










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkCiencias marinas
versão impressa ISSN 0185-3880
Cienc. mar vol.31 no.4 Ensenada Dez. 2005
Artículos
Estructura poblacional y distribución espacial de los moluscos gasterópodos Osilinus atrata y Osilinus sauciatus en el intermareal rocoso de las Islas Canarias (Atlántico centro-oriental)
Population structure and spatial distribution of the gastropod molluscs Osilinus atrata and Osilinus sauciatus in the rocky intertidal zone of the Canary Islands (Central East Atlantic)
R. Ramírez1*, F. Tuya1, P. Sánchez-Jerez2, C. Fernández-Gil1, O. Bergasa1, R.J. Haroun1 y J.J. Hernández-Brito1
1 BIOGES Faculty of Marine Sciences Campus de Tafira University of Las Palmas de G.C E-35017, Las Palmas, Canary Islands, Spain. * E-mail: ruben.ramirez101@doctorandos.ulpgc.es
2 Marine Biology Lab University of Alicante POB 99 E-03080, Alicante, Spain.
Recibido en julio de 2004;
aceptado en agosto.
Resumen
Por medio de un diseño de muestreo jerárquico se estudió la estructura poblacional y los patrones espaciales de variabilidad horizontal (diferencias entre las islas, las localidades dentro de las islas y los sitios dentro de localidades) y vertical (diferencias entre bandas de la zona intermareal) de los moluscos gasterópodos Osilinus atrata y O. sauciatus a lo largo del intermareal rocoso de las Islas Canarias. Se observaron diferencias en los patrones espaciales de distribución, abundancia y tamaño para ambas especies. Osilinus atrata fue más abundante que O. sauciatus en todo el Archipiélago Canario. Su abundancia varió entre las islas, presentando una distribución heterogénea entre las bandas intermareales de una isla a otra, si bien fue más frecuente en las bandas intermedia y superior. Osilinus sauciatus sólo se encontró en las islas orientales y no exhibió diferencias significativas entre las bandas, a pesar de que 91.35% de los individuos aparecieron en la banda superior y el resto en la banda intermedia. Osilinus sauciatus mostró una talla mayor que las de O. atrata. A su vez, ambas especies presentaron en general las mayores tallas medias en la banda superior.
Palabras clave: gasterópodos, Osilinus atrata, Osilinus sauciatus, estructura poblacional, intermareal rocoso.
Abstract
A hierarchical sampling design was used to study the population structure and the horizontal (differences among islands, locations within islands and sites within locations) and vertical (differences among intertidal bands) distribution patterns of the gastropod molluscs Osilinus atrata and O. sauciatus along the rocky intertidal zone of the Canary Islands. Differences in the spatial patterns of abundance, size and distribution were recorded for both species. Osilinus atrata was more abundant than O. sauciatus throughout the archipelago. Its abundance varied among islands and it presented a heterogeneous distribution in the bands from one island to another, though it was more frequent in the middle and high bands. Osilinus sauciatus was only found at the eastern islands and did not exhibit significant differences between the bands, even though 91.35% of the individuals appeared in the high band and the rest in the middle band. On the other hand, O. sauciatus had a larger mean size than O. atrata, but both species presented the largest mean sizes in the high band.
Key words: gastropods, Osilinus atrata, Osilinus sauciatus, population structure, rocky intertidal zone.
Introducción
La zona intermareal posee condiciones muy extremas que condicionan la colonización por diferentes especies y su distribución espacial. En pocos metros a lo largo de su eje vertical, las condiciones ambientales (e.g., temperatura, salinidad, oleaje e irradiación) varían desde un medio completamente acuático hasta uno completamente terrestre, originando gradientes muy acusados (Bertness, 1999; Harley y Helmuth, 2003) y haciendo del intermareal un hábitat heterogéneo. De este modo, los organismos se distribuyen en la zona interma-real presentando una zonación (Stephenson y Stephenson, 1949) que no puede explicar por sí sola la variabilidad en los modelos de distribución espacial de las poblaciones (Underwood y Chapman, 1996; Menconi et al., 1999; Benedetti-Cecchi, 2001).
De los grupos más abundantes, los gasterópodos son los mejor representados en la zona intermareal. En las Islas Canarias destacan varias especies del género Patella (Navarro et al., 2005), los burgados (Osilinus spp.) y la carnadilla (Stramonita haemastoma). No existen trabajos para el conjunto del archipiélago sobre los burgados; los que hay no son específicos y además se limitan a una isla (Núñez, 1995). Las especies que se dan cita en el Archipiélago Canario son Osilinus atrata Wood, 1828 y Osilinus sauciatus Koch, 1845 (Moro et al., 2003). La primera especie se encuentra en todas las islas, mientras que O. sauciatus sólo está en Lanzarote, Fuerteventura, Gran Canaria, Tenerife (Núñez, 1995; Pérez-Sánchez y Moreno, 1991) y el Archipiélago Chinijo. Sus distribuciones también abarcan otros archipiélagos de la región macaronésica como Madeira, las Islas Salvajes y Cabo Verde para O. atrata (Núñez, 1995; Hawkins et al., 2000) y Madeira y las Islas Salvajes para O. sauciatus (Hawkins et al., 2000). Estas especies muestran morfologías distintas. Osilinus atrata es de color verde-oliváceo con manchas oscuras repartidas de forma irregular, presenta mayor anchura (abertura) que altura, una concha lisa y paredes gruesas (Núñez, 1995; Pérez-Sánchez y Moreno, 1991). Osilinus sauciatus es de coloración pardo-violácea, es más alta que ancha (o con una relación próxima a 1:1), y sus paredes son más gruesas (Núñez, 1995; Pérez-Sánchez y Moreno, 1991) y rugosas que las de O. atrata.
Las diferencias morfológicas entre ambas especies y de distribución horizontal a lo largo del Archipiélago Canario parecen apuntar a una evolución divergente en el pasado. Así, parámetros poblacionales como la abundancia, talla y distribución en el eje vertical deberían reflejar patrones dispares. De este modo, el objetivo general del presente trabajo fue establecer si existe una segregación espacial en el eje vertical y horizontal en las dos especies de Osilinus, burgado hembra (O. atrata) y burgado macho (O. sauciatus), presentes en las Islas Canarias; es decir, identificar los patrones de abundancia y estructura de tallas de las poblaciones de ambas especies en diferentes escalas espaciales estructuