










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkCiencias marinas
versão impressa ISSN 0185-3880
Cienc. mar vol.32 no.4 Ensenada Dez. 2006
Notas de Investigación
Ciclo estacional de una pradera marina de Cymodocea nodosa y la ictiofauna asociada en Playa Dorada(Lanzarote, Islas Canarias, Atlántico oriental)
Seasonal cycle of a Cymodocea nodosa seagrass meadow and of the associated ichthyofauna at Playa Dorada (Lanzarote, Canary Islands, eastern Atlantic)
F Tuya, JA Martín, A Luque*
Departamento de Biología, Universidad de Las Palmas de Gran Canaria, 35017 Campus de Tafira s/n, Las Palmas de Gran Canaria, Islas Canarias. * E-mail: aluque@dbio.ulpgc.es
Recibido en junio de 2005
Aceptado en agosto de 2006.
Resumen
Se estudió el ciclo estacional de una pradera mixta de Cymodocea nodosa y Caulerpa prolifera, así como el de la ictiofauna asociada, desde marzo de 2001 hasta abril de 2002, en Playa Dorada (Lanzarote, Islas Canarias, Atlántico oriental). Algunas variables estudiadas presentaron un claro comportamiento estacional (bimodal), con máximo en primavera-verano (abril-septiembre) y mínimos en invierno (diciembre-marzo). Así, la densidad de haces osciló entre 372 y 1014 haces m-2, la biomasa de C. nodosa entre 94 y 271 g PS m-2, la biomasa vegetal total entre 94 y 383 g PS m-2, y la longitud de hoja entre 10 y 18 cm. La riqueza media ictiológica fluctuó entre 0.7 y 5.3 especies por censo, y la abundancia total de individuos entre 2.0 y 144.0 ind 100 m-2. Aparece una clara correlación entre el ciclo anual de C. nodosa y los descriptores del poblamiento ictiológico de la pradera.
Palabras clave: praderas marinas, fanerógamas marinas, comunidades de peces, estacionalidad, Islas Canarias.
Abstract
The seasonal cycle of a mixed seagrass meadow of Cymodocea nodosa and Caulerpa prolifera and of the associated ichthyofauna was studied at Playa Dorada (Lanzarote, Canary Islands, eastern Atlantic) from March 2001 to April 2002. A seasonal (bimodal) cycle was clearly observed, with maximum values in spring and summer (April-September) and minimum in winter (December-March). Shoot densities oscillated between 372 and 1014 shoots m-2, the biomass of Cymodocea nodosa between 94 and 271 g DW m-2, total vegetal biomass between 94 and 383 g DW m-2 and mean leaf lengths between 10 and 18 cm. Mean fish richness fluctuated between 0.7 and 5.3 species and total fish densities between 2.0 and 144.0 ind 100 m-2. There was a clear correlation between the cycles of C. nodosa and the associated fish population.
Key words: seagrass meadows, marine phanerogams, fish assemblages, seasonality, Canary Islands.
Introducción
Las praderas marinas de Cymodocea nodosa (Ucria) Ascherson se distribuyen por todo el Mediterráneo y la costa norte de África, siendo la fanerógama marina más abundante en los alrededores de las Islas Canarias (Afonso-Camllo y Gil-Rodríguez 1980, Reyes 1993, Reyes et al. 1995, Barberà et al. 2005). Esta planta se localiza en Canarias en fondos sedimentarios entre 2-30 m de profundidad, principalmente al sur y sureste de las islas, frecuentemente formando praderas mixtas con individuos del género Caulerpa particularmente Caulerpa prolifera (Forsskal) Lamouroux a poca profundidad (<10 m) (Pavón-Salas et al. 2000).
Cymodocea nodosa está declarada como especie vulnerable dentro del catálogo de especies amenazadas de Canarias (BOCAC 2001) y existen varios Lugares de Interés Comunitario (LIC) creados por su posible presencia sobre bancos de arena poco profundos (DOCE 2002).
