










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCiencias marinas
versión impresa ISSN 0185-3880
Cienc. mar vol.32 no.1a Ensenada mar. 2006
Nota de Investigación
Ecología de postlarvas de camarón azul (Litopenaeus stylirostris) y camarón café (Farfantepenaeus californiensis) en el Estuario del Río Colorado
Postlarval ecology of the blue shrimp (Litopenaeus stylirostris) and brown shrimp (Farfantepenaeus californiensis) in the Colorado River Estuary
RA Ramírez-Rojo y EA Aragón-Noriega*
Centro de Investigaciones Biológicas del Noroeste (CIBNOR), Unidad Sonora, Km 2.35 Camino al Tular, Estero Bacochibampo, Guaymas 85454, Sonora, México. * E-mail: aaragon04@cibnor.mx
Recibido en septiembre de 2003.
Recibido en su forma actual en septiembre de 2004.
Aceptado en julio de 2005.
Resumen
Se describe el uso del hábitat por las postlarvas de camarones peneidos que ingresan al delta del Río Colorado. Se seleccionó una localidad en el Canal Santa Clara, dentro de la zona núcleo de la Reserva de la Biosfera Alto Golfo de California y Delta del Río Colorado, donde se realizaron recolecciones durante tres días consecutivos de cada marea viva de mayo a octubre de 2000. Se ancló una red con luz de malla de 505 |am que se mantuvo en el agua 10 minutos cuando la altura de marea se encontraba a 1 m sobre el nivel del canal, tanto en el flujo como en reflujo. Se capturaron postlarvas de camarón azul Litopenaeus stylirostris y camarón café Farfantepenaeus californiensis con proporciones de 80% y 20%, respectivamente. La abundancia de postlarvas de camarón azul fue significativamente mayor en el flujo que en reflujo en mayo, junio y septiembre, pero significativamente mayor en el reflujo que en el flujo en julio y agosto. Las tallas de las postlarvas de camarón azul fueron significativamente mayores en el reflujo que en el flujo. En las postlarvas de camarón café no se encontró diferencia entre la abundancia del flujo y el reflujo. Se concluye que el delta del Río Colorado, es un área de crianza sólo de camarón azul y no para camarón café.
Palabras clave: postlarvas, camarón, Río Colorado, Litopenaeus stylirostris, Farfantepenaeus californiensis.
Abstract
The study describes habitat use by penaeid shrimp larvae that enter the Colorado River Delta. A location was chosen in the Santa Clara Channel in the core zone of the Upper Gulf of California and Colorado River Delta Biosphere Reserve, where collections were done during three consecutive days of each spring tide from May to October 2000. A 505-| m net was anchored and left in the water for 10 minutes when the height of the tide was 1 m above the level of the channel, during both flood tide and ebb tide. Blue shrimp (Litopenaeus stylirostris) and brown shrimp (Farfantepenaeus californiensis) postlarvae were caught in proportions of 80% and 20%, respectively. The postlarval abundance of blue shrimp was significatively greater during flood tide than during ebb tide in May, June and September, but significatively greater during ebb tide than during flood tide in July and August. The blue shrimp postlarval sizes were significatively greater during ebb tide than during flood tide. No differences in the abundance of brown shrimp postlarvae were found between flood and ebb tides. It is concluded that the Colorado River Delta is a breeding area for blue shrimp, but not for brown shrimp.
Key words: postlarvae, shrimp, Colorado River, Litopenaeus stylirostris, Farfantepenaeus californiensis.
Introducción
El camarón café Farfantepenaeus californiensis (Holmes 1900) y el azul Litopenaeus stylirostris (Stimpson 1874) se distribuyen en la región zoogeográfica del Pacífico oriental, desde Estados Unidos hasta Perú, incluyendo las islas Galápagos y el Golfo de California, el primero, y desde el Alto Golfo de California hasta Perú, el segundo (Hendrickx 1996).
Todos los miembros conocidos de la familia Penaeidae tienen un ciclo de vida similar: larvas planctónicas con estadios nauplius, protozoea, mysis y postlarva, seguidos de estadios juvenil y adulto (Dall et al. 1990). La gran diferencia entre géneros consiste en los hábitat preferidos por las postlarvas, juveniles y adultos, los cuales pueden ser estuarios y lagunas costeras, la plataforma continental o el océano.
Dall et al. (1990) mencionan que la base para clasificar el ciclo de vida de los peneidos es el ambiente en el cual desovan los adultos y el que usan las postlarvas para asentarse, distinguiendo cuatro tipos de ciclo de vida con la aclaración de que puede haber traslapo entre ellos.
Para el caso de este estudio es necesaria la descripción de los ciclos de vida "tipo 2" y "tipo 3". En el ciclo de vida tipo 2 la vida adulta y la fase reproductora transcurren en el hábitat oceánico (entre mar adentro y la plataforma continental). Los huevos son expulsados por la hembra al medio marino donde transcurren los cuatro estadios larvales. Las postlarvas de las especies que presentan este ciclo de vida utilizan ambientes estuarinos para asentarse, pasan por el estadio juvenil y migran al océano en su estadio preadulto. En el ciclo de vida tipo 3 los adultos y larvas ocupan hábitat netamente oceánicos y las postlarvas y juveniles costeros, pero no estuarinos (Dall et al. 1990). Estos mismos autores mencionan que la mayoría de las especies del género Penaeus y Metapenaeus presentan ciclo de vida tipo 2.
Los miembros del género Penaeus fueron divididos por Pérez-Farfante (1969) en cuatro subgéneros: Litopenaeus, Penaeus, Melicertus y Fenneropenaeus. Posteriormente Burukovsky (1972) clasificó como Farfantepenaeus a Melicertus. Pérez-Farfante y Kensley (1997) elevaron a rango de género los subgéneros Litopenaeus y Farfantepenaeus. Considerando la nueva nomenclatura, las especies que presentan un ciclo de vida tipo 2 están incluidas en los géneros Litopenaeus y Farfantepenaeus.
Para facilitar comparaciones, García (1985) separó los camarones de género Penaeus en blancos (Litopenaeus; ciclo de vida tipo 2) y oscuros (Farfantepenaeus; ciclo de vida tipo 3). Hacemos notar que esta clasificación fue hecha antes de Pérez-Farfante y Kensley (1997). García (1985) incluyó a L. stylirostris entre los camarones blancos y a F. californiensis entre los oscuros. Después de esto, los ciclos de vida de las especies nunca más han sido cuestionados.
En una revisión bibliográfica de la presencia de camarón café y azul en las lagunas costeras del litoral mexicano encontramos que en Huizache-Caimanero, Sinaloa, las postlarvas penetran por las bocas del sistema, pero posteriormente no se capturan los juveniles en el interior (Poli 1983, Del Valle 1989). En el interior de la laguna costera Mar Muerto, Oaxaca, se han encontrado juveniles de camarón café (Medina-Reyna et al. 2001) y por la boca El Oro del sistema Corralero-Alotengo, Oaxaca, se ha registrado la entrada de postlarvas de esta especie (Sánchez-Meraz y Martínez-Vega 2000). En las lagunas costeras de Sonora (esteros El Soldado y La Cruz y Agiabampo) durante todo el año se han encontrado juveniles de camarón café mayores a 100 mm de longitud total. En Estero La Cruz y Agiabampo se registró ingreso de postlarvas (Loesch 1980, Grijalva-Chon y Barraza-Guardado 1992, Manzano-Sarabia 2003). Con lo anterior se puede concluir que el camarón café del Pacífico mexicano presenta un traslapo entre los ciclos de vida tipo 2 y 3, lo que aparentemente sucede de manera diferenciada con respecto a la latitud: en algunos sitios se presenta como tipo 2 y en otros como tipo 3. Es importante definir el ciclo de vida de esta especie en una zona determinada para establecer su área de crianza en el sitio de estudio.
El Estuario del Río Colorado (ERC) se encuentra localizado en la parte más norteña del Golfo de California, dentro de la zona núcleo de la Reserva de la Biosfera del Alto Golfo de California y Delta del Río Colorado (RBAGCDRC). Esta zona es considerada de gran importancia para la crianza de camarón y otras especies que dependen de los estuarios (Cudney-Bueno y Turk-Boyer 1998, Galindo-Bect et al. 2000). Snyder-Conn y Brusca (1975) concluyeron que no existe una descripción adecuada del ciclo de vida, ecología y patrón migratorio del camarón en el Alto Golfo de California. Desafortunadamente esta conclusión es aún vigente a pesar de que los esfuerzos de investigación sobre postlarvas de camarón se han incrementado en los últimos diez años. A partir de estos estudios se han identificado bien las zonas de desove de los camarones (Aragón-Noriega 2000) y su periodo de mayor reclutamiento (Aragón-Noriega y García-Juárez 2002), así como las posibles rutas de transporte que siguen las larvas desde la zona de desove hasta el ERC (Calderón-Aguilera et al. 2003, Galindo-Bect 2003).
Actualmente dos tópicos se manifiestan como importantes en el estudio de postlarvas de camarón en el ERC: (1) describir el tipo de ciclo de vida de las especies que allí habitan y (2) aportar información cuantitativa de las especies que hacen uso del ERC como área de crianza. Reconociendo el valor que cada aportación representa para el acervo de conocimientos de esta importante área ecológica, el objetivo de este trabajo fue presentar datos cuantitativos sobre el uso que hacen las postlarvas de camarón azul (L. stylirostris) y camarón café (F. californiensis) del ERC.
Material y métodos
Se realizaron muestreos durante las mareas vivas del 30 de mayo al 12 de octubre de 2000 en el Canal Santa Clara, localizado en los límites de la zona núcleo de la RBAGCDRC (fig. 1). Las recolecciones se hicieron cada 15 días, durante tres días, recolectando solamente a una altura de marea de 1 m durante el flujo y el reflujo de la marea. Esto es que un equipo permaneció en el lugar de muestreo durante tres días continuos. En el Alto Golfo se presenta una marea semi-diurna (dos flujos y dos reflujos al día), por lo que se tienen de cuatro a seis muestreos tanto en flujo como en reflujo en cada campaña.
Se utilizó una red de plancton con un tamaño de malla de 505 | m y boca circular de 62 cm de diámetro equipada con un flujómetro mecánico y atada a una embarcación menor que permaneció anclada en el Canal Santa Clara. La red permaneció en el agua durante 10 minutos cuando la marea tenía una altura de 1 m sobre el fondo del canal. Las muestras recolectadas se depositaron en frascos de 500 mL y se fijaron con formol al 4% neutralizado con borato de sodio, y posteriormente se preservaron en alcohol isopropílico al 70%.
De las muestras recolectadas en el campo se separaron sólo las postlarvas de camarón y el resto se regresó al frasco de muestra. Una vez separadas se contaron en su totalidad y se tomó una submuestra de 100 organismos para obtener la longitud total (del margen anterior del rostrum al extremo posterior del telson) de las postlarvas con un micrómetro montado en un microscopio compuesto utilizando los objetivos 10x y 20x. La identificación se realizó siguiendo las claves propuestas por Mair (1979) y Calderón-Pérez et al. (1989a, 1989b). Se utilizaron también los organismos de referencia cultivados por Aragón-Noriega (2000).
Se hizo un análisis de varianza de dos factores en el que la variable considerada independiente fue la abundancia. Como factor I se consideró las especies y como factor II el estado de la marea (flujo vs. reflujo). De igual forma, se hizo un análisis considerando la longitud total como la variable independiente. Posteriormente la longitud de las postlarvas de camarón azul se comparó de acuerdo al estado de la marea y el ciclo diurno (noche vs. día). El análisis de varianza se realizó con un 95% de confiabilidad y cuando se encontraron diferencias significativas se aplicó la prueba Tukey (Zar, 1996).
Resultados
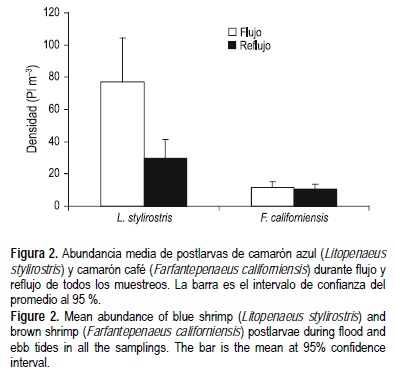
A lo largo del periodo de muestreo se encontraron sólo dos especies de camarones peneidos, camarón azul y camarón café, siendo más abundantes en el ERC las postlarvas (Pl) de camarón azul (F(1,120) = 178; P < 0.0001) que las de camarón café. La abundancia media (± intervalo de confianza al 95% de significancia) de camarón azul fue de 53 (±16) Pl m-3, mientras que la de camarón café fue de 10 (±3) Pl m-3 (fig. 2). En la figura 2 se sintetiza la abundancia de postlarvas de ambas especies de acuerdo al estado de la marea (flujo o reflujo), siendo mayor la abundancia de postlarvas de camarón azul (F(1,120)= 53; P < 0.0001) en el flujo (77 ± 27 Pl m-3) que en el reflujo (30 ± 11 Pl m-3), mientras que las postlarvas de camarón café no presentaron diferencia significativa entre flujo (12 ± 3 Pl m-3) y reflujo (11 ± 3 Pl m-3).
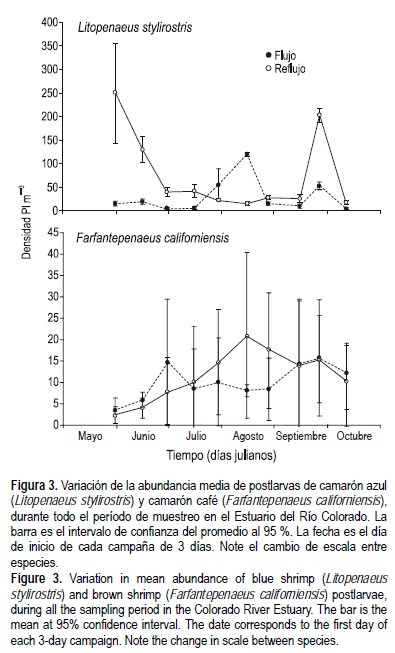
El mayor reclutamiento se presentó en mayo-junio y septiembre (fig. 3). En todas las campañas de muestreo, excepto en julio y agosto, las postlarvas de camarón azul fueron más abundantes durante el flujo que en el reflujo, mientras que las de camarón café no presentaron diferencias entre flujo y reflujo en todo el periodo de muestreo.
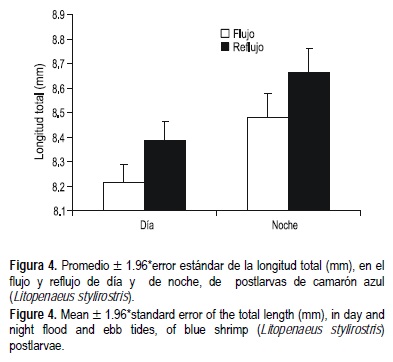
Durante todo el periodo de recolección el tamaño medio de las postlarvas del camarón azul fue de 8 ± 0.79 mm, mientras que el de las postlarvas de camarón café fue de 9 ± 0.87 mm. En la figura 4 se presentan las tallas medias de las postlarvas capturadas en el ERC durante el flujo, el reflujo, el día y la noche, a lo largo de todo el periodo de muestreo. Las variaciones en talla evidencian un patrón de comportamiento de las postlarvas para entrar y/o salir del ERC: las postlarvas más grandes se mueven por la noche y las más pequeñas lo hacen en el día.
Discusión
De acuerdo con la definición de ciclos de vida propuesta por Dall et al. (1990), en el ERC el camarón azul presenta un ciclo de vida tipo 2 y el camarón café presenta uno tipo 3, por lo que se deduce que el camarón azul utiliza esta zona como área de crianza, mientras que el camarón cafe presenta menor posibilidad de usar el ERC con estos fines.
