión magnética Brunhes/Matuyama (B/M) entre 1.0 y 0.6 ma (Lutze 1979, Caralp 1985, Thomas 1987, Weinholz y Lutze 1989, Schönfeld 1996). Las extinciones sucedieron de 0.1 a 0.2 ma antes, a profundidades mayores a 3000 m y en latitudes meridionales (Weinholz y Lutze 1989, Schönfeld 1996). La extinción de Stilostomella es un evento estratigráfico de utilidad global cerca de la frontera B/M: e.g., en el Océano Índico (Gupta 1993, Kawagata et al. 2006), el Atlántico Norte (Caralp 1985, Kawagata et al. 2005), el Mar Caribe (Hayward et al. 2006), el Atlántico Sur (O'Neill et al. en prensa), el Pacífico noroccidental (Keller 1980, Kaiho 1992, Jian et al. 2000, Hess y Kuhnt 2005, Kawagata et al. en prensa), el Pacífico Sur central (Schönfeld 1995), el Pacífico sudoriental (Schönfeld y Spiegler 1995) y el Pacífico sudoccidental (Hayward 2001, 2002). Estos estudios también documentaron una disminución significativa en la abundancia de los géneros elongados, Pleurostomella y varios nodosáridos a partir del Pleistoceno temprano en el Pacífico Sur central (Schönfeld 1995) y el Atlántico (Caralp 1985, Thomas 1987). Schönfeld (1996) realizó una revisión de la extinción de Stilostomella durante el Pleistoceno medio con base en registros globales e indicó que la desaparición de los especímenes de mayor tamaño (>150 μm) ocurrió entre 1.0 y 0.6 ma (predominantemente de 0.8 a 0.7 ma) con un ritmo muy variable. Más recientemente, varios estudios (Hayward 2001, 2002; Hayward y Kawagata 2005; Hayward et al. 2006; Kawagata et al. 2005, 2006, 2007; O'Neill et al. en prensa) han mostrado que la transición climática del Plioceno tardío al Pleistoceno medio (3.0-0.55 ma) resultó ser un intervalo de mucho recambio con la extinción de 96 especies y 19 géneros (>63 μm) de foraminíferos bentónicos elongados y cilíndricos (de las familias Stilostomellidae, Pleurostomellidae y Nodosariidae). La mayoría (76 especies) desaparecieron durante la TPM (1.20.55 ma), con el tiempo de extinción final de este grupo normalmente entre 0.7 y 0.55 ma. Aunque no se extinguieron todos los nodosáridos, la mayoría de los taxones elongados uniseriales en aguas profundas desaparecieron o decrecieron considerablemente en abundancia (e.g., Hayward 2002).
Aún no se entiende bien el significado paleoceanográfico de la extinción de Stilostomella, aunque se ha especulado que se relaciona con el enfriamiento glacial total y una mayor ventilación de las aguas profundas, o con cambios rápidos en el suministro de alimentos. El objetivo de este trabajo fue documentar, con mayor detalle, las extinciones de foraminíferos bentónicos de aguas profundas en el Pacífico sudoriental usando un testigo fechado de la dorsal de Iquique, frente a la costa norte de Chile (~25°S), debajo del Agua Profunda del Pacífico (APP), un área donde se han realizado procos estudios sobre la escala de tiempo de la evolución Cuaternaria.