Los estudios realizados sobre la dinámica de las praderas de C. nodosa en el Mediterráneo (Pérez y Camp 1986, Peduzzi y Vukovic 1990, Terrados y Ros 1992, Mazella et al. 1993, Vidondo et al. 1997) han mostrado una clara estacionalidad anual de esta planta, al igual que lo mostró el estudio desarrollado por Reyes et al. (1995) en el sur de Tenerife. Sin embargo, ninguna investigación ha estudiado su estacionalidad en la isla de Lanzarote, que se encuadra en un contexto biogeográfico diferente al de Tenerife y al del Mediterráneo. Al ser la isla más oriental del Archipiélago Canario, por su cercanía al continente africano sus aguas pueden verse afectadas de forma más importante por las fluctuaciones, tanto térmicas como en disponibilidad de recursos nutritivos, asociadas al cercano afloramiento sahariano.
Si bien se ha observado una notable estacionalidad en las comunidades de macrofauna invertebrada asociadas a tres praderas marinas de C. nodosa de Canarias (Tuya et al. 2001), ningún estudio ha analizado la posible relación entre el ciclo anual de C. nodosa y las comunidades de peces asociadas a dichas praderas.
Consecuentemente, los objetivos de este estudio fueron: estudiar las variaciones anuales en densidad, biomasa y longitud de hoja de una pradera marina mixta de C. nodosa y Caulerpa prolifera en Playa Dorada (sur de Lanzarote, Islas Canarias, Atlántico oriental) y estudiar la posible estacionalidad del poblamiento íctico asociado a dicha pradera, a efectos de verificar si existe relación entre el ciclo anual de biomasa de C. nodosa y el poblamiento íctico asociado.
Material y métodos
Lugar de estudio
La pradera marina mixta de Cymodocea nodosa y Caulerpa prolifera se localiza frente a Playa Dorada (municipio de Yaiza, Lanzarote; 28°51.5' N, 15°49.4' W) entre los 4 y 8 m de profundidad, sobre un fondo de arena biogénica. El estudio se desarrolló de marzo de 2001 a abril de 2002, con toma de muestras mensual, excepto en los meses de agosto y noviembre de 2001 y enero y marzo de 2002 (diez muestreos en total). A lo largo del estudio se midieron los dos parámetros abióticos que más influyen en las fluctuaciones estacionales de las praderas marinas de C. nodosa (Sand-Jensen 1975, Pérez y Camp 1986), la temperatura del agua de mar en superficie, mediante sonda YSI©, y la densidad de flujo fotónico en superficie en el espectro del visible (PAR), mediante el radiómetro Li-Cor 4π©.
Muestreo de los descriptores de la pradera
En cada muestreo se recolectaron aleatoriamente, a 5 m de profundidad, nueve muestras (n = 9) de pradera mixta de C. nodosa y C. prolifera mediante cajas de plástico de 15 χ 26 cm con ayuda de dos palas y equipos autónomos de buceo. Las muestras se conservaban en nevera hasta su posterior análisis en el laboratorio. Después de lavarse, se contaron el número de haces y se midió la longitud de treinta hojas de cada muestra seleccionadas aleatoriamente, siguiendo las indicaciones de Phillips y McRoy (1980). Estas medidas permitieron la estimación de dos de los cuatro descriptores de la pradera, (1) la densidad de haces y (2) la longitud media de hoja para cada periodo de muestreo. Posteriormente en el laboratorio se obtuvo (3) la biomasa en peso seco (g PS) de C. nodosa y (4) la biomasa total (C. nodosa + C. prolifera), tras secar las muestras 24 h a 100°C (Sand-Jensen 1975, Pérez y Camp 1986). Todos los datos obtenidos se expresaron utilizando el m2 como unidad de superficie.
Ictiofauna
Se realizaron tres censos visuales (n = 3) mediante transec-tos de 25 χ 4 m (100 m2 de observación cada censo) en cada uno de los diez muestreos, siguiendo procedimientos estandarizados (Lincoln-Smith 1988, Kingsford y Battershill 1998, García-Charton et al. 2000). Para cada muestreo se obtuvieron como descriptores de la comunidad íctica: (1) la riqueza media de especies, (2) la riqueza total de especies ícticas observadas y (3) la abundancia total de individuos visualizados. Estos descriptores han mostrado ser eficaces para caracterizar las comunidades ícticas litorales en el Archipiélago Canario (Bortone et al. 1991, Falcón et al. 1996).