En estudios anteriores se ha mencionado que en junio y julio ocurre el mayor reclutamiento de camarón al área litoral (Aragón-Noriega y García-Juárez 2002) y al ERC (Ramírez-Rojo 2001). En la zona litoral del Golfo de Santa Clara, Calderón-Aguilera et al. (2002) encontraron una abundancia media similar de postlarvas de camarón café y azul que varió de 1 Pl m-3 en 1995 hasta 151 Pl m-3 en 1996. En este estudio se encontró una abundancia media de 50 Pl m-3 de L. stylirostris a lo largo de todo el periodo de recolección, mientras que la abundancia media de F. californiensis fue de 10 Pl m-3 en el ERC. Esto puede indicar, en primer lugar, que la abundancia varía anualmente, pero al no encontrar diferencias en la abundancia de cada especie para un mismo sitio y año (Calderón-Aguilera et al. 2002) también sugiere que la diferencia puede ser además entre sitios. Si en la zona marina no hay diferencias en la abundancia de postlarvas y en el canal del estuario éstas sí se presentan, es posible que las postlarvas (en este caso de camarón azul) viajen hasta ese sitio para asentarse y completar su ciclo de vida. La presencia de postlarvas de camarón café en la zona estuarina posiblemente sea causada por una advección pasiva, lo que se deduce de que su abundancia durante el flujo no fue significativamente diferente a la del reflujo en un una misma marea viva a lo largo del periodo de muestreo.
Aplicando el mismo criterio de las diferencias en la abundancia de las postlarvas de camarón azul durante el flujo y el reflujo, encontramos que las diferencias sí fueron significativamente mayores en los flujos por lo que se puede sugerir que el ERC es un lugar de crianza para el camarón azul, pero no para el camarón café.
Ahora bien, todo lo anterior es válido si la ruta de entrada y salida de las postlarvas es la misma, esto es, que el sitio seleccionado para la recolección realmente refleja el hecho de que las postlarvas entran y salen en las cantidades aquí encontradas y no presentan una ruta de entrada diferente a la de salida que pudiera sesgar los datos.
Otro aspecto de particular importancia fueron las longitudes de camarón azul encontradas tanto en el flujo diurno y nocturno como en el reflujo diurno y nocturno. En los resultados se observa que las postlarvas de tallas grandes salen del estuario hacia el mar durante la noche. En cambio, las postlarvas de menor talla lo hacen durante el día. No se tienen elementos para asegurar que ésta sea una estrategia de defensa ante depredadores o que la diferencias encontradas en las tallas con relación a los ritmos de marea (flujo, reflujo) y diurno (día, noche) sea un simple hecho casual.
Las diferencias en longitud encontradas en las postlarvas durante flujo y reflujo pueden ser una evidencia más para afirmar que las postlarvas de camarón azul utilizan el ERC como zona de crianza, al igual que evidenciaron que las postlarvas de camarón café no lo hacen dadas las tallas similares encontradas en sus postlarvas durante el flujo y el reflujo. Las postlarvas de camarón azul entran pequeñas (8 mm de longitud total) al ERC y salen más grandes (9 mm de longitud total), es decir, que salen del estuario después de un cierto periodo de crecimiento.
Las diferencias de talla encontradas entre las postlarvas de ambas especies es otro punto importante a discusión, dado que las tallas de camarón café encontradas fueron sistemáticamente mayores que las de camarón azul, a diferencia de lo reportado hasta el momento por otros autores en la zona (Castillo-Moreno 1999, Aragón-Noriega y Calderón-Aguilera 2001), quienes encontraron tallas mayores de camarón azul que de camarón café. Cabe señalar que estos autores presentan datos de la zona de rompiente de playa, por lo que es posible que las postlarvas capturadas durante los muestreos del presente estudio, que corresponden al área del estuario, lleguen a éste una vez que han alcanzado tallas más grandes. Como los datos de tallas referidos por esos autores fueron obtenidos en años diferentes al de este estudio, es posible que la diferencia en tallas se deba a una diferencia en los periodos de desove de las especies en cada año en particular. Esto es que, particularmente en 2000, el camarón azul haya desovado después que el camarón café, cuya tasa de crecimiento es menor (Aragón-Noriega y Calderón-Aguilera 2001). Si el desove de ambas especies se hubiera presentado al mismo tiempo, las postlarvas de camarón azul serían mayores por tener mayor tasa de crecimiento.
