










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkCiencias marinas
versão impressa ISSN 0185-3880
Cienc. mar vol.32 no.2 Ensenada Jun. 2006
Artículos
Corales pétreos (Scleractinia) de las Islas Marías, Pacífico de México
Stony corals (Scleractinia) from the Marías Islands, Mexican Pacific
TL Pérez-Vivar1, H Reyes-Bonilla2*, C Padilla3
1 Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México, Apartado postal 70-305, Ciudad Universitaria, CP 04510 México DF, México.
2 Departamento de Biología Marina, Universidad Autónoma de Baja California Sur, Apartado postal 19-B, CP 23080 La Paz, Baja California Sur, México. *E-mail: hreyes@uabcs.mx
3 Centro Regional de Investigaciones Pesqueras Puerto Morelos, Instituto Nacional de la Pesca, Calle Matamoros No. 7 CP 77580 Puerto Morelos, Quintana Roo, México.
Recibido en febrero de 2005;
Aceptado en marzo de 2006.
Resumen
Estudios recientes llevados a cabo en la costa occidental de México han dado a conocer un gran número de arrecifes o parches coralinos en la región, pero aún hay áreas poco conocidas. Una de ellas es el Archipiélago de las Islas Marías (21°N), formado por cuatro islas: María Madre, María Magdalena, María Cleofas y San Juanito. El objetivo de este trabajo fue llevar a cabo una evaluación general de la situación de los corales del archipiélago por medio de la revisión del elenco sistemático de las especies, y del análisis de la estructura comunitaria y los patrones biogeográficos de la fauna. En agosto de 1997 se visitaron 21 sitios en la Isla María Madre y 2 en San Juanito, y en cada uno se realizó un transecto de video (150 m de largo) para denotar la zonación coralina y uno de línea (20 m de largo) para hacer análisis cuantitativos. Esa información fue complementada con revisiones de literatura y datos de ejemplares de museos de México y Estados Unidos. Una vez obtenido el elenco sistemático de corales de las islas, se calculó el nivel de similitud de la fauna con el de cinco regiones cercanas (incluyendo las Islas Revillagigedo y el margen continental entre 23°N y 18°N), y se condujeron análisis de ordenación. En el campo se observaron 13 especies de corales pétreos (incluyendo ocho nuevos registros), pero no se encontró a Porites baueri, especie nominalmente endémica de las Islas Marías. Las revisiones de museos arrojaron dos nuevos registros. Así, sumando la información se conocen 20 especies como residentes de la zona de estudio. En el archipiélago no se vieron arrecifes en sentido construccional pero había gran abundancia de coral, especialmente en la Isla María Madre. Las comunidades mostraban una clara zonación con Pocillopora en agua somera y Porites, Pavona y Psammocora en sitios profundos. Finalmente, el análisis biogeográfico indicó que la boca del Golfo de California representa una región clave para la inmigración de especies desde el Pacífico occidental, y las Islas Marías parecen jugar un punto nodal para la dispersión coralina hacia el norte.
Palabras clave: Islas Marías, Scleractinia, Pacífico mexicano, arrecifes coralinos, biogeografía.
Abstract
Recent studies performed off the Pacific coast of Mexico have shown a large number of reefs or coral patches in the region, but information is still lacking for some areas. One of these is the Marías Archipelago (21°N), composed of four islands: María Madre, María Magdalena, María Cleofas and San Juanito. The objective of this paper was to make a general evaluation of the status of the corales in the archipelago by revising the systematic list of species and analyzing the community structure and biogeographic patterns of the fauna. Twenty-one locations at María Madre Island and two at San Juanito Island were visited in August 1997, and at each we conducted a video transect (150 m long) to describe zonation and a line transect (20 m long) for quantitative analyses. This information was complemented with literature reviews and data from specimens deposited in museums in Mexico and the United States. After obtaining the systematic list of corals from the islands, we compared the level of similarity of the fauna with that from five nearby regions (including the Revillagigedo Islands and the mainland from 23°N to 18°N), and ordination analyses were run. In the field we observed 13 species of stony corals (including eight new records), but did not find Porites baueri, a nominally endemic species from the Marías Islands. The museum revisions produced two new records. Thus, analyzing the full data set, 20 species are considered residents of the study area. There were no constructional reefs in the archipelago, but coral abundance was high, especially at María Madre Island. The communities showed a clear zonation, with Pocillopora in shallow water and Porites, Pavona and Psammocora in deeper localities. Finally, the biogeographic analysis indicated that the mouth of the Gulf of California represents a key area for species immigration from the western Pacific, and that the Marías Islands seem to act as a nodal point for coral dispersal to the north.
Key words: Marías Islands, Scleractinia, Mexican Pacific, coral reefs, biogeography.
Introducción
Trabajos recientes llevados a cabo en el Pacífico mexicano han dado a conocer un gran número de localidades caracterizadas por la presencia de arrecifes o parches coralinos, las cuales están distribuidas a lo largo de la costa y en las islas oceánicas adyacentes (Reyes-Bonilla 2003). Aunque los datos disponibles se han incrementado sustancialmente, aún existen sitios poco estudiados, entre ellos el Archipiélago de las Islas Marías. Este conjunto insular está situado al oeste de la costa del Estado de Nayarit, y está formado por cuatro islas: María Madre, María Magdalena, María Cleofas y el islote de San Juanito (fig. 1). La región presenta un clima semiárido (García 1989), con temperaturas del aire de 18°C a 22°C en invierno y de entre 30°C y 34°C en verano (Vidal-Zepeda 1989). La temperatura superficial del mar en la zona fluctúa entre 24°C y 28°C, con promedio de 26.5°C, y la salinidad media es de 34.5 ups (WOA 2001). Las Islas Marías se encuentran cercanas a Bahía de Banderas, una importante zona de surgencia en el Océano Pacífico que presenta valores medios de fosfatos (0.42 μM), silicatos (2.8 μM) y nitratos (0.6 μM) que la hacen estar entre las zonas con mayor concentración de nutrientes en el Pacifico mexicano (WOA 2001).

En la más grande de las islas (María Madre) se localiza un penal de alta seguridad desde principios del siglo XX (Revueltas 1988), por lo que el acceso al archipiélago está restringido. La condición de aislamiento de las islas aunada a la mínima explotación ejercida sobre sus recursos naturales (la zona sólo fue usada para extracción de madera y de sal entre 1880 y 1905; Guzmán 1959) ha hecho que sus comunidades marinas se encuentren en un excelente estado de conservación (Small 1998), y ello motivó a las autoridades federales de México a declarar al archipiélago como Reserva de la Biosfera (Diario Oficial de la Federación, 27 de noviembre de 2000).
En la literatura existen varios registros secundarios sobre la presencia de corales en el archipiélago (Wells 1988, Reyes-Bonilla y López-Pérez 1998, Spalding et al. 2000), pero hay únicamente dos estudios de primera mano. Durham y Barnard (1952) recolectaron un ejemplar de Pocillopora capitata Verrill, 1864 (citado como P. robusta Verrill, 1870, un sinónimo; Reyes-Bonilla 2002), en "la Isla Isabel o en las Islas Marías". Posteriormente, Squires (1959) refirió la presencia de siete especies en el archipiélago, una de las cuales (Porites baueri Squires, 1959) es endémica. No existen más datos disponibles, aunque Carriquiry y Reyes-Bonilla (1997) citaron a las especies de las islas dentro del listado de los corales de Nayarit, y Reyes-Bonilla et al. (1999) y Glynn (2000) mencionaron que P. baueri puede ser una identificación errónea de Porites lobata Dana, 1846, coral del Indo Pacífico que ha sido observado en México en las Islas Revillagigedo, y de Nayarit a Guerrero (Reyes-Bonilla 2003, Reyes-Bonilla et al. 2005). Lo escasez de información hace evidente la necesidad de generar más datos sobre los sistemas marinos de las Islas Marías, por lo que este estudio se desarrolló con el objetivo de llevar a cabo una evaluación general de la situación de los corales por medio de la revisión del elenco sistemático de las especies, y del análisis de la estructura comunitaria y la situación biogeográfica de la fauna.
Métodos
En agosto de 1997 se visitaron las dos islas más norteñas del archipiélago, María Madre y San Juanito, y se llevaron a cabo revisiones de las asociaciones coralinas. En cada isla se realizaron prospecciones cualitativas de presencia y ausencia de especies de corales pétreos en 21 y 2 estaciones (respectivamente) situadas a lo largo del litoral, por medio de buceo libre y autónomo, a profundidades de 2 a 15 m. Además, en cada estación se efectuó un transecto de video de 150 m de longitud en dirección perpendicular a la costa desde 1 m de profundidad, con el fin de obtener perfiles de la comunidad coralina; el buzo se colocaba a una distancia aproximada de 1 m sobre el fondo y filmaba un área de 40 x 40 cm siguiendo una cinta de fibra de vidrio. Finalmente, se hicieron transectos de línea de 20 m de largo, paralelos a la costa, y a profundidades de 3 a 8 m (la principal zona de presencia coralina), con el fin de registrar la abundancia de las especies coralinas. En este caso, se identificaron las colonias que intersectaron puntos marcados a cada 20 cm de la línea, y la cobertura (en porcentaje) se calculó dividiendo el número de puntos con presencia de corales entre 120 (número total), y multiplicándolo por 100 (English et al. 1997). La identificación de los especímenes se hizo in situ, principalmente con base en los criterios de Wells (1983) y Veron (1986), y se recolectaron ejemplares de varias especies que luego fueron incorporados a la colección del Museo de Historia Natural de la UABCS (La Paz, BCS, México). Adicionalmente, se visitaron las siguientes colecciones en los Estados Unidos: California Academy of Sciences en San Francisco (CASIZ), University of California Museum of Paleontology en Berkeley (UCMP), Santa Barbara Museum of Natural History (SBMNH), Natural History Museum of Los Angeles County (IC-LACM) y Scripps Institution of Oceanography (SIO) en San Diego, de donde se tomaron los registros de los ejemplares recolectados en las Islas Marías. Finalmente, además de los datos biológicos se tomó la temperatura superficial del agua en cada sitio entre las 11 y 13 horas, con un termómetro digital sumergible (precisión ±0.1°C).
El elenco sistemático de las especies zooxanteladas presentes en las islas (definido con base en datos de museos, literatura y de campo) fue comparado con el de cinco áreas cercanas: Cabo Pulmo, en el Golfo de California (23°N), costa de Nayarit (20°N), costa de Jalisco (19°N), costa de Colima (19°N) y las Islas Revillagigedo (18°N). Posteriormente se calculó el nivel de similitud cualitativa entre sitios con el índice de Bray-Curtis, y se hizo un análisis de ordenación con la técnica de escalamiento multidimensional no métrico (Clarke y Warwick 2001). Ésta permite construir un "mapa" de semejanza entre sitios trazado en dos o tres dimensiones, y puntualizar agrupaciones que tienen significancia estadística definida por el nivel de deformación que presente el mapa ("stress"), el cual debe ser menor a 0.10. Al final se esbozó el árbol de distancia mínima entre localidades ("minimum spanning tree"; Quinn y Keough 2002), que por medio de líneas dibujadas en un gráfico de NMDS conecta todos los puntos en una red y además da a conocer qué pares discretos de localidades tienen mayor semejanza entre ellas. Se considera que dicho trazo es un indicativo confiable de las rutas más parsimoniosas de conectividad entre sitios (Crisci et al. 2003).
Resultados
Elenco sistemático
En 1997 se observaron 13 especies de escleractinios en las Islas Marías, pertenecientes a siete géneros y seis familias (incluyendo en esa cifra a cinco que habían sido registradas en el archipiélago con anterioridad; tabla 1). Además de éstas, entre 1959 y 1960 fueron recolectados en la zona ejemplares de otras dos que no habían sido referidos en la literatura, no obstante estar depositados en colecciones estaounidenses (UCMP y SIO). Por último, existen sobre otras cinco especies anotaciones que provienen exclusivamente de la literatura. Así, el listado actual de corales pétreos del archipiélago incorpora 20 especies de nueve géneros y ocho familias (tabla 1).
La revisión de campo produjo ocho nuevos registros para el archipiélago y entre ellos se incluye la primera mención de corales azooxantelados: Tubastraea coccinea Lesson, 1834, Astrangia equatorialis (Durham y Barnard, 1952) y Oulangia bradleyi (Verrill, 1866). La característica principal de estos taxa en la zona de estudio fue que normalmente ocupaban posiciones crípticas en los ambientes arrecifales, ya sea en cuevas, paredes o en la parte inferior de las rocas. Entre las especies zooxanteladas vistas por primera vez en la localidad es particularmente llamativa la presencia de Pocillopora eydouxi Milne Edwards y Haime, 1860, y de Pavona varians Verrill, 1864, ya que aunque estos corales ocurren en otros arrecifes de México (Reyes-Bonilla 2003), sólo son comunes en las Islas Revillagigedo. Otros hallazgos para las Islas Marías son Pavona clavus Dana, 1846, Pavona gigantea Verrill, 1866 y Porites lobata, especies que también se distribuyen en la costa de Nayarit y Jalisco (Reyes-Bonilla y López-Pérez 1998). Finalmente, la existencia en colecciones científicas de dos colonias de Pocillopora damicornis (L. 1758) recolectadas en María Cleofas en 1959 (UCMP 160733 y 161951), y de un ejemplar de Heterocyathus aequicostatus Milne Edwards y Haime, 1848, tomado de las "Islas Marías" en 1960 (SIO 1293), también fueron relevantes. La primera especie es una de las dominantes en arrecifes coralinos a todo lo largo del Pacífico americano (Glynn y Ault 2000); sin embargo, no fue vista en 1997 ni había sido anotada para las Islas Marías por otros autores. La segunda habita aguas profundas y es relativamente inconspicua, quizá por lo cual sólo había sido registrada una vez para México con anterioridad (en la costa de Jalisco; Durham y Barnard 1952). Contrario a estos hallazgos, es interesante que durante el trabajo de campo no se hallaron colonias que pudieran ser asignadas inequívocamente a la especie nominal Porites baueri, aunque sí muchas de P. lobata, lo que puede apoyar la idea de que la primera es en realidad un sinónimo de la segunda (Reyes-Bonilla et al. 1999). Tampoco se denotaron especímenes de Fungia distorta Michelin, 1842 ni de F. curvata Hoeksema, 1989, las cuales fueron registradas originalmente en las islas por Squires (1959) bajo sus sinónimos Cycloseris mexicana Durham, 1947 y C. elegans Verrill, 1866, respectivamente.
Descripción de las comunidades
Las islas María Madre y San Juanito no presentaron arrecifes en el sentido construccional, sino acumulaciones de colonias de diversos tamaños y parches compuestos de extensiones grandes de Pocillopora, quizá originados en gran parte por acopio y regeneración de fragmentos desprendidos por ciclones u otros fenómenos. No obstante, la cobertura de coral superó 40% del sustrato en varios de los sitios estudiados en María Madre y San Juanito (fig. 2a). Las comunidades coralinas se desarrollaban en el margen costero de las islas y principalmente sobre paredones y parches rocosos rodeados de arena. También se pudo observar acumulación de sedimentos finos sobre las rocas, lo cual evidenciaba el arrastre de material desde la porción emergida de las islas. La zonación arrecifal es clara ya que las especies de Pocillopora son dominantes en todos los sitios (fig. 2b), especialmente entre el intermareal y 6 m de profundidad, y era particularmente notable la abundancia de p capitata. Secciones de la isla entre -6 y -10 m estaban caracterizadas por la ocurrencia de Porites lobata y Pavona clavus.

A escala de regiones, la porción sureste de la Isla María Madre fue la que tuvo mayor presencia de sustrato rocoso y presentó alta abundancia coralina (fig. 2a). Las colonias que se encontraron eran grandes (hasta >1 m de diámetro en Pocillopora y >2 m en Pavona), aunque también hubo abundantes coralla de Pavona clavus y Porites lobata de tallas menores a 30 cm de diámetro. En esta zona también había rocas grandes con grietas profundas, las cuales estaban ocupadas con frecuencia por Tubastraea coccinea. En la parte noreste de la isla se observó un pavimento de aproximadamente 50 m de ancho donde se había desarrollado una comunidad con alta cobertura (promedio superior a 30%; fig. 2a), compuesta por colonias de Pocillopora de tallas medianas (30 a 50 cm de diámetro), y con cierta abundancia de Porites (fig. 2b). En claro contraste, la zona noroeste de la isla estaba dominada por sustratos arenosos, la cobertura de coral era mínima (>10%; fig. 2), y sólo se hallaron colonias aisladas de Pocillopora spp. y ocasionalmente, de Porites lobata. En esta parte se encuentra el faro de la isla, el cual está sobre un promontorio rocoso que bajo la superficie presentaba abundantes colonias de Pavona, así como de especies azooxanteladas (Oulangia bradleyi y Astrangia equatorialis).
La Isla San Juanito se caracterizó por tener aguas más claras que María Madre, posiblemente por su menor extensión y elevación, lo cual reduce el aporte de material terrígeno. Esa localidad se caracterizó por contar con una línea de rompiente formada de grandes rocas separadas unos 30 m de la franja de costa, donde la comunidad de corales estaba bien desarrollada y se constituía de especies de Pocillopora y Porites. Bajo las rocas se podían advertir grandes colonias de Tubastraea y en agua profunda apareció Pavona spp. Finalmente, de acuerdo con Small (1998), la parte sureste de San Juanito tiene un pavimento rocoso similar al visto en el noreste de María Madre, pero la zona no fue visitada por los autores.
Durante el trabajo de campo se observó en ambas islas la presencia de colonias aisladas de Pocillopora que presentaban blanqueamiento, es decir, la pérdida o decoloración de sus zooxantelas simbiontes. El fenómeno se advirtió sobre todo en aguas someras (<3 m), no se presentó en corales de otros géneros, y aquí se sugiere que éste debió haber sido causado por las elevadas temperaturas medidas en la localidad en agosto de 1997 (32°C a 34°C en superficie).
Aspectos biogeográficos
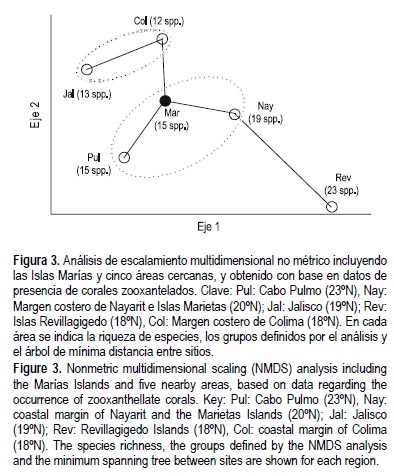
El análisis de ordenación que incluyó a las Islas Marías y a las cinco regiones arrecifales más cercanas al sitio en México (datos en tabla 1 y Apéndice), indicó que las islas están incluidas en un grupo junto con Cabo Pulmo (en el sur del Golfo de California) y la costa de Nayarit, el cual es relativamente independiente del formado por Jalisco y Colima, y de las Islas Revillagigedo (fig. 3). Además, se encontró que de acuerdo con el árbol de distancia mínima, la región de la entrada del Golfo de California (representada por las Islas Marías, Cabo Pulmo y la costa de Nayarit) funciona como un punto intermedio de conectividad entre los grupos mayores denotados por el escalamiento multidimensional, y que las Islas Marías deben jugar un papel destacado en este proceso.
Discusión
El contraste en el elenco de especies registradas en las Islas Marías antes de este trabajo (9 especies) y el actualizado en la tabla 1 (20 especies) se puede deber a diversos factores. Primero podemos citar las diferencias entre conceptos taxonómicos. A mediados del siglo XX los investigadores tendían a considerar que las colonias sufrían modificaciones en sus características debido a diferencias en condiciones ambientales o geográficas (Squires 1959), y por ello agrupaban muchas formas dentro del mismo taxón. Sin embargo, estudios posteriores han demostrado que aunque existen especies polimórficas, la mayoría de las veces la variación en la forma es una pista eficiente para separar entre especies válidas (Veron 2000). La falta de reconocimiento de estas "especies hermanas" aparentemente ha hecho que se subestime la riqueza específica en los arrecifes de coral (Knowlton 1993). Dentro de este apartado puede mencionarse también la relevancia que tuvo la revisión de ejemplares en diversos museos de México y los Estados Unidos para complementar el listado de especies, ya que la labor permitió la inclusión en la tabla 1 de material recolectado por otras expediciones y cuyos datos seguían inéditos.
Por otra parte, en nuestro estudio se realizó un gran esfuerzo de muestreo (siete días de trabajo) que, presumiblemente, es el más elevado hasta la fecha ya que en visitas previas las partidas permanecieron en el archipiélago un máximo de tres días (Durham y Barnard 1952, Squires 1959). También, durante el trabajo de campo se revisaron diversos hábitats y una extensión considerable de los arrecifes, lo cual debe haber ayudado a mejorar la probabilidad de encuentro de especies poco abundantes. Esta investigación además contó con la ventaja que ofrece el uso de buceo autónomo para el recorrido de las áreas, lo cual facilitó la búsqueda de organismos y permitió hacer recolectas manuales donde se pudieron seleccionar colonias a mayor profundidad y con mucho mayor detalle que en estudios previos donde se usó buceo libre (Squires 1959). También nos habilitó para obtener material en paredes, cuevas o los mismos macizos coralinos donde es relativamente difícil conducir dragados como los efectuados en investigaciones anteriores (Durham y Barnard 1952). A pesar de lo anterior, es importante notar que hubo dos especies del género Fungia citadas por Squires (1959) que no fueron vistas por nosotros, ni se encontraron ejemplares depositados en colecciones científicas. Es posible que la falta de registro se deba a que tanto en el estudio de 1997 como en los anteriores, los muestreos y observaciones no se efectuaran en zonas arenosas, que son el hábitat más común de ocurrencia de las especies de la familia Fungiidae (Squires 1959, Veron 2000).
El sistema de desarrollo de los parches coralinos encontrados en las Islas Marías corresponde al observado frecuentemente en áreas como Bahía San Gabriel y Cabo Pulmo, en el Golfo de California, y en las Bahías de Huatulco, Oaxaca (Glynn y Leyte-Morales 1997, Reyes-Bonilla 2003), donde los arrecifes han crecido por acumulación de fragmentos de Pocillopora. El acaparamiento de espacio en agua somera resultante de la colonización del sustrato por los pocilopóridos puede ser la clave para explicar la clara zonación observada en las Islas Marías, y paralelamente ser la razón por la cual los corales de los géneros Porites, Pavona y Psammocora aparecieron casi siempre de -6 m a -15 m aun cuando tienen la capacidad de presentarse en sitios menos profundos del Golfo de California, Oaxaca y Centroamérica (Guzmán y Cortés 1993, Glynn y Leyte-Morales 1997, Reyes-Bonilla 2003). También es posible que el tipo de zooxantela ("clado") y la influencia de peces de la familia Pomacentridae estuvieran ayudando a definir la zonación, tal como se ha sugerido que ocurre en el sur del Golfo de California y Panamá (Wellington 1982, Iglesias-Prieto et al. 2004). Sin embargo, es necesario realizar estudios más detallados con el fin de dilucidar con mayor precisión cuáles son los agentes causales de los patrones de distribución coralina observados en las Islas Marías.
Los niveles de abundancia de coral registrados en las visitas de agosto de 1997 a las Marías (fig. 2) son comparables a los observados a mediados de esa década en el resto del Pacífico mexicano (ver resumen en tabla 2 de Carriquiry y Reyes-Bonilla 1997). Lo mismo puede decirse en relación con la abundancia relativa de los distintos géneros, ya que Pocillopora era el más importante en todo el occidente de México, seguido por Porites, Pavona y Psammocora, en ese orden (Reyes-Bonilla 1993, Guzmán y Cortés 1993), situación que aún prevalece (Reyes-Bonilla 2003). En resumen, las asociaciones coralinas observadas en las Islas Marías a mediados de 1997 mostraban los patrones esperados en una comunidad de su tipo. No obstante, hay que remarcar que la situación actual de los arrecifes del archipiélago es desconocida y puede ser notablemente distinta a la descrita ya que la zona, al igual que toda la costa occidental de las Américas, estuvo afectada en 1997 por el evento de El Niño más fuerte del siglo. Este fenómeno trajo como resultado un incremento notable en la temperatura superficial en el Océano Pacífico (Glynn 2000), y particularmente en el Pacífico mexicano causó fuertes blanqueamientos y mortalidad coralina debido a que las anomalías térmicas superaron los 2°C por varios meses (Reyes-Bonilla et al. 2002). Considerando que el área más afectada del litoral mexicano fue la Bahía de Banderas, donde la mortalidad superó 60% de la cobertura coralina viva (Carriquiry et al. 2001), es factible que en las Islas Marías se haya presentado una situación semejante dada su cercanía geográfica y el hecho que durante nuestra visita ya se denotaba un incremento de más de 4°C sobre la temperatura máxima mensual registrada en promedio para la zona (Carriquiry et al. 2001).
A pesar de que el blanqueamiento de coral de 1997 afectó notablemente la estructura comunitaria en el Pacífico mexicano, no hay evidencias de que haya causado extirpaciones locales de especies en ningún sitio de la región (Glynn 2000, Reyes-Bonilla 2003); por ello, el estudio biogeográfico presentado aquí y centrado en las especies zooxanteladas (para las cuales hay datos muy confiables de distribución geográfica; Reyes-Bonilla y López-Pérez 1998, Reyes-Bonilla et al. 2005), debe ser robusto. El análisis de ordenación (fig. 3) evidenció que las comunidades coralinas de las Islas Marías son muy semejantes a las encontradas en la costa de Nayarit y a las del sur del Golfo de California, y que tienen ciertas discrepancias en la composición de sus ensamblajes comparadas con áreas continentales más sureñas como Jalisco y Colima, o con islas oceánicas como las Revillagigedo. Este arreglo es ligeramente distinto al propuesto por Glynn y Ault (2000), quienes sugirieron que la zona de las Revillagigedo y el Pacífico tropical mexicano (de Nayarit a Oaxaca) son más parecidos entre sí que con el Golfo de California. La diferencia de concepto debe resultar de la mejoría de información lograda durante el último lustro (Reyes-Bonilla et al. 2005), que nos permite tener ahora una idea más clara de la situación biogeográfica del grupo en el Pacífico de México. No obstante, ambos trabajos coinciden en anotar que la región de la boca del Golfo de California es un sitio de particular interés biogeográfico.
El árbol de distancia mínima arroja información que confirma lo anterior al indicar que la entrada del Golfo de California tiene una conexión importante con la zona de las Revillagigedo, la cual se caracteriza por ser un sitio clave para la inmigración de especies del Indo Pacífico hacia México (Ketchum y Reyes-Bonilla 2001, Robertson y Allen 2002). Según la figura 3, la conectividad entre la costa de Nayarit y las Revillagigedo es alta, y quizá la primera zona sea el sitio principal de entrada a México para corales del Pacífico occidental (Reyes-Bonilla y López-Pérez 1998). Sin embargo, el árbol muestra que las Islas Marías son clave como sitio intermedio de dispersión hacia el norte para propágulos ya establecidos en la costa continental.