Material y métodos
Sitio de estudio
El testigo de sedimento GeoB 3388-1 fue obtenido bajo las aguas superficiales de la Corriente Perú-Chile. Esta corriente, la Corriente de Frontera Oriental más larga, se deriva de la bifurcación de la Corriente Circumpolar Antártica entre 40°S y 45°S, y se mueve hacia el norte a lo largo de la costa de Sudamérica hasta unirse a ~5°S con la Corriente Ecuatorial del Sur que fluye hacia el oeste (fig. 1a). El Océano Pacífico sudoriental tiene una rica estructura de masas de agua, con aguas que se originan en el Pacífico ecuatorial y subtropical al norte y en el Océano Sur al sur. El Agua Superficial Subtropical de alta salinidad se forma al sur de ~5°S y al norte del Frente Subtropical, donde la evaporación excede la precipitación (Stramma et al. 1995). Una capa somera de salinidad mínima aparece por debajo del Agua Superficial Subtropical (e.g., Reid 1973) y por arriba del Agua Ecuatorial Subsuperficial, de alta salinidad y bajo contenido de oxígeno, de la Corriente Submarina Gunther que fluye hacia el sur entre 100 y 400 m de profundidad sobre la plataforma y el talud continental frente a la costa central y norte de Chile (Toggweiler et al. 1991). Otra masa de agua más profunda es el Agua Intermedia Antártica, caracterizada por baja salinidad y alto contenido de oxígeno, la cual fluye hacia el ecuador entre profundidades de 400 y 1200 m. Por debajo de ésta prevalece el Agua Profunda del Pacífico, que fluye lentamente hacia el sur (fig. 1b). Una buena parte de la exportación de fosfatos y nitratos en el Pacífico ocurre en esta última masa de agua (Wunsch et al. 1983).
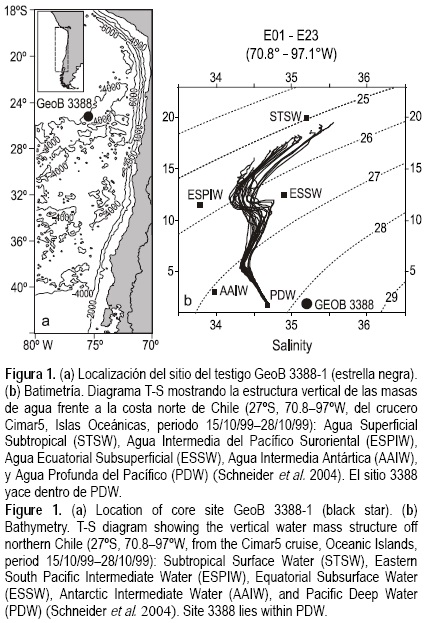
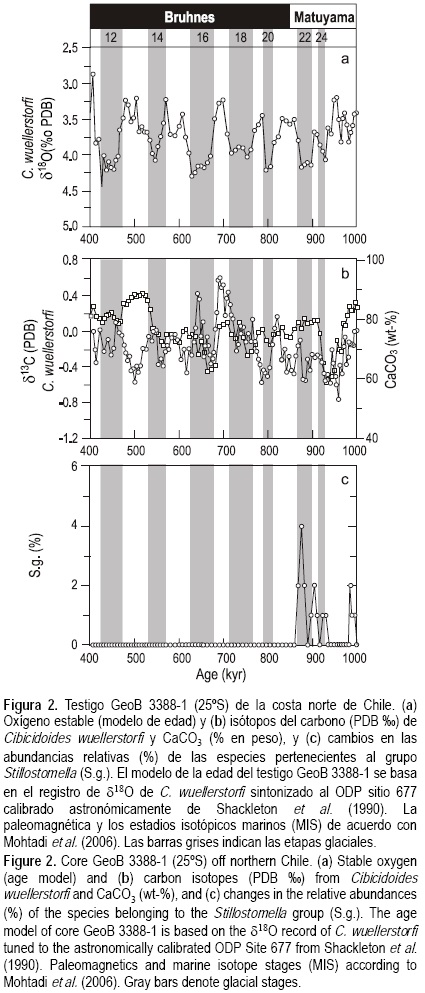
El testigo GeoB 3388-1 fue recolectado de la dorsal de Iquique (25°13.2' S, 75°31.5' W, 3558 m de profundidad del agua, 722 cm de largo) durante el crucero CHIPAL (SO-102) a bordo del B/O Sonne en 1995 (Hebbeln et al. 1995, fig. 1a). Dos muestras de 10 cm3 fueron tomadas cada 5 cmy liofilizadas. Se realizaron conteos de todos los foraminíferos bentónicos elongados (>150 μm) en las muestras compactadas de sedimento abarcando el último millón de años, enfocándose aquí sólo en el periodo de 1.0 a 0.5 ma AP. Se midió la composición de los isótopos de carbono y del oxígeno estable de las conchas del foraminífero bentónico Cibicidoides wuellerstorfi con un espectrómetro de masas Finnigan MAT 251 (datos tomados de Mohtadi et al. 2006). Para cada medición se seleccionaron 20 conchas individuales (>212 μm). El modelo de edad del testigo GeoB 3388-1 está basado en el registro de los isótopos de oxígeno estable de C. wuellerstorfi sintonizado al sitio 677 del Programa de Perforación en el Océano (ODP por