










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkCiencias marinas
versão impressa ISSN 0185-3880
Cienc. mar vol.32 no.4 Ensenada Dez. 2006
Artículos
Distribución de la macrofauna intermareal del Delta del Río Colorado, norte del Golfo de California, México
Distribution of the intertidal macrofauna of the Colorado River Delta, northern Gulf of California, Mexico
GE Avila-Serrano1*, KW Flessa2, MA Téllez-Duarte1, CE Cintra-Buenrostro2
1 Facultad de Ciencias Marinas, Universidad Autónoma de Baja California, Apartado postal 453, Ensenada CP 22800, Baja California, México. * E-mail: gavila@uabc.mx
2 Department of Geosciences, University of Arizona, Tucson, Arizona 85721, USA.
Recibido en junio de 2005
Aceptado en septiembre de 2006.
Resumen
Entre 1999 y 2000 se determinaron la composición y distribución de la macrofauna bentónica (moluscos, equinodermos y braquiópodos) en la zona intermareal del Delta del Río Colorado, al norte del Golfo de California, México. Los muestreos se obtuvieron a lo largo de diez transectos de la zona mesomareal desde la parte superior a inferior, en dos épocas del año para determinar la variación estacional de la fauna. Fueron identificadas 26 especies en 112 muestras con un total de 1954 individuos. Dominó el gasterópodo epifaunal Nassarius moestus, bivalvos y equinoideos infaunales y el braquiópodo linguloide Glottidia palmeri. La densidad de la fauna fue mayor hacia la parte inferior de la zona intermareal. La riqueza de las especies no varió significativamente entre transectos, pero la diversidad (índice de Shannon) y equitatividad fueron mayores en los transectos del sur, los más alejados de la boca del río. La diversidad y equitatividad de los transectos del sur pueden explicarse por la variación en la región de la salinidad y tipo del sustrato. Actualmente la densidad de la fauna es de 3 a 7 ind m-2, mucho menor a la reportada antes de la construcción de las presas y que afectó los hábitat en el Delta del Río Colorado, principalmente por la disminución en la población del bivalvo máctrido Mulinia coloradoensis.
Palabras clave: Delta del Río Colorado, norte del Golfo de California, moluscos marinos.
Abstract
Surveys were conducted in 1999 and 2000 to document the composition and distribution of the shelly (mollusk, echinodermi and brachiopod) fauna of the intertidal zone of the Colorado River Delta, northern Gulf of California, Mexico. Samples of the benthic fauna were taken along ten high-to-low intertidal transects, during two seasons to assess seasonal variation in the fauna. Twenty-six species were identified among 1954 individuals in 112 samples. The fauna was dominated by the epifaunal gastropod Nassarius moestus, infaunal bivalves, infaunal echinoids and the lingulid brachiopod Glottidia palmeri. Faunal density was generally greater toward the lower part of the intertidal zone. Species richness did not vary significantly among transects, although Shannon diversity and equitability were greater in the southern transects, those distant from the river mouth. The distinctive species composition, diversity and equitability of the southern transects may be the result of regional variation in salinity and substrate. Current faunal densities of 3-7 ind m-2 are much lower than the estimates before upstream dams and water diversions affected the habitats of the Colorado River Delta. The post-dam decrease in density is largely the result of the decline in the population of the mactrid bivalve Mulinia coloradoensis.
Key words: Colorado River Delta, northern Gulf of California, marine mollusks.
Introducción
La construcción de presas y encauzamiento de las aguas del Río Colorado para uso agrícola y doméstico en los Estados Unidos de Norteamérica y México, ha ocasionado prácticamente un nulo aporte de agua dulce a su delta, excepto cuando existen excedentes durante años de precipitaciones intensas (Lavín y Sánchez 1999). Esta disminución del flujo del río ha producido efectos en la hidrología (Carbajal et al. 1997, Lavín et al. 1998, Lavín y Sánchez 1999) y procesos sedimentarios (Carriquiry y Sánchez 1999) tanto en el delta como al norte del Golfo de California. La importancia del flujo de agua dulce del río es determinante en la pesquería del camarón (Aragón-Noriega y Calderón-Aguilera 2000, Galindo-Bect et al. 2000, Calderón-Aguilera et al. 2003), la reproducción de la corvina del Golfo Cynoscion othonopterus (Rowell et al. 2005) y del bivalvo Mulinia coloradoensis (Rodríguez et al. 2001a). Por otro lado, hay que destacar la importancia biológica del delta por la presencia de dos especies endémicas del norte del Golfo de California, la totoba (Totoaba macdonaldi) y la marsopa llamada localmente vaquita marina (Phocoena sinus), ambas actualmente en peligro de extinción tanto por la sobrepesca (Flanagan y Hendrickson 1976) como por la pesca incidental (D'Agrosa et al. 2000). La disminución en la diversidad de la fauna deltaica no se ha estudiado completamente, particularmente de la macrobentónica. Sólo existen unos pocos estudios muy generales, como el de Villarreal-Chávez y Carmona-Cedillo (1999), quienes realizaron un estudio preliminar de la infauna bentónica al sur del delta en la costa de Sonora, encontrando que la estructura de la comunidad está muy influenciada por la bioturbación y tamaño del sedimento.
Aun cuando la productividad en el norte del Golfo de California es alta sin aporte de agua dulce (Hernández-Ayón et al. 1993, Millán-Núñez et al. 1999), Kowalewski et al. (2000) sugieren que la productividad de la fauna de moluscos ha disminuido hasta en un 90% por la falta de flujo de agua del Río Colorado. Sus conclusiones se basan en la comparación de la abundancia de las conchas depositadas en la playa con datos preliminares de la abundancia de fauna viva.
En este artículo se reporta por primera vez un análisis cualitativo y cuantitativo de la distribución de la macrofauna viva intermareal de moluscos, braquiópodos y equinodermos en las planicies del Delta del Colorado, y proporciona un punto de comparación con la fauna presente antes de la construcción de las presas, la cual se ha determinado con las tanatocenosis recientes (Kowalewski et al. 1994). Lo anterior presenta una alternativa para evaluar el impacto ambiental del ambiente por la desviación de las aguas del río.
Materiales y métodos
La parte marina del delta del Río Colorado está dentro de la Reserva de la Biosfera del Alto Golfo de California y Delta del Río Colorado (31°01׳32°22-׳ N; 112°59׳115°13-׳ W), a lo largo de la línea de costa entre San Felipe, Baja California, y el Golfo de Santa Clara, Sonora (fig. 1).
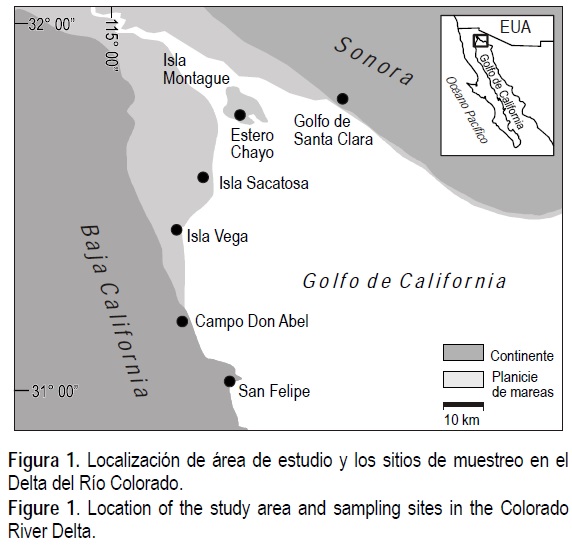
La región es cálida y árida, el promedio anual de lluvia es 68 mm (Miranda-Reyes et al. 1990), la media mensual de la temperatura del aire está entre 14°C y 34°C (Miranda-Reyes et al. 1990) y la temperatura superficial en las planicies de marea es entre 3.3°C y 40.6°C (Goodwin et al. 2001). La región es macromareal, con un máximo de 12 m en la boca del río (Thompson 1968). El amplio rango de marea y la poca pendiente en las planicies de marea, generan una planicie expuesta de hasta más de 2 km durante marea baja. Los sedimentos característicos son arenas finas y limos en el sur, y limos y lodos en el norte (Thompson 1968).
Se hicieron muestreos en cinco localidades (fig. 1, tabla 1): Campo Don Abel (CDA), Isla Vega (IV), Isla Sacatosa (IS) Estero Chayo (ECH) y aproximadamente 8 km al norte del Golfo de Santa Clara (GSC). En CDA se hicieron dos transectos paralelos separados 50 m, en octubre de 1999 (CDA 1 y 2) y mayo de 2000 (CDA 3 y 4). Los transectos de IS se realizaron en diciembre de 1999 y agosto de 2000. Este muestreo durante meses diferentes fue con el fin de detectar variaciones estacionales de la fauna.
Los transectos se orientaron perpendiculares a la línea de costa, dentro del rango del nivel medio del mar hasta el límite de la marea inferior. Los muestreos se realizaron durante las mareas más bajas del mes. Los intervalos de las estaciones en cada transecto fueron de 50 ó 100 m (tabla 1). Se muestreó un total de 112 estaciones mediante cuadrantes que cubrían un área de 1 m2 en cada estación a una profundidad de 10 cm mediante tamizado en un cernidor con una abertura de malla de 6 mm. La fauna viva se identificó y contó en el sitio, salvo la poco común cuya identificación se realizó utilizando como referencia a Keen (1971) y Keen y Coan (1974), así como la colección de referencia de la Universidad de Arizona y del Museo de Historia Natural de San Diego.
Para cada transecto se calculó la riqueza de especies (número de especies), la diversidad con el índice de Shannon, y la equitatividad con el máximo hipotético observado en el índice de Shannon (Krebs 1998). Se utilizó el índice de Bray- Curtis para medir la similitud con análisis de agrupamiento modo R y Q. Para el análisis de agrupamiento se obtuvo la abundancia absoluta para reducir el efecto de la dominancia y se usó el método de par de grupo sin peso (unweighted pair-group method). Los cálculos se realizaron con los programas Excel, Matlab 5.3 y PAST (Palaeontological Statistics) versión 1.21 (Hammer et al. 2001; http://folk.uio.no/ohammer/past/index.html).
Resultados
Composición de especies
En las 112 muestras se identificaron un total de 26 especies de 1954 individuos contabilizados (tabla 2, apéndice 1). Los datos para cada estación están disponibles en www.geo.arizona.edu/ceam/data1/. La fauna se caracteriza por la dominancia de pocas especies, ya que sólo tres de ellas comprenden 75% del total de los individuos y siete poco más de 96%. La especie más abundante fue el gasterópodo epifaunal somero Nassarius moestus, que se presentó en 53 de 112 estaciones. La segunda y tercer especie más abundante es el bivalvo infaunal somero Tellina meropsis y el equinoideo Mellita longifissa, respectivamente. Ambas especies (T. meropsis y M. longifissa) sólo se encontraron en los transectos de CDA. Un bivalvo infaunal, Chione fluctifraga, es la cuarta especie más abundante y se encontró en los transectos al norte de CDA, en 42 de las 112 estaciones. El braquiópodo infaunal somero Glottidia palmeri apareció en dos localidades (IV y GSC), además de los transectos cercanos a CDA, y se caracteriza por tener una distribución en parches. Los bivalvos infaunales someros, Donax navicula y Chione pulicaria, sólo se encontraron en los tran-sectos de CDA. Únicamente se encontró cuatro individuos del bivalvo infaunal profundo Tagelus affinis, que se distingue por la forma alargada de sus conchas para enterrarse en el sustrato.

Se encontraron 15 especímenes del molusco bivalvo epifaunal somero Mulinia coloradoensis, uno en los transectos de CDA y los otros en IV e IS. E Coan (2003, com. pers.) reporta que en breve será publicada la sinónima entre esta especie y Mulinia modesta; mientras tanto se seguirá llamando M. coloradoensis a los especímenes de esta especie en el Delta del Río Colorado, en donde alguna vez constituyeron los invertebrados calcáreos dominantes (Kowaleski et al. 2000).
Densidad
La densidad media fue de 17 ind m-2. Esta densidad fue mayor en los transectos 3 y 4 de CDA (tabla 3), probablemente por la abundancia estacional de Tellina meropsis y Donax navícula. También Mellita longifissa fue abundante en los transectos de CDA y no apareció en las otras localidades. La densidad más baja fue en el transecto IV2, donde no se encontraron organismos vivos, seguido por los transectos ECH2 e IS1 con aproximadamente 4 y 5 ind m-2, respectivamente. La mayor densidad fue durante mayo en CDA, y la más baja en IS durante agosto.
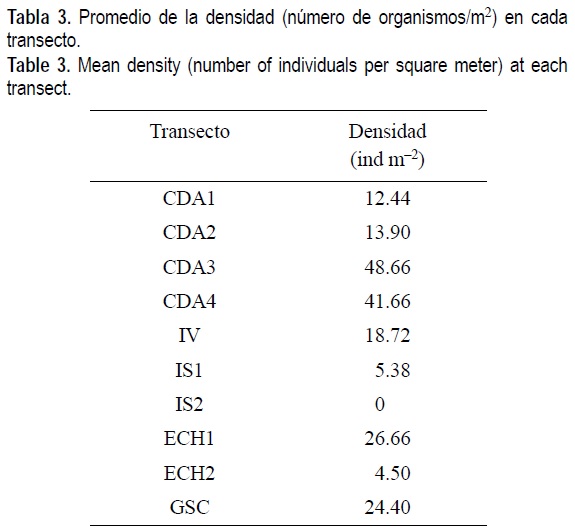
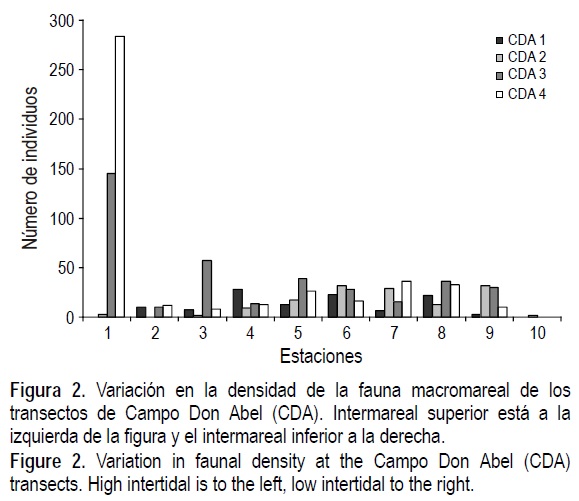
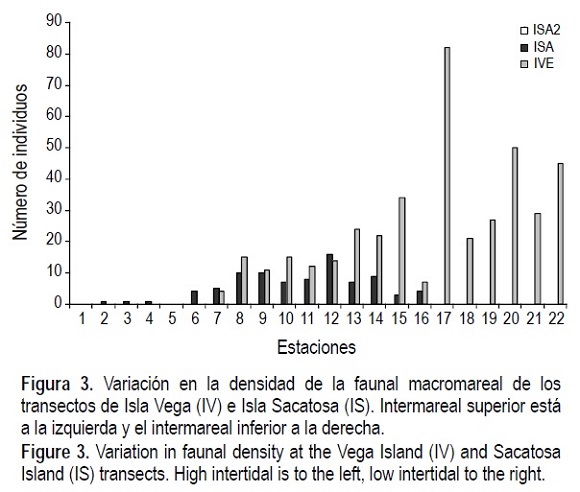
En cuanto a densidad total de organismos, considerando todos los transectos, las mayores se registraron en las estaciones del mesomareal inferior y las menores en las estaciones del mesomareal superior (figs. 2, 3, 4). La excepción fueron CDA3 y CDA4 (fig. 2), donde la especie T. meropsis fue muy abundante en las dos estaciones del mesomareal superior.
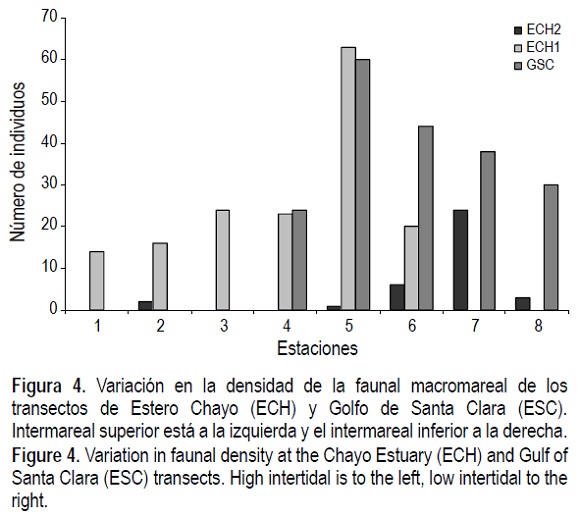
Riqueza, diversidad y equitatividad de especies
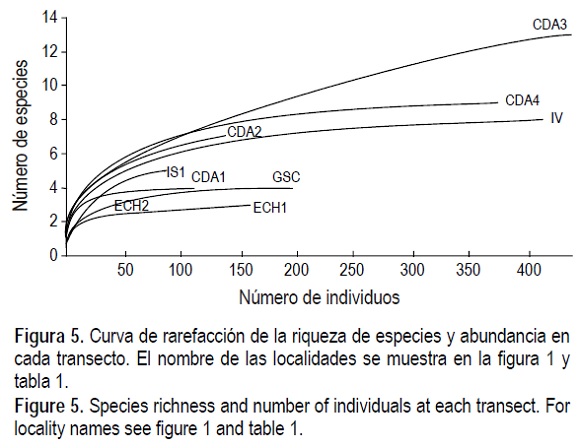
La riqueza de especies y abundancia relativa fueron mayores en los transectos de CDA e IV, en el sur del área de estudio (tabla 4, fig. 5). Las máximas riqueza y abundancia relativa se presentaron en CDA3 con 13 especies, y las mínimas ocurrieron en IS2, donde no se encontraron organismos vivos.
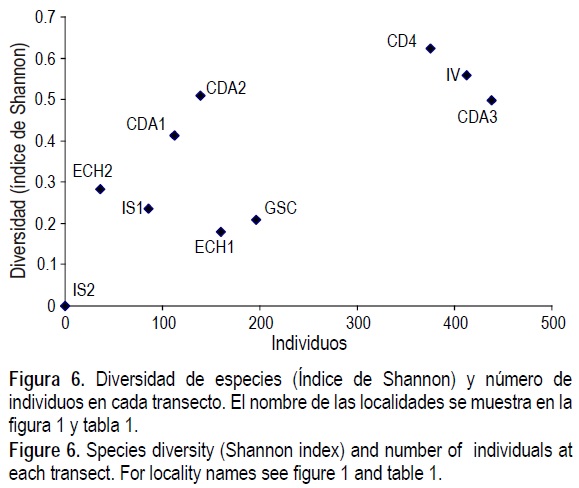
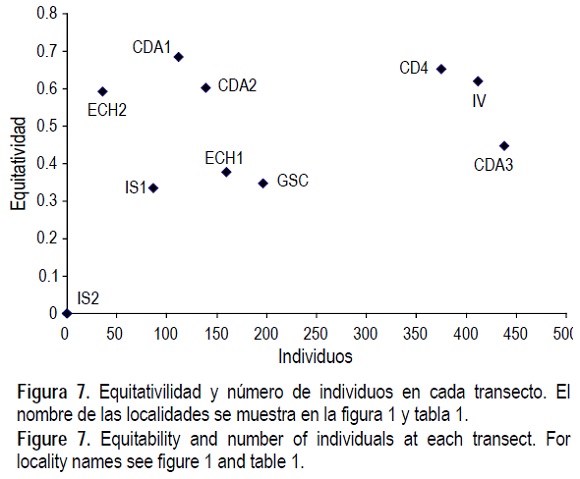
Los valores de diversidad de especies mediante el índice de Shannon fueron mayores en los cinco transectos del sur, y la equitatividad fue también alta en cuatro de estos transectos (CDA e IV), no obstante la baja abundancia de organismos en CDA1 y CDA2 (figs. 6, 7). En los cinco transectos del norte (IS1, IS2, ECH1, ECH2 y GSC), los valores de diversidad y equitatividad fueron bajos.
Distribución y asociación de especies
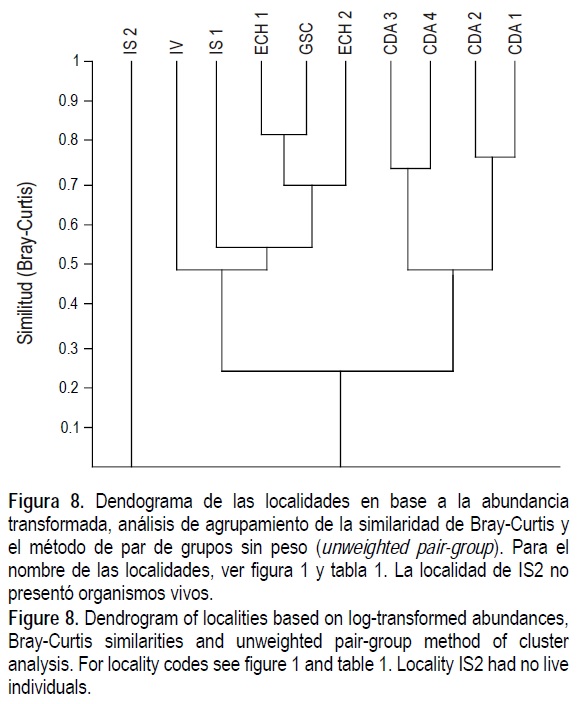
En el dendograma de localidades se forman dos agrupaciones mayores, la de mayor similitud que agrupa los cuatro transectos del sur (CDA1 a CDA4), y la segunda, que comprende el resto de localidades (fig. 8). En este segundo grupo muestran mayor similitud los tres transectos cercanos a la boca del río (ECH1, ECH2 y GSC).
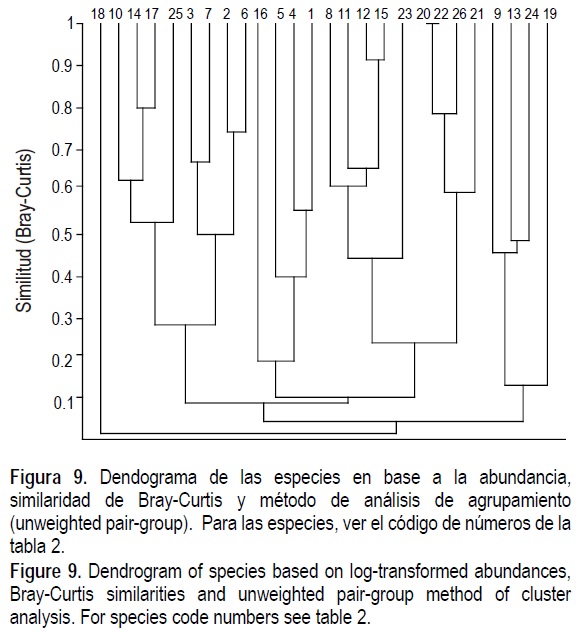
En el dendograma de especies (fig. 9) se pueden identificar varios grupos distintos. Las especies más abundantes forman tres grupos: el primero caracterizado por los ampliamente distribuidos Nassarius moestus, Chione fluctifraga y Glottidia palmeri; el segundo integrado por las especies comunes en la parte sur (CDA), Mellita longifissa, Chione pulicaria, Tellina meropsis y Donax navicula; y el tercero formado por las especies más comunes en las localidades intermedias (IV y IS1), Mulinia coloradoensis, Strigilla interrupta, Natica chemnitzi y Protothaca grata. En el resto del diagrama de agrupamiento están las especies con baja abundancia y que generan pocos cambios en la composición y arreglo del diagrama.
Discusión
Composición de especies
Como en el Delta del Colorado el sustrato es de arenas medias a lodos (Thompson 1968), están ausentes especies incrustantes y domina en la fauna mesomareal el gastrópodo
Nassarius moestus, un carroñero epifaunal. Son abundantes, en el área norte el braquiópodo Glottidia palmeri y en la sur los equinoideos sedimentívoros, someros e infaunales, Mellita longifissa y Encope grandis, porque su hábitat está limitado por las características del sustrato (Keen 1971, Keen y Coan 1974). Unas especies sólo se encontraron en el muestreo de verano, seguramente por ser su época de reclutamiento ya que predominaron organismos juveniles. Además, otras especies como el gasterópodo predador Natica chemitzi resultaron raras.
Mulinia coloradoensis tiene el menor porcentaje en la fauna viva, pero sus conchas aportan constituyen hasta 84% de las conchas de los depósitos de las playas adyacentes (Kowalewski et al. 1994). Estos depósitos están formados por las conchas de estos organismos que vivieron antes de la construcción de las presas (Kowalewski et al. 1998). Este dramático cambio de abundancia puede atribuirse a la alteración ambiental por la reducción del flujo del río hacia su delta, desapareciendo sus condiciones estuarinas del pasado. Lo anterior se deduce de estudios con isótopos de oxígeno en conchas de moluscos que vivieron antes de la construcción de las presas, los cuales indican que M. coloradoensis prosperaba en aguas menos saladas (Rodríguez et al. 2001a, 2001b).