En conclusión, las asociaciones de corales de las Islas Marías constan de 20 especies. En 1997 la región no presentaba arrecifes en sentido construccional pero tenía elevada cobertura coralina sobre el fondo, especialmente en la Isla María Madre. Las comunidades de las islas evidenciaron una clara zonación con Pocillopora en agua somera y Porites, Pavona y Psammocora en sitios profundos. Finalmente, desde la perspectiva biogeográfica la entrada del Golfo de California representa una región clave para la inmigración de especies desde el Pacífico oeste, y las Islas Marías parecen jugar un papel preponderante para la dispersión de esas especies hacia el norte. Esperamos que la información aquí generada sea útil para ayudar a preservar uno de los últimos sitios en estado prístino de conservación en el Pacífico mexicano.
Agradecimientos
Se agradece el patrocinio para la visita a las islas por el Instituto Nacional de la Pesca, del Gobierno Federal y del Estado de Nayarit. También a O Ramírez, F Uribe, N Ceseña, la Cooperativa Pesquera de Islas Marías, y a la Dirección de la Colonia Penal Federal de Islas Marías, por la ayuda durante el trabajo de campo. El estudio también recibió apoyo por parte del proyecto SEMARNAT-CONACYT-CONABIO AS007 (responsable: LE Calderón Aguilera, CICESE, Ensenada). Los curadores de las colecciones visitadas (R van Syoc, CASIZ; D Haasl, UCMP; E Hochberg, SBMNH; KD Johnson, LACM) otorgaron todas las facilidades para la revisión de ejemplares en sus instituciones. VH Beltrán (James Cook University, Townsville), J Ketchum (University of California, Davis), seis árbitros y el editor de la revista hicieron sugerencias relevantes para mejorar el contenido del manuscrito.
Referencias
Carriquiry JD, Reyes-Bonilla H. 1997. Estructura de la comunidad y distribución geográfica de los arrecifes coralinos de Nayarit. Cienc. Mar. 23: 227-248. [ Links ]
Carriquiry JD, Cupul-Magaña A, Rodríguez-Zaragoza F, Medina-Rosas P. 2001. Coral bleaching and mortality in the Mexican Pacific during the 1997-98 El Niño, and prediction from a remote sensing approach. Bull. Mar. Sci. 69: 237-249. [ Links ]
Clarke KR, Warwick RM. 2001. Change in Marine Communities. 2nd ed. Primer-E, Plymouth, 144 pp. [ Links ]
Crisci JV, Katinas L, Posadas P. 2003. Historical Biogeography. Harvard Univ. Press, Boston, 264 pp. [ Links ]
Durham JW, Barnard JL. 1952. Stony corals of the eastern Pacific collected by the Velero III and Velero IV. Allan Hancock Pac. Exped. 16: 1-110. [ Links ]
English S, Wilkinson C, Baker V. 1997. Survey Manual for Tropical Marine Resources. AIMS, Townsville, 312 pp. [ Links ]
García E. 1989. Climas. Atlas Nacional de México. Instituto de Geografía, Univ. Nacional Autónoma de México, IV.4.10. [ Links ]
Glynn PW. 2000. Effects of the 1997-98 El Niño Southern Oscillation on eastern Pacific corals and coral reefs: An overview. Proc. 9th Int. Coral Reef Symp., Bali, 2: 1169-1174. [ Links ]
Glynn PW, Leyte-Morales GE. 1997. Coral reefs of Huatulco, Mexico: Reef development in upwelling Gulf of Tehuantepec. Rev. Biol. Trop. 45: 1033-1048. [ Links ]
Glynn PW, Ault JS. 2000. A biogeographic analysis and review of the far eastern Pacific coral reef region. Coral Reefs 19: 1-23. [ Links ]
Guzmán ML. 1959. Islas Marías. Compañía General de Ediciones, México, 121 pp. [ Links ]
Guzmán HM, Cortés J. 1993. Arrecifes coralinos del Pacífico oriental tropical: Revisión y perspectivas. Rev. Biol. Trop. 41: 535-557. [ Links ]
Iglesias-Prieto R, Beltrán VH, La Jeunesse T, Reyes-Bonilla H, Thomé PE. 2004. The presence of specific algal symbionts explains the vertical distribution patterns of hermatypic corals. Proc. R. Soc. London, Ser. B, 271: 1757-1763. [ Links ]
Ketchum JT, Reyes-Bonilla H. 2001. Taxonomía y distribución de los corales hermatípicos (Scleractinia) del Archipiélago de Revillagigedo, Pacífico de México. Rev. Biol. Trop. 49: 803-848. [ Links ]
Knowlton N. 1993. Sibling species in the sea. Annu. Rev. Ecol. Syst. 24: 189-216. [ Links ]
Quinn GP, Keough MJ. 2002. Experimental design and data analysis for biologists. Cambridge Univ. Press, Cambridge, 556 pp. [ Links ]
Revueltas J. 1988. Los Muros de Agua. Era, México, 65 pp. [ Links ]
Reyes-Bonilla H. 1993. Biogeografía y ecología de los corales hermatípicos (Anthozoa: Scleractinia) del Pacífico de México. En: Salazar-Vallejo SI, González NE (eds.), Biodiversidad Marina y Costera de México. CONABIO/CIQRO, Chetumal, pp. 207-222. [ Links ]
Reyes-Bonilla H. 2002. Checklist of valid names and synonyms of stony corals (Anthozoa: Scleractinia) of the eastern Pacific Ocean. J. Nat. Hist. 36: 1-13. [ Links ]
Reyes-Bonilla H. 2003. Coral reefs of the Pacific coast of Mexico. In: Cortés J (ed.), Latin American Coral Reefs. Elsevier, Amsterdam, pp. 331-349. [ Links ]
Reyes-Bonilla H, López-Pérez A. 1998. Biogeografía de los corales pétreos (Scleractinia) del Pacífico de México. Cienc. Mar. 24: 211-224. [ Links ]
Reyes-Bonilla H, Pérez-Vivar TL, Ketchum-Mejía JT. 1999. Distribución geográfica y depredación de Porites lobata (Anthozoa: Scleractinia) en la costa occidental de México. Rev. Biol. Trop. 47: 273-279. [ Links ]
Reyes-Bonilla H, Carriquiry JD, Leyte-Morales GE, Cupul-Magaña AL. 2002. Effects of the El Niño-Southern Oscillation and the anti-El Niño event (1997-1999) on coral reefs of the western coast of Mexico. Coral Reefs 21: 368-372. [ Links ]
Reyes-Bonilla H, Calderón-Aguilera LE, Cruz-Piñón G, Medina-Rosas P, López-Pérez RA, Herrero-Pérezrul MD, Leyte-Morales GE, Cupul-Magaña AL, Carriquiry-Beltrán JD. 2005. Atlas de los corales pétreos (Scleractinia) del Pacífico mexicano. CICESE/CONABIO/CONACYT/UABCS/Universidad de Guadalajara/Universidad del Mar, 124 pp. [ Links ]
Robertson DR, Allen GR. 2002. Shorefishes of the tropical eastern Pacific. CD-ROM. Smithsonian Tropical Research Institute. [ Links ]
Small M. 1998. Ten days in May: A trip to las Islas Tres Marías. Festivus 30: 75-82. [ Links ]
Spalding M, Ravilious C, Green EP. 2000. World Atlas of Coral Reefs. Univ. California Press, Berkeley, 416 pp. [ Links ]
Squires DF. 1959. Corals and coral reefs in the Gulf of California. Bull. Am. Mus. Nat. Hist. 118: 367-432. [ Links ]
Veron JEN. 1986. Corals of Australia and the Indo-Pacific. Angus and Robertson, Sydney, 580 pp. [ Links ]
Veron JEN. 2000. Corals of the World. Vols. 1-3. Australian Institute of Marine Science, Townsville, 1382 pp. [ Links ]
Wellington GM. 1982. Depth zonation of corals in the Gulf of Panamá: Control and facilitation by resident reef fishes. Ecol. Monogr. 52: 223-241. [ Links ]
Wells JW. 1983. Annotated list of the scleractinian corals of the Galápagos Islands. In: Glynn PW, Wellington GM (eds.), Corals and Coral Reefs of the Galápagos Islands. Univ. California Press, Berkeley, pp. 209-292. [ Links ]
Wells SM. 1988. Coral Reefs of the World. Vol. 1. Atlantic and Eastern Pacific. IUCN, Cambridge, 373 pp. [ Links ]
WOA, World Ocean Atlas. 2001. http://www.nodc.noaa.gov/OC5/WOA01/pr_woa01.html. [ Links ]
Vidal-Zepeda R. 1989. Precipitación. Atlas Nacional de México. Instituto de Geografía, Univ. Nacional Autónoma de México, IV.4.4. [ Links ]