Como conclusión del presente estudio se establece que el ERC es un área de crianza sólamente para el camarón azul y no para el camarón café, mientras que en el área del delta del Río Colorado el camarón café tiene un ciclo de vida tipo 3 y el camarón azul tiene un ciclo tipo 2.
Agradecimientos
El presente estudio fue financiado con fondos regulares (EP 1.2 y AYCG 20) del Centro de Investigaciones Biológicas del Noroeste (CIBNOR). Agradecemos a Edgar Alcántara-Razo su apoyo técnico.
Referencias
Aragón-Noriega EA. 2000. Ecología del reclutamiento del camarón azul Litopenaeus stylirostris (Stimpson, 1871) en el Alto Golfo de California. Tesis de doctorado, Centro de Investigación Científica y de Educación Superior de Ensenada, Baja California, México, 117 pp. [ Links ]
Aragón-Noriega EA, Calderón-Aguilera LE. 2001. Age and growth of shrimp postlarvae in the Upper Gulf of California. Aqua, J. Ichthyol. Aquat. Biol. 4: 99-104. [ Links ]
Aragón-Noriega EA, García-Juárez AR. 2002. Reclutamiento de postlarvas de camarón azul Litopenaeus stylirostris (Stimpson, 1871) a condiciones antiestuarinas provocadas por actividades antropogénicas. Hidrobiológica 12: 37-46. [ Links ]
Burukovsky RN. 1972. Nekotorye voprosy sistematiki i rasprostraneniya krevetok roda Penaeus. Rybokhozyaistvennye issledovaniya v Atlanticheskom okeane. (Some problems of the systematics and distribution of shrimps of the genus Penaeus.) Trudy AtlantNIRO, Kaliningrad, 2: 3-21. [ Links ]
Calderón-Aguilera LE, Aragón-Noriega EA, Licón HA, Castillo-Moreno G, Maciel-Gómez A. 2002. Abundance and composition of penaeid postlarvae in the Upper Gulf of California. In: Hendrickx ME (ed.), Contributions to the Study of East Pacific Crustaceans. Instituto de Ciencias del Mar y Limnología, UNAM, pp. 281-291. [ Links ]
Calderón-Aguilera LE, Marinone SG, Aragón-Noriega EA. 2003. Influence of oceanographic processes on the early life stages of the blue shrimp (Litopenaeus stylirostris) in the Upper Gulf of California. J. Mar. Syst. 39: 117-128. [ Links ]
Calderón-Pérez JA, Macías-Regalado E, Abreu-Grobois FA, Rendón-Rodríguez S. 1989a. Antennular flagella: A useful character for distinguishing subgenera among postlarval shrimp of the genus Penaeus (Decapoda) from the Gulf of California. J. Crust. Biol. 9: 482-491. [ Links ]
Calderón-Pérez JA, Macías-Regalado E, Rendón-Rodríguez S. 1989b. Clave de identificación para los estadios de postlarva y primeros juveniles de camarón del género Penaeus (Crustacea: Decapoda) del Golfo de California, México. Cienc. Mar. 15(3): 57-70. [ Links ]
Castillo-Moreno G. 1999. Efecto de algunas variables sobre la abundancia relativa de postlarvas de camarón del genero Litopenaeus y Farfantepenaeus (Crustacea: Decapoda) en el Alto Golfo de California. Tesis de maestría, Centro de Investigación Científica y de Educación Superior de Ensenada, Baja California, México, 74 pp. [ Links ]
Cudney-Bueno R, Turk-Boyer PJ. 1998. Pescando entre Mareas del Alto Golfo de California. Centro Intercultural de Estudios de Desiertos 7 Océanos, Puerto Peñasco, Sonora, México, 166 pp. [ Links ]
Dall W, Hill BJ, Rothlisberg PC, Staples DJ. 1990. The biology of the Penaeidae. In: Blaxter JHS, Southward AJ (eds.), Advances in Marine Biology. Vol. 27. Academic Press, London, pp. 1-189. [ Links ]