La segunda y sexta especies más abundantes, Tellina meropsis y Donax navicula, respectivamente, sólo se presentaron en dos transectos durante mayo de 2000. Probablemente, su presencia sólo sea temporal debido a un ciclo de vida corto y por distribuirse en parches, ya que no se encontraron organismos vivos en el muestreo de octubre ni en otro transecto.
Densidad
El aumento de la densidad de organismos con la profundidad del sustrato en la zona mesomareal probablemente se deba al mayor tiempo que están cubiertos por el agua. En el meso-mareal inferior los organismos están menos tiempo expuestos, las temperaturas son menos extremas y, por tanto, son capaces de alimentarse mejor que la población de la zona mesomareal superior (Keen y Coan 1974).
No hay un patrón geográfico claro en el cambio de densidad de organismos en el área de estudio. En unos transectos de substrato lodoso del norte (ECH1, GSC) disminuye la densidad con respecto a los arenosos de la parte sur, pero otros de sustrato lodoso en el norte (IS1, IS2, ECH2) muestran una mayor variación dado que su densidad es muy baja. La disminución en la densidad de ECH2 puede ser por su elevada posición en la zona intermareal. En el campo es difícil precisar su nivel de altura, pero la presencia de la vegetación de marisma (Distichlys palmeri) confirma dicha posición intermareal.
En los dos transectos de IS (IS1 e IS2) la densidad de especies fue baja. Esta característica puede ser consecuencia de que el área recibe agua del canal de Baja California con una salinidad ~39%o, siendo de 3%o a 4%o más alta que en los transectos aledaños (Carriquiry y Sánchez 1999). La ausencia de organismos vivos en IS2 es sorprendente. Este transecto se muestreó durante agosto, la temporada más cálida del año, en condiciones de "antiniño", con viento caliente y seco procedente del norte que produce mucha turbidez. Por este alto rigor ambiental probablemente los organismos infaunales se entierran más profundo durante este periodo para evitar las altas temperaturas y poder desplazarse hacia la parte inferior de la zona mesomareal. Para resolver esta interrogante es necesario un muestreo más frecuente e intensivo.
Se obtuvo un promedio de abundancia de 17 ind m-2, la cual no se puede comparar directamente con la de 25 a 50 ind m-2 obtenida por Kowalewski et al. (2000) de las tanatocenosis anteriores a la construcción de las presas debido a que en los muestreos se utilizaron tamaños de malla diferentes. Kowalewski et al. (2000) hicieron sus observaciones con base a organismos mayores a 12.5 mm, mientras que en este trabajo se utilizó una abertura de malla de 6 mm. Sin considerar las especies menores a 12.5 mm, N. moestus, T. meropsis y D. navicula, la densidad es de aproximadamente 7 ind m-2. Los dos valores obtenidos deberían ser mayores a los de Kowalewski et al (2000), ya que en nuestras estimaciones incluimos especies como M. longifissa y G. palmeri, que son muy frágiles para preservarse en los depósitos de playa que muestrearon esos investigadores. Al eliminar las especies frágiles y pequeñas del muestreo realizado, la densidad resulta de 3 ind m-2. Por lo tanto, se concluye que a partir de que las aguas del Río Colorado fueron confinadas y desviadas, la densidad de la macrofauna intermareal, se redujo de 25-59 ind m-2, hasta 3-7 ind m2. Esto representa una disminución de entre 72% (de 25 a 7 m-2) y 94% (de 50 a 3 m-2).
Aunque la causa en la disminución de la densidad no está del todo clara, es obvio que de alguna forma influye el flujo del río hacia el delta. El río debió aportar nutrientes al delta y al norte del Golfo de California en cantidades suficientes para mantener las densas poblaciones de organismos del pasado. Sin embargo, en la actualidad la parte norte del golfo es altamente productiva, sólo que parece estar más limitada por la cantidad de luz que por la disponibilidad de nutrientes (Hernández-Ayón et al. 1993, Millán-Núñez et al. 1999). Por lo tanto, el aparente aumento de la salinidad debido a que ya no llega agua del río, puede ser la causa de la disminución de la densidad de fauna macrobentónica, como es el caso específico del bivalvo M. coloradoensis. Rodríguez et al. (2001a, 2001b) argumentan que esta especie vivía en aguas con menor salinidad que la que se presenta actualmente. Finalmente, el déficit de sedimento, que no llega por la presencia de las presas río arriba, así como la ausencia en su descarga, da como resultado que el sedimento sea erosionado y resuspendido sobre toda la planicie del delta (Carriquiry y Sánchez 1999).
Si en el estudio de Kowalewski et al. (2000) se elimina M. coloradoensis, el cálculo de la densidad es de 4-8 ind m-2, muy cerca del ajuste realizado a nuestros resultados, 3-7 ind m-2. Así, la disminución en la densidad de individuos desde que se alteró la influencia del río puede ser explicada por la drástica reducción de la población de M. coloradoensis.