sus siglas en inglés) (Shackleton et al. 1990), el cual es sustentado por análisis paleomagnéticos (Mohtadi et al. 2006) (fig. 2a). Con base en el modelo de edad, se calcularon las tasas de sedimentación (TS) lineales para cada edad a partir de la diferencia: TS = D2 - D1/A2 - A1, donde D1/A1 y D2/A2 son la menor/mayor profundidad/edad (cm ka-1), respectivamente. Finalmente, para determinar el contenido de carbono orgánico total (COT), 25 mg del material de muestra triturado fueron descalcificados con HCl 6 N, secados sobre una hornilla a 80°C y medidos en un analizador elemental Heraeus-CHN de acuerdo con Müller et al. (1994). El contenido de carbono total (CT) se obtuvo de muestras no tratadas. El contenido de carbonato fue calculado a partir de la diferencia entre los contenidos de CT y COT: CaCO3 = (TC - TOC) × 8.33 (fig. 2b).
Resultados
En el sitio GeoB 3388-1, por lo menos siete especies desaparecieron durante la TMP (grupo Stilostomella; figs. 2c, 3), incluyendo un miembro de la familia Pleurostomellidae, el foraminífero biserial Pleurostomella pleurostomella (Silvestri 1904) (678 cm, 1.0 ma), y todos los miembros de la familia Stilostomellidae: las formas elongadas uniseriales de Myllostomella sp. Hayward 2002 (518 cm, 0.795 ma), Myllostomella costai (Schwager 1866) (588 cm, 0.91 ma), Myllostomella hyugaensis (Ishizaki 1943) (598 cm, 0.92 ma), Siphonodosaria sagrinensis (Bagg 1912) (558 y 583 cm, 0.87 y 0.9 ma), y Siphonodosaria spinea (Cushman 1939) (588 cm, 0.87 ma).
El modelo de edad del testigo GeoB 3388-1 se basó en el registro de isótopos de oxígeno estable del foraminífero bentónico C. wuellerstorfi, el cual es sustentado por análisis paleo-magnéticos (Mohtadi et al. 2006). El testigo GeoB 3388-1 se remonta a -1.03 ma AP (fig. 2a). Por tanto, se pueden observar valores de δ18O más pesados/ligeros durante los ciclos glaciales/interglaciales (intervalos de 3.4 a 44/2.9 a 4.2, respectivamente), los cuales están relacionados con un mayor/ menor volumen de hielo. Las tasas de sedimentación variaron entre 0.3 y 1.2 cm ka-1, con una media de 0.75 cm ka-1, correspondiendo a una resolución de muestra promedio de 6-7 ka. En general, los valores de δ13C más ligeros de C. wuellerstorfi (δ13C) caracterizan los sedimentos depositados entre 0.97 y 0.60 ma (i.e., estadios isotópicos marinos (MIS por sus siglas en inglés) 21-15), seguidos por un incremento notable durante el MIS 13 interglacial, simultáneamente con un incremento en el contenido de CaCO3 (fig. 2b). A escala glacial/ interglacial, en general se obervan valores de δ13C más ligeros/ pesados (intervalos de -0.8 a 0.2/3.4 a 4.4, respectivamente), lo que indica un mayor/menor contenido de nutrientes, respectivamente.