Del Valle I. 1989. Estrategia de explotación y producción en una laguna costera en México. Tesis de doctorado, Universidad Poliotécnica de Cataluña, Barcelona, España, 266 pp. [ Links ]
Galindo-Bect MS. 2003. Larvas y postlarvas de camarones peneidos en el Alto Golfo de California y capturas de camarón con relación al flujo del Río Colorado. Tesis de doctorado, Universidad Autónoma de Baja California, Ensenada, México, 146 pp. [ Links ]
Galindo-Bect MS, Glenn EP, Page HM, Fitzsimmons K, Galindo-Bect LA, Hernández-Ayón JM, Petty RL, García-Hernández J, Moore D. 2000. Penaeid shrimp landings in the Upper Gulf of California in relation to Colorado River freshwater discharge. Fish. Bull. 98: 222-225. [ Links ]
García S. 1985. Reproduction, stock assessment models and population parameters in exploited penaeid shrimp populations. In: Rothlisberg PCB, Hill J, Staples DJ (eds.), Second Australian National Prawn Seminar, NOS2. Cleveland, Australia, pp. 139-158. [ Links ]
Grijalva-Chon JM, Barraza-Guardado RH. 1992. Distribución y abundancia de las postlarvas y juveniles de los camarones del género Penaeus en Bahía Kino y laguna de La Cruz, Sonora, México. Cienc. Mar. 18(3): 153-169. [ Links ]
Hendrickx ME. 1996. Los Camarones Penaeoidea Bentónicos (Crustacea: Decapoda: Dendrobranchiata) del Pacífico Mexicano. ICMyL-UNAM-CONABIO, México, 147 pp. [ Links ]
Loesch H. 1980. Some ecological observations on slow-swimming nekton with emphasis on penaeid shrimp in a small Mexican west coast estuary. Anal. Centro Cienc. Mar Limnol. Univ. Nac. Autón. Mex. 7: 15-26. [ Links ]
Mair JM. 1979. The identification of postlarvae of four species of Penaeus (Crustacea: Decapoda) from the Pacific Coast of Mexico. J. Zool. 188: 347-351. [ Links ]
Manzano-Sarabia MM. 2003. Distribución y abundancia de camarón café Farfantepenaeus californiensis en el sistema lagunar de Agiabampo, Sonora-Sinaloa, México. Tesis de maestría, Centro de Investigaciones Biológicas del Noroeste, La Paz, Baja California Sur, 76 pp. [ Links ]
Medina-Reyna CE, Sánchez-Meraz B, Chávez-Otañez LA, Zayas-Alvarez A. 2001. ¿Qué tanto afecta el "Norte" a las postlarvas y juveniles de camarón en la laguna Mar Muerto. Ciencia y Mar, 5(14): 3-12. [ Links ]
Pérez-Farfante I. 1969. Western Atlantic shrimp of the genus Penaeus. Fish. Bull. 67: 461-591. [ Links ]
Pérez-Farfante I., Kensley B. 1997. Penaeoid and segestoid shrimps and prawns of the world. Keys and diagnoses for the families and genera. Mem. Mus. Natl. Hist. Nat. (France) 175: 1-233. [ Links ]
Poli CR. 1983. Patrón de inmigración de postlarvas de Penaeus spp. (Crustacea: Decapoda: Penaidae) en la boca del Río Baluarte, Sinaloa, México. Tesis de doctorado, UACP/PCCH, UNAM, 182 pp. [ Links ]
Ramírez-Rojo RA. 2001. Ecología de postlarvas de camarón azul (Litopenaeus stylirostris) y camarón café (Farfantepenaeus californiensis) en el estuario del Río Colorado. Tesis de licenciatura, ITMAR 03, Guaymas, Sonora, 47 pp. [ Links ]
Sánchez-Meraz B, Martínez-Vega JA. 2000. Inmigración de postlarvas de camarón Litopenaeus sp. y Farfantepenaeus sp. a través de la Boca el Oro del sistema lagunar Corralero-Alotengo, Oaxaca.Ciencia y Mar 4(12): 29-46. [ Links ]
Snyder-Conn E, Brusca RC. 1975. Shrimp population dynamics and fishery impact in the northern Gulf of California. 1967-1968. Cienc. Mar., 2(2): 54-67. [ Links ]
Zar JH. 1996. Biostatistical Analysis. 3rd ed. Prentice Hall, New Jersey, 662 pp. [ Links ]