Análisis estadístico
Para cada uno de los descriptores de la pradera y de la comunidad íctica observada se ejecutó un ANOVA de un factor, correspondiente al factor "periodo de muestreo". Se empleó el contraste a posteriori SNK para detectar diferencias entre los distintos niveles de dicho factor. Detectamos una fuerte heterogeneidad de varianzas que no pudo evitarse con la transformación de los datos. Por ello, y con objeto de hacer más conservador el análisis, fijamos α en 0.01 en vez de en 0.05 (Underwood 1997).
Ejecutamos un análisis de correlación (coeficiente de Pearson) entre el ciclo anual de la biomasa de la planta Cymodocea nodosa y los descriptores de la comunidad ictiológica analizada. Todos los análisis los realizamos con el programa de cómputo SPSS 10.0.3©.
Resultados
Descriptores de la pradera
Para los cuatro parámetros estudiados se observó un patrón bimodal de variación temporal, con máximos en los meses correspondientes a primavera-verano (de abril a septiembre) y mínimos en los meses de invierno (diciembre a marzo). Para la densidad de haces (fig. 1) se obtuvo un promedio máximo de 1014 haces m-2, en abril de 2001, que se mantuvo con ligeras oscilaciones hasta septiembre y descendió paulatinamente hasta un promedio mínimo de 372 haces m-2 en diciembre de 2001. La biomasa de C. nodosa muestra un comportamiento similar (fig. 2) con un promedio máximo de 271 g PS m-2 en septiembre de 2001, que bajó hasta un mínimo de 94 g PS m-2 en febrero de 2002. Para la biomasa vegetal total (fig. 3) detectamos el máximo en abril de 2001 (383 g PS m-2) y el mínimo en diciembre de 2001 (94 g PS m-2). Finalmente, para la longitud media de hoja (fig. 4) obtuvimos un promedio máximo de 18 cm en septiembre de 2001 y un mínimo de 10 en febrero de 2002.
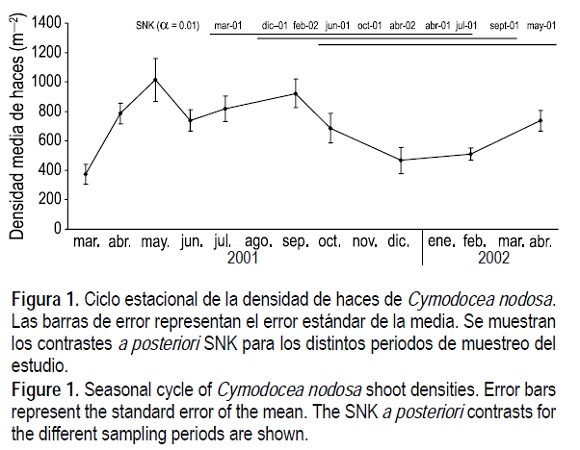


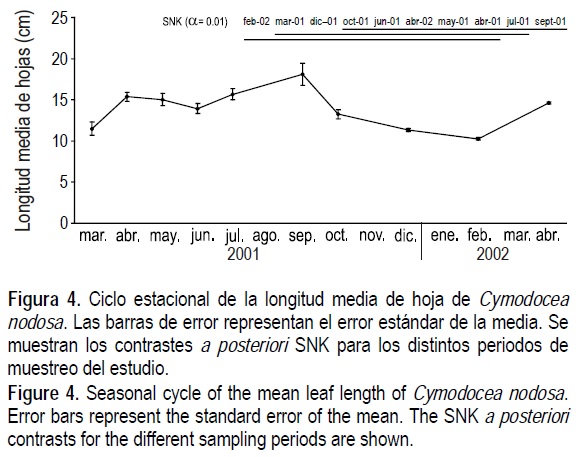
Para los cuatro parámetros se detectaron diferencias significativas (P < 0.01) debido al factor "periodo de muestreo". En cada una de las figuras se muestran los contrastes a posteriori SNK que indican las diferencias significativas observadas entre periodos.
Ictiofauna
La tabla 1 muestra la evolución del poblamiento íctico a lo largo del estudio. Se observó un total de 13 especies. Los espáridos fueron la familia más abundante con un total de cuatro especies observadas. Las especies más frecuentes (% frecuencia > 50%) fueron Xyrichthys novacula (90%) y Sphoeroides marmoratus (80%).