Discusión
Los foraminíferos bentónicos generalmente tienen tasas de recambio faunal relativamente lentas según la estabilidad relativa del ambiente de aguas profundas (McKinney 1987), con varios episodios de mayor intercambio, el último de los cuales fue durante la TPM. En el testigo de este estudio, se relacionó este último episodio de mayor extinción global con la última fecha de aparición (UFA) de siete especies pertenecientes al grupo Stilostomella: Myllostomella sp., M.costai, M. hyugaensis, Siphonodosaria sagrinensis, S. spinea y Pleurostomella pleurostomella. Estas especies fueron documentadas en trabajos previos (Hayward 2001, 2002; Hayward et al. 2006; Kawagata et al. 2006, y respectivas referencias), lo que sugiere que sus extinciones fueron eventos mundiales.
La desaparición de las especies del grupo Stilostomella ha sido registrada a nivel mundial. La UFA menor (0.8 ma) del testigo GeoB 3388-1 está dentro del intervalo ya documentado (1.0 a 0.6 ma; Weinholz y Lutze 1989; Schónfeld 1996; Hayward 2001, 2002; Hayward et al. 2006). De manera interesante, en otro reporte se indica que la extinción de Stilostomella en el Pacífico sudoriental (ODP sitio 861, 45°51' S, 75°41' W, 1652 m de profundidad del agua) sucedió más temprano (1.3 ma) que en otras partes de los océanos mundiales (1 a 0.6 ma) (Gavriloff 2006) y también cae dentro del intervalo documentado para la extinción de Stilostomella.
Entre las causas de los resultados de nuestro reporte anterior sobre la UFA del grupo Stilostomella se encuentran: la fracción del tamaño utilizada (>150 μm en vez de >63 μm, Hayward 2001) y los cambios en las propiedades de la masa de agua (en realidad APP) que bañó nuestro testigo GeoB 3388-1 (3.5 km de profundidad).
En otros trabajos se ha resaltado la importancia de los cambios en las masas de agua con relación a la extinción del grupo Stilostomella. Kawagata et al. (2005) relacionaron la desaparición más temprana en el sitio 758 del Océano Índico (2925 m), en comparación con el sitio más somero 722 (2045 m), con cambios oceanográficos ocurridos antes en masas de agua profundas (e.g., agua de fondo de origen Antártico) que en masas de agua intermedias (e.g., Agua Intermedia Glacial del Atlántico Norte). Asimismo, Hayward (2001) sugirió que la desaparición de las formas elongadas uniseriales está relacionada con masas de agua más frías y probablemente más oxigenadas, lo cual indica cambios en la circulación del agua profunda. Los cambios en las masas de agua afectan el suministro y la calidad del alimento de los foraminíforos bentónicos. Se ha sugerido que las especies que pertenecen al grupo Stilostomella estaban bien adaptadas a condiciones de alto flujo de carbono/bajo contenido de oxígeno. Con base en nuestros datos, se puede observar que a escala glacial/interglacial existen cambios en los valores de δ13C que se han asociado con cambios en el reservorio de carbono (Mohtadi et al. 2006), probablemente el contenido de nutrientes (con valores más ligeros/pesados indicando más/menos nutrientes). No obstante, de 1 a 0.5 ma el contenido medio de carbono orgánico en el sedimento del testigo GeoB 3388-1 es constante y bajo (<0.5%), lo que sugiere un desfase entre la señal de δ13C como marcador de nutrientes y la preservación del carbono orgánico en nuestros sedimentos. El periodo de la extinción mundial del grupo Stilostomella se caracteriza por un bajo contenido de CaCO3 en el sitio de nuestro testigo en comparación con otros valores documentados, especialmente del MIS 15 al MIS 13 (fig. 2b). Se han asociado los cambios en la preservación de carbonatos durante la TPM con los cambios en la intensidad de la formación del agua profunda del Atlántico Norte, así como con el crecimiento de sábanas de hielo en los últimos 0.9 ma (MIS 22) (Schmieder et al. 2000). Estos resultados confirman el acontecimiento de cambios importantes en los flujos de carbono y las condiciones químicas de los carbonates frente a la costa norte de Chile durante la TPM. Como el carbono orgánico de nuestro testigo GeoB 3388-1 no cambia a lo largo del periodo y con base en los datos disponibles, relacionamos las causas de la extinción discutidas más con el oxígeno que con la disponibilidad de alimento.