Riqueza, diversidad y equitatividad de especies
Las curvas de rarefacción de los transectos muestran diferencias debido a las condiciones ambientales que caracterizan a las localidades. Quienes tienen condiciones ambientales menos rigurosas son: CDA1, ECH1, ECH2 y GSC están por abajo de los demás (fig. 5), las de CDA3 y ECH1 quienes están en los extremos, presentan la mayor diferencia por la menor interacción biológica. Las estaciones de Estero Chayo (ECH1 y ECH2), GSC, IS1 y CDA1 representan las localidades con condiciones ambientales más rigurosas, lo cual se traduce en una más baja diversidad. En CDA3, CDA4, CDA2 e IV, indican condiciones ambientales menos rigurosas y donde se da un mayor número de interacciones biológicas, lo cual se manifiesta en una mayor diversidad, lo cual cabe esperar por ser las estaciones donde existe menor variabilidad en los parámetros ambientales de la zona intermareal, como salinidad, temperatura y tiempo de exposición en las planicies de mareas.
En la comparación del índice de diversidad de Shannon y la equitatividad con el tamaño de muestra (figs. 6, 7), se observa que las localidades del sur (CDA1 a 4, IV) son muy diversas y con alta equitatividad, mientras que las del norte tienen diversidad y equitatividad baja (IS1, ECH1, ECH2, GSC). Aun cuando estos valores pueden ser afectados por el tamaño de muestra (Krebs 1998), su explicación más probable es que se relacionen con el rigor ambiental, en forma similar como se explicó con anterioridad para las curvas de rarefacción.
Distribución y asociación de especies
El dendograma por localidades separa los cuatro transectos del sur (CDA) de los del norte (IS, IV, ECH, GSC; fig. 8). Esta agrupación se explica por la diferencia de sustratos y de la salinidad. El substrato de CDA es predominantemente arena fina, en tanto hacia el norte predominan los limos y los lodos (Thompson 1968). En tanto, la salinidad es mayor en el norte por la alta evaporación y lo somero de las aguas. Además, la circulación en el sur está influenciada por el agua marina con salinidades típicas del Golfo, como son mostradas por Carriquiry y Sánchez (1999).
La figura 9 tiene un grupo característico para los cuatro transectos del sur (CDA), además de que dichas especies muestran preferencia por sustrato arenoso y salinidad marina; en el segundo grupo están las especies de las localidades intermedias (IV y IS) y, por último, el tercer grupo lo forma las especies que se encuentran más dispersas. Las localidades de más al norte, las cercanas a la boca del río, no presentan especies características.
En la macrofauna intermareal de las planicies de marea en el Delta del Río Colorado domina el gastrópodo epifaunal N. moestus, los moluscos bivalvos infaunales T. meroposis, C. fluctifraga, D. navicula, el equinoideo infaunal M. longifissa y el braquiópodo infaunal G. palmeri. La abundancia de los equinoideos se restringe al sur del área, en partes arenosas de planicie de marea, y los bivalvos T. meroposis y D. navicula sólo se encontraron en el muestreo de verano.
La densidad en los transectos, varía desde cero individuos por metro cuadrado hasta 49 ind m-2, con un promedio de 17 ind m-2. Cuando no se consideran los individuos pequeños y frágiles, la densidad promedio es 3 a 7 ind m-2. Este ajuste de la densidad es mucho menor de los 25 a 50 ind m-2 que obtuvieron antes de la construcción de las presas en el Río Colorado (Kowalewski et al. 2000). Esta disminución en la densidad de la fauna, se debe a que el molusco bivalvo M. coloradoensis no es tan abundante a partir de la construcción de las presas.
La diversidad y equitatividad en la macrofauna es mayor en los transectos del sur; sin embargo, esta también se debe al tamaño más grueso del sedimento y a la salinidad. La fauna de las localidades del sur es diferente de las otras localidades posiblemente como una consecuencia en la diferencia del sustrato y la salinidad. Las localidades cerca de la desembocadura del río no tienen una composición de especies que se distinga.
Comparando la macrofauna intermareal del Delta del Río Colorado actual con la que existía antes de la construcción de las presas, ahora ésta es menos abundante y la especie que dominaba, M. coloradoensis, ya no es tan común. El aumento de la salinidad en el norte del Golfo de California y los cambios del régimen sedimentario a partir de la disminución del flujo del Río Colorado son los dos factores que explican estos cambios.
Agradecimientos
Agradecemos a C Rodriguez, D Goodwin, A Garry, E Johnson, M Juárez, M Marrón, D Sánchez, R Ulloa y S Ramos por su ayuda en el trabajo de campo. Este trabajo fue apoyado por la Universidad Autónoma de Baja California, Facultad de Ciencias Marinas, CONACYT proyecto 28122T y National Science Foundation. Ésta es la publicación número 51 del CEAM.