Al igual que observamos con C. nodosa, este estudio mostró una clara estacionalidad en los estadísticos descriptores de la comunidad de peces, apareciendo diferencias significativas (P < 0.01) debido al factor "periodo de muestreo", tanto para la riqueza media de especies ícticas como para la abundancia total media de peces. Se observó un valor máximo de riqueza media por censo (fig. 5) en septiembre de 2001, equivalente a 5.3 especies 100 m-2. El mínimo se observó en marzo de 2001 (0.7 especies 100 m-2). El mayor valor de riqueza total de especies observadas se obtuvo en septiembre de 2001, y fue de un total de 8 especies (fig. 6). El mínimo obtenido fue de una única especie tanto en diciembre de 2001 como en febrero de 2002. La abundancia total media de individuos (fig. 7) no mostró un patrón de estacionalidad uniforme debido a la presencia de un cardumen denso de bogas (Boop boops) en abril de 2001, que produjo la estimación de una densidad media de 144.0 ind 100 m-2. El mínimo de abundancia total fue de 2 ind 100 m-2 en diciembre de 2001. En cada una de estas figuras se muestran los contrastes a posteriori SNK que indican las diferencias significativas observadas.
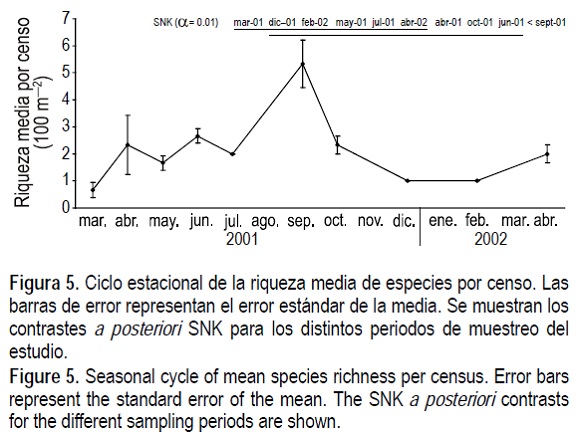


Correlación entre los ciclos de la planta C. nodosa y los descriptores del poblamiento íctico
Se obtuvo una correlación significativa y positiva entre el ciclo de biomasa media mensual de C. nodosa y el ciclo anual de la riqueza media de especies ícticas observadas (n = 10, rs = 0.753, P < 0.05). Sin embargo, no se detectó una correlación significativa con el ciclo anual de la abundancia media total de individuos observados (n = 10, rs = 0.582, P > 0.05). La falta de correlación se debe a la presencia de un valor anormalmente alto en abril de 2001 que, como se comentó anteriormente, está motivado por la visualización de un cardumen denso de bogas (B. boops). Al eliminar este dato (abril de 2001) obtuvimos un valor significativo de correlación entre ambos ciclos (n = 9, rs = 0.703, P < 0.05).
Parámetros abióticos
La temperatura del agua de mar osciló entre un máximo de 23.1°C (septiembre y octubre) y un mínimo de 18.2°C en marzo (fig. 8). La densidad de flujo fotónico varió entre un valor medio de 2650 μmol m-2 s-1 en julio y septiembre y 1550 μmol m-2 s-1 en marzo (fig. 8).

Discusión
La estacionalidad de la planta C. nodosa es un hecho bien conocido en el Mediterráneo (Pérez y Camp 1986, Peduzzi y Vukovic 1990, Terrados y Ros 1992, Mazella et al. 1993,Vidondo et al. 1997), así como en Canarias (Reyes et al. 1995), de manera que estas fluctuaciones están ligadas a variaciones en factores externos (Buia et al. 2000). Nuestro estudio corroboró estas observaciones. Sin embargo, es preciso señalar que las fluctuaciones estacionales de estos parámetros son inferiores a las detectadas en algunas zonas del Mediterráneo posiblemente en respuesta al menor gradiente anual de temperatura de agua de mar en Lanzarote (18.2-23.1°C) en comparación con el Mediterráneo (8-25°C, Pérez y Camp 1986). El ciclo de producción de la planta muestra un comportamiento bimodal, con un periodo de alta producción en primavera-verano y otro de baja producción en otoño-invierno. En la zona de estudio la biomasa de C. nodosa fluctuó aproximadamente entre 100 y 275 g PS m-2, mientras que en algunas áreas del Mediterráneo se han señalado fluctuaciones mucho mayores (175-631 g PS m-2 en el delta del Ebro, Pérez y Camp 1986; 323-1020 g PS m-2 en el golfo de Trieste, Peduzzi y Vukovic 1990). En cualquier caso, el rango de valores obtenidos en la presente investigación son inferiores a los mostrados por estos autores en el Mediterráneo y Reyes et al. (1995) en Tenerife. Asimismo, la densidad de haces detectada en el presente estudio (400-1000 haces m-2) es inferior a la observada por Pérez (1989, 1000-2000 haces m-2), Terrados y Ros (1992, 1000-1900 haces m-2), Mazella et al. (1993, 925-1925 haces m-2) y Reyes et al. (1995, 1000-2000 haces m-2).