Agradecimientos
Este trabajo fue apoyado por los programas de becas DAAD y MCESUP/UCO-0002, el Centro de Investigación Oceanográfica en el Pacífico Suroriental (FONDAP-COPAS Proyecto No. 150100007), la Comisión Nacional Chilena de Ciencia y Tecnología (FONDECYT No. 1040968), y la Secretaría Federal de Educación e Investigación de Alemania (BMBF) a través de financiamiento del proyecto CHIPAL y el crucero a bordo del B/O Sonne (SO-102). Agradecemos al capitan y la tripulación del B/O Sonne sus esfuerzos y apoyo durante el crucero, así como a M Segl y B Meyer-Schack su apoyo en las mediciones de los isótopos estables.
Traducido al español por Christine Harris.
Referencias
Bagg RM Jr. 1912. Pliocene and Pleistocene foraminifera from Southern California. Bull. US Geol. Surv. 513: 1-153. [ Links ]
Berger A, Mélice JL, Loutre MF. 2005. On the origin of the 100-kyr cycles in the astronomical forcing. Paleoceanography 20: doi:10.1029/2005PA001173. [ Links ]
Berger WH, Jansen E. 1994. Mid-Pleistocene shift: The Nansen connection. Geophys. Monogr. 84: 295-311. [ Links ]
Caralp MH. 1985. Quaternary calcareous benthic foraminifers, Leg 80. In: De Graciansky PC et al. (eds.), Initial Reports of the Deep Sea Drilling Project. US Government Printing Office, Washington DC, pp. 725-755. [ Links ]
Cushman JA. 1939. Eocene foraminifera from submarine cores off the eastern coast of North America. Contrib. Cushman Lab. Foraminifer. Res. 15: 49-76. [ Links ]
Gavriloff I. 2006. The mid-Pleistocene Stilostomella extinction event in the southeast Pacific Ocean: A review. Anu. Inst. Geocien., UFRJ ISSN 0101-9759, 29 : 490-491. [ Links ]
Gupta AK. 1993. Biostratigraphic vs. paleoceaonographic importance of Stilostomella lepidula (Schwager) in the Indian Ocean. Micropaleontology 39: 47-52. [ Links ]
Hayward BW. 2001. Global deep-sea extinctions during the Pleistocene ice-ages. Geology 29: 599-602. [ Links ]
Hayward BW. 2002. Late Pliocene to middle Pleistocene extinctions of deep-sea benthic foraminifera ("Stilostomella extinction") in the Southwest Pacific. J. Foraminifer. Res. 32: 274-306. [ Links ]
Hayward BW, Kawagata S. 2005. Extinct foraminifera figured in Brady's Challenger Report. J. Micropalaeontol. 24: 171-175. [ Links ]
Hayward BW, Kawagata S, Grenfell HR, Droxler AW, Shearer, M. 2006. Mid-Pleistocene extinction of bathyal benthic foraminifera in the Caribbean Sea. Micropaleontology 52: 53-73. [ Links ]
Hebbeln D. et al. 1995. Report and preliminary results of SONNE Cruise SO 102, Valparaiso-Valparaiso, 95. pp. 1-134. [ Links ]
Hess S, Kuhnt W. 2005. Neogene and Quaternary paleoceanographic changes in the southern South China Sea (Site 1143): The benthic foraminiferal record. Mar. Micropaleontol. 54: 63-87. [ Links ]
Imbrie J, Berger A, Boyle EA, Clemens SC, Duffy A, Howard WR, Kukla G, Kutzbach J, Martinson DG, McIntyre A, Mix AC, Molfino B, Morley JJ, Peterson LC, Pisias NG, Prell WL, Raymo ME, Shackleton NJ, Toggweiler JR. 1993. On the structure and origin of major glaciation cycles. 2. The 100,000-year cycle. Paleoceanography 8: 699-735. [ Links ]
Ishizaki K. 1943. On the species of Ellipsonodosaria from Japan. Trans. Nat. Hist. Soc. Taiwan 33: 678-689. [ Links ]
Jian Z, Wang P, Chen M-P, Zhao Q, Buhring C, Laj C, Lin H-L, Pflaumann U, Bian Y, Wang R, Cheng X. 2000. Foraminiferal response to major Pleistocene paleoceanographic changes in the southern China Sea. Paleoceanography 15: 229-243. [ Links ]
Kaiho K. 1992. Eocene to Quaternary benthic foraminifers and paleobathymetry of the Izu-Bonin Arc, Legs 125 and 126. In: Taylor B et al. (eds.), Proceedings of the Ocean Drilling Program, Scientific Results. College Station, Texas, pp. 285-310. [ Links ]
Kawagata S, Hayward BW, Grenfell, HR, Sabaa AT. 2005. Mid-Pleistocene extinction of deep-sea foraminifera in the North Atlantic Gateway (ODP Sites 980 and 982). Palaeogeogr. Palaeoclimatol. Palaeoecol. 221: 267-291. [ Links ]
Kawagata S, Hayward BW, Gupta AK. 2006. Benthic foraminiferal extinctions linked to late Pliocene-Pleistocene deep-sea circulation changes in the northern Indian Ocean (ODP Sites 722 and 758). Mar. Micropaleontol. 58: 219-242. [ Links ]
Kawagata S, Hayward BW, Kuhnt W. Extinction of deep-sea foraminifera as a result of Pliocene-Pleistocene deep-sea circulation changes in the South China Sea (ODP Sites 1143 and 1146). Quaternary. Sci. Rev. 26: 808-827. [ Links ]
Keller G. 1980. Benthic foraminifers and paleobathymetry of the Japan Trench area, Leg 57, Deep Sea Drilling Project. In: Von Huene R et al. (eds.), Initial Reports of the Deep Sea Drilling Project. US Government Printing Office, Washington DC, pp. 835-865. [ Links ]
Lutze GF. 1979. Benthic foraminifera at Site 397: Fluctuations and ranges in the Quaternary. In: Von Rad U et al. (eds.), Initial Reports of the Ocean Drilling Program. Vol. 47. US Government Printing Office, Washington DC, pp. 419- 431. [ Links ]
McKinney ML. 1987. Taxonomic selectivity and continuous variation in mass and background extinctions of marine taxa. Nature 325: 143-145. [ Links ]
Mohtadi M, Hebbeln D, Nünez-Ricardo S, Lange CB. 2006. El Ninolike pattern in the Pacific during marine isotope stages (MIS) 13 and 11. Paleoceanography 21: PA1015, doi:10.1029/2005PA001190. [ Links ]
Müller PJ, Schneider R, Ruhland G. 1994. Late Quaternary PCO2 variations in the Angola Current: Evidence from organic carbon δ13C and alkenone temperature. In: Zahn R, Pedersen TF, Kaminski MA, Labeyrie L. (eds.), Carbon Cycling in the Glacial Ocean: Constraints on the Ocean's Role in Global Change. NATO ASI Ser., Springer-Verlag, New York, Vol. I, pp. 343-366. [ Links ]
O'Neill TA, Hayward BW, Kawagata S, Sabaa AT, Grenfell HR. Pleistocene extinctions of deep-sea foraminifera: The South Atlantic record. J. Palaeontology (in press). [ Links ]
Palmer DK, Bermüdez PJ. 1936. An Oligocene foraminiferal fauna from Cuba. Mem. Soc. Cubana Hist. Nat. Felipe Poey 10: 227-271. [ Links ]
Raymo ME, Oppo DW, Curry W. 1997. The mid-Pleistocene climate transition: A deep sea carbon isotopic perspective. Paleoceanography 12: 546-559. [ Links ]
Reid JL. 1973. The shallow salinity minima of the Pacific Ocean. Deep-Sea Res. 20: 51-68. [ Links ]
Ruddiman WF. 2003. Orbital insolation, ice volume, and greenhouse gases. Quatern. Sci. Rev. 22: 1597-1629. [ Links ]
Schmieder F, von Dobeneck T, Bleil U. 2000. The mid-Pleistocene climate transition as documented in the deep South Atlantic Ocean: Initiation, interim state and terminal event. Earth Planet. Sci. Lett. 179: 539-549. [ Links ]
Schneider W, Fuenzalida R, Garcés J. 2004. Corrientes marinas y masas de agua. In: Werlinger C (eds.), Biología Marina y Oceanografía: Conceptos y Procesos. Vol. 1. Universidad de Concepción, Chile, pp. 177-194. [ Links ]
Schönfeld J. 1995. Biostratigraphy and assemblage composition of benthic foraminifera from the Manihiki Plateau, southwestern tropical Pacific. J. Micropalaeontol. 14: 165-175. [ Links ]
Schönfeld J. 1996. The "Stilostomella Extinction": Structure and dynamics of the last turn-over in deep-sea benthic foraminiferal assemblages. In: Moguielvsky EA, Whatley R (eds.), Microfossils and Oceanic Environments. Aberystwyth Press, Wales, pp. 27-37. [ Links ]
Schönfeld J, Spiegler D. 1995. Benthic foraminiferal biostratigraphy of Site 861, Chile Triple Junction, Southeastern Pacific. In: Lewis SD et al. (eds.), Proceedings of the Ocean Drilling Program: Scientific Results. Vol. 141. College Station, Texas, pp. 213- 224. [ Links ]
Schwager C. 1866. Fossile Foraminiferen von Kar Nicobar: Novara Expeditions. Geol. Theil. 2: 187-268. [ Links ]
Shackleton NJ, Berger A, Peltier WR. 1990. An alternative astronomical calibration of the lower Pleistocene timescale based on ODP Site 677. Trans. R. Soc. Edinb. Earth Sci. 81: 251-261. [ Links ]
Silvestri O. 1904. Forme nuove o poco conosciute di Protozoi miocenici piemontesi. Atti Accad. Sci. Torino (1903-1904) 39: 415. [ Links ]
Stramma L, Peterson RG, Tomczak M. 1995. The South Pacific Current. J. Phys. Oceanogr. 25: 77- 91. [ Links ]
Thomas E. 1987. Late Oligocene to Recent deep-sea benthic foraminifera from the central equatorial Pacific Ocean. In: Ruddiman WF et al. (eds.), Initial Reports of the Deep Sea Drilling Project. Vol. 94. US Government Printing Office, Washington DC, pp. 997-1032. [ Links ]
Toggweiler JR, Dixon K, Broecker WS. 1991. The Peru upwelling and the ventilation of the South Pacific thermocline. J. Geophys. Res. 20: 467-497. [ Links ]
Weinholz P, Lutze GF. 1989. The Stilostomella extinction. In: Ruddiman WF et al. (eds.), Proceedings of the Ocean Drilling Program: Scientific Results. College Station, Texas, pp. 113-117. [ Links ]
Wunsch C, Hu DX, Grant B. 1983. Mass, heat, salt, and nutrient fluxes in the South Pacific Ocean. J. Phys. Oceanogr. 13: 725-753. [ Links ]