Referencias
Aragón-Noriega EA, Calderón-Aguilera LE. 2000. Does damming of the Colorado River affect the nursery area of blue shrimp Litopenaeus stylirostris (Decapoda; Penaeidae) in the upper Gulf of California? Rev. Biol. Trop. 48: 867-871. [ Links ]
Calderón-Aguilera LE, Marinone SG, Aragón-Noriega EA. 2003. Influence of oceanographic processes on the early life states of the blue shrimp (Litopenaeus stylirostris) in the Upper Gulf of California. J. Mar. Syst. 39: 117-128. [ Links ]
Carbajal N, Souza A, Durazo R. 1997. A numerical study of the ex- ROFI of the Colorado River. J. Mar. Syst. 12: 17-33. [ Links ]
Carriquiry JD, Sánchez A. 1999. Sedimentation in the Colorado River delta and upper Gulf of California after nearly a century of discharge loss. Mar. Geol. 158: 125-145. [ Links ]
D'Agrosa C, Lennert-Cody CE, Vidal O. 2000. Vaquita by-catch in Mexico's artisan gillnets fisheries: Driving a small population to extinction. Conserv. Biol. 14: 1110-1119. [ Links ]
Flanagan CA, Hendrickson JR. 1976. Observations on the commercial fishery and reproductive biology of the totoaba, Cynoscion macdonaldi, in the northern Gulf of California. Fish. Bull. 74: 531-544. [ Links ]
Galindo-Bect MS, Glenn EP, Page HM, Galindo-Bect LA, Hernández-Ayón JM, Petty RL, García-Hernández J. 2000. Analysis of the penaeid shrimp catch in the northern Gulf of California in relation to Colorado River discharge. Fish. Bull. 98: 222-225. [ Links ]
Goodwin DH, Flessa KW, Schöne BR, Dettman DL. 2001. Cross-calibration of daily increments, stable isotope variation and temperature in the Gulf of California bivalve mollusk Chione cortezi: Implications for paleoenvironmental analysis. Palaios 16: 387-398. [ Links ]
Hammer O, Harper DAT, Ryan PD. 2001. PAST: Palaeontological statistics software package for education and data analysis. Palaeontologia Electronica 4(1): 9 pp. [ Links ]
Hernández-Ayón JM, Galindo-Bect MS, Flores-Báez BP, Álvarez-Borrego S. 1993. Nutrient concentrations are high in the turbid waters of the Colorado River Delta. Estuar. Coast. Shelf Sci. 37: 593-602. [ Links ]
Keen AM. 1971. Sea Shells of Tropical West America: Marine Mollusks from Baja California to Peru. 2nd ed. Stanford Univ. Press, Stanford, California, 1064 pp. [ Links ]
Keen AM, Coan E. 1974. Marine Molluscan Genera of Western North America: An Illustrated Key. 2nd ed. Stanford Univ. Press, Stanford, California, 208 pp. [ Links ]
Kowalewski M. 1996. Taphonomy of a living fossil: The lingulide brachiopod Glottidia palmeri from Baja California, Mexico. Palaios 11: 244-265. [ Links ]
Kowalewski M, Flessa KW, Aggen JA. 1994. Taphofacies analysis of recent shelly cheniers (beach ridges), northeastern Baja California. Facies 31: 209-242. [ Links ]
Kowaleski M, Goodfriend GA, Flessa KW. 1998. High-resolution estimates of temporal mixing within shell beds: The evils and virtues of time-averaging. Paleobiology 24: 287-304. [ Links ]
Kowalewski M, Avila-Serrano GE, Flessa KW, Goodfriend GA. 2000. Dead delta's former productivity: Two trillion shells at the mouth of the Colorado River. Geology 28: 1059-1062. [ Links ]
Krebs CJ. 1998. Ecological Methodology. 2nd ed. Pearson Benjamin Cummings, San Francisco, 620 pp. [ Links ]
Lavín MF, Sánchez S. 1999. On how the Colorado River affected the hydrography of the upper Gulf California. Cont. Shelf Res. 19: 1545-1560. [ Links ]
Lavín MF, Godínez VM, Álvarez LG. 1998. Inverse-estuarine features of the upper Gulf of California. Estuar. Coast. Shelf Sci. 47: 769795. [ Links ]
Millán-Núñez R, Santamaría-del-Ángel E, Cajal-Medrano R, Barocio-León OA. 1999. El delta del Río Colorado: Un ecosistema con alta productividad. Cienc. Mar. 25: 509-524. [ Links ]
Miranda-Reyes F, Reyes-Coca S, García-López J. 1990. Climatología de la región noroeste de México. Parte I. Precipitación. Rep. Tec. EBA No. 3, CICESE, Ensenada, Baja California, México, 160 pp. [ Links ]
Rodríguez CA, Flessa KW, Dettman DL. 2001a. Effects of upstream diversion of Colorado River water on the estuarine bivalve mollusc Mulinia coloradoensis. Conserv. Biol. 15: 249-258. [ Links ]
Rodríguez CA, Flessa KW, Téllez-Duarte MA, Dettman DL, Avila-Serrano GE. 2001b. Macrofaunal and isotopic estimates of the former extent of the Colorado River estuary, Upper Gulf of California, Mexico. J. Arid Environ. 49: 183-193. [ Links ]
Rowell KR, Flessa KW, Dettman DL, Román M. 2005. The importance of Colorado River flow to nursery habitats of the Gulf Corvina (Cynoscion othonopterus). Can. J. Fish Aquatic Sci. 62: 2874 -2885. [ Links ]
Thompson RW. 1968. Tidal flat sedimentation on the Colorado River delta, northwestern Gulf of California. Bull. Geol. Soc. Am. Mem. 107, 133 pp. [ Links ]
Villarreal-Chávez G, Carmona-Cedillo CM. 1999. Estructura de la comunidad bentónica (infauna) en el delta del Río Colorado, México. Rev. Biol. Trop. 47: 809-812. [ Links ]