Es preciso señalar que la ausencia de diferencias significativas entre muchos de los periodos de muestreo se debe a (1) el empleo de un nivel de significancia α = 0.01 y (2) a la gran variabilidad en los descriptores analizados, tanto para el poblamiento íctico como para la propia pradera. Esto se demuestra con los elevados valores de error estándar obtenidos para todos ellos, que reflejan la gran variabilidad de los procesos naturales asociados a los mismos.
También se ha observado una clara tendencia estacional (bimodal) en el poblamiento íctico de la pradera. Si bien la detección de patrones estacionales en comunidades de peces litorales es un hecho conocido en substratos rocoso-arenosos (García-Rubies 1997, Valle 2005) y praderas marinas en el Mediterráneo (Francour 1997), su detección es un hecho poco común en Canarias (Falcón et al. 1996). La ausencia de patrones estacionales en comunidades en peces litorales en Canarias responde a dos factores: (1) el escaso gradiente térmico anual, en comparación con otras zonas templado-cálidas como el Mediterráneo, y (2) a la sobrexplotación de los recursos demersales, que puede llegar a enmascarar a los propios procesos naturales (productividad, reclutamiento, etc.) (Falcón et al. 1996). La clara estacionalidad observada se debe a que las praderas marinas son emplazamientos donde el calado de nasas, que representa el 70% del esfuerzo pesquero en el litoral del Archipiélago (Hernández-García et al. 1998), no se desarrolla de forma intensiva debido a su escasa profundidad, y a que no albergan poblaciones de gran interés comercial (p.e. serránidos) que desarrollan sus ciclos de vida sobre fondos duros. También es debida a que una notable parte de los individuos visualizados son reclutas de talla pequeña que escapan de las artes de pesca. Estos reclutas encuentran alimento y refugio en las praderas de C. nodosa de Canarias, donde desarrollan sus primeros estadíos de vida (Brito 1991).
Nuestro estudio mostró una clara correlación entre el ciclo anual de biomasa de la fanerógama C. nodosa y el poblamiento íctico de la pradera. Este solapamiento está motivado, principalmente, por que el reclutamiento de ciertas especies de peces (p.e. Sparisoma cretense, Symphodus (Crenilabrus) mediterraneus, Diplodus annularis, Mullus surmuletus) durante primavera-verano, coincidiendo con los periodos de máximo crecimiento de la planta. Este mismo comportamiento fue obtenido por Guidetti y Bussotti (1997) para los Espáridos Diplodus annularis y Spondyliosoma cantharus en praderas marinas del Mediterráneo, mediante técnicas de censos visuales como las empleadas en el presente estudio. Posteriormente y estudiando el mismo entorno, Guidetti y Bussotti (2000) observaron que la riqueza, diversidad y densidad de especies ícticas demersales alcanzaban sus máximos entre primavera e inicio de otoño, si bien cada especie mostró un patrón particular. Asimismo y como mostraron Tuya et al. (2001) en tres praderas de C. nodosa en Gran Canaria, es a lo largo de la primavera-verano cuando se observan los valores más altos de riqueza, diversidad y abundancia de comunidades de macrofauna invertebrada asociadas a dichas praderas. Esta misma tendencia ha sido observada por diferentes autores en distintas partes del mundo (Ansari et al. 1991, Ansari y Parulekar 1994, Conolly y Butler 1996). Consecuentemente, el patrón estacional de esta planta regula las fluctuaciones estacionales de los descriptores de los poblamientos de macrofauna asociados a las mismas, tanto para vertebrados (peces) como para invertebrados. Este comportamiento podría deberse a que el de C. nodosa es un sistema de menor complejidad estructural que las praderas constituidas por otras fanerógamas (p.e. Posidonia oceanica), de manera que las comunidades de macrofauna están más ligadas a la variabilidad estacional de C. nodosa, al ser los sistemas de esta planta menos estables, donde no se desarrolla una compleja red de interacciones entre las diferentes comunidades existentes (Buia et al. 2000).
Agradecimientos
Nuestras gracias a J Pérez y AL Luque-Sollheim por su ayuda en los muestreos y a M Pérez por permitirnos disponer de su embarcación.
Referencias
Afonso-Carrillo J, Gil-Rodríguez MC. 1980. Cymodocea nodosa (Ucria) Ascherson y las praderas submarinas o "sebadales" en el Archipiélago Canario. Vieraea 8: 365-376. [ Links ]
Ansari ZA, Parulekar AH. 1994. Meiobenthos in the sediments of seagrass meadows of Lakshadweep atolls, Arabian Sea. Vie Milieu 44: 185-190. [ Links ]
Ansari ZA, Rivonker CU, Ramani P, Parulekar AH. 1991. Seagrass habitat complexity and macroinvertebrate abundance in Lakshadweep coral reef lagoons, Arabian Sea. Coral Reefs 10: 127-131. [ Links ]
Barberá C, Tuya F, Boyra A, Sánchez-Jérez P, Blanch I, Haroun RJ. 2005. Spatial variation in the structural parameters of Cymodocea nodosa seagrass meadows in the Canary Islands. Bot. Mar. 48: 122-126. [ Links ]
BOCAC. 2001. Boletín Oficial de la Comunidad Autónoma de Canarias Decreto 151/2001 de 23 de julio, por el que se crea el Catálogo de Especies Amenazadas de Canarias. [ Links ]
Bortone SA, Van Tasell J, Brito A, Falcón JM, Bundrick CM. 1991. A visual assessment of the inshore fishes and fishery resources off El Hierro, Canary Islands: A baseline survey. Sci. Mar. 55: 529-541. [ Links ]
Brito A. 1991. Catálogo de los Peces de las Islas Canarias. Francisco Lemus Editor, La Laguna, 230 pp. [ Links ]
Buia MC, Gambi MC, Zupo V. 2000. Structure and functioning of Mediterranean seagrass ecosystems: An overview. Biol. Mar. Mediterr. 7: 167-190. [ Links ]
Conolly RM, Butler AJ. 1996. The effects of altering seagrass canopy height on small, motile invertebrates of shallow Mediterranean embayments. PSZNI Mar. Ecol. 17: 637-652. [ Links ]
DOCE. 2002. Diario Oficial de las Comunidades Europeas de 9.1.2002 (L5/16) por el que se se crean los espacios de interés comunitario de la Macaronesia, 92/43/CEE del Consejo de Europa. [ Links ]
Falcón JM, Bortone SA, Brito A, Bundrick CM. 1996. Structure and relationships within and between the littoral rock-substrate fish communities off four islands in the Canarian Archipelago. Mar. Biol. 125: 215-231. [ Links ]
Francour P. 1997. Fish assemblages of Posidonia oceanica beds at Port-Cross (France). Assessment of composition and long-term fluctuations by visual census. PSZNI Mar. Ecol. 18: 157-173. [ Links ]
García-Charton JA, Pérez Ruzafa A, Marcos-Diego C. 2000. Fish visual census methods for detecting gradients of abundance and biomass across boundaries of MPAs. In: Goñi R, Harmelin-Vivien M, Badalamenti F, Le Diréach L, Bernard G (eds.), Introductory Guide to Methods for Selected Ecological Studies in Marine Reserves. GIS Posidonie Publ., France, pp. 29-34. [ Links ]
García-Rubies A. 1997. Estudi ecológic de les poblacions de peixes litorals sobre sustrat rocós a la Mediterránia occidental: Effects de la fondária, el sustrat, l'estacionalitat i la protecció. Tesis doctoral, Univ. Barcelona, 261 pp. [ Links ]
Guidetti P, Bussotti S. 1997. Recruitment of Diplodus annularis and Spondyliosoma cantharus in shallow seagrass beds along the Italian coasts (Mediterranean Sea). Mar. Life 7: 47-52. [ Links ]
Guidetti P, Bussotti S. 2000. Fish fauna of a mixed meadow composed by the seagrass Cymodocea nodosa and Zostera noltii in the western Mediterranean. Oceanol. Acta 23: 759-770. [ Links ]
Hernández-García V, Hernández-López JL, Castro JJ. 1998. The octopus (Octopus vulgaris) in the small-scale trap fishery off the Canary Islands (Central East Atlantic). Fish. Res. 35: 183-189. [ Links ]
Kingsford M, Battershill C. 1998. Studying Marine Temperate Environments: A Handbook for Ecologists. Canterbury Univ. Press. Christchurch, New Zealand, 335 pp. [ Links ]
Lincoln-Smith MP. 1988. Effects of observer swimming speed on sample counts of temperate rocky reef fish assemblages. Mar. Ecol. Prog. Ser. 43: 223-231. [ Links ]
Mazella L, Scipione MB, Gambi MC, Buia MC, Lorenti M, Zupo V, Cancemi G. 1993. The Mediterranean seagrass Posidonia oceanica and Cymodocea nodosa. A comparative overview. First International Conference on the Mediterranean Coastal Environment, MEDCOAST '93, Antalya, Turkey, pp. 103-116. [ Links ]
Pavón-Salas N, Herrera R, Hernández-Guerra A, Haroum R. 2000. Distributional patterns of seagrasses in the Canary Islands (Central-East Atlantic Ocean). J. Coast. Res. 16: 328-335. [ Links ]
Peduzzi P, Vukovic A. 1990. Primary production of Cymodocea nodosa in the Gulf of Trieste (northern Adriatic Sea): A comparison of methods. Mar. Ecol. Prog. Ser. 64: 197-207. [ Links ]
Pérez M. 1989. Fanerógamas marinas en sistemas estuáricos: Producción, factores limitantes y algunos aspectos del ciclo de nutrientes. Tesis doctoral, Univ. Barcelona, 244 pp. [ Links ]
Pérez M, Camp J. 1986. Distribución espacial y biomasa de las fanerógamas marinas de las bahías del Delta del Ebro. Invest. Pesq. 50: 519-530. [ Links ]
Phillips RC, McRoy CP. 1980. Handbook of Seagrass Biology: An ecosystem perspective. Garland STPM Press, New York, 353 pp. [ Links ]
Reyes J. 1993. Estudio de las praderas marinas de Cymodocea nodosa (Cymodoceaceae, Magnoliophyta) y su comunidad de epífitos, en El Médano (Tenerife, Islas Canarias). Tesis doctoral. Univ. La Laguna, 424 pp. [ Links ]
Reyes J, Sansón M, Afonso-Carrillo J. 1995. Leaf phonology, growth and production of the seagrass Cymodocea nodosa at El Médano (south of Tenerife, Canary Islands). Bot. Mar. 38: 457-465. [ Links ]
Sand-Jensen K. 1975. Biomass, net production and growth dynamics in an eelgrass (Zostera marina) populations in Vellerup Vig., Denmark. Ophelia 14: 185-201. [ Links ]
Terrados J, Ros JD. 1992. Growth and primary production of Cymodocea nodosa (Ucria) Ascherson in a Mediterranean coastal lagoon: The Mar Menor (SE Spain). Aquat. Bot. 43: 63-74. [ Links ]
Tuya F, Pérez J, Medina L, Luque A. 2001. Variaciones estacionales de la macrofauna invertebrada de tres praderas marinas de Cymodocea nodosa en Gran Canaria (centro-este del Océano Atlántico. Cienc. Mar. 27: 223-234. [ Links ]
Underwood AJ. 1997. Experiments in Ecology: Their Logical Design and Interpretation using Analysis of Variance. Cambridge Univ. Press, 504 pp. [ Links ]
Valle C. 2005. Impactos antrópicos sobre la ictiofauna litoral. Tesis doctoral, Univ. Alicante, 409 pp. [ Links ]
Vidondo B, Duarte CM, Middelboe AL, Stefansen K, Lützen T, Nielsen SL. (1997). Dynamics of a landscape mosaic: Size and age distributions, growth and demography of seagrass Cymodocea nodosa patches. Mar. Ecol. Prog. Ser. 158: 131-138. [ Links ]

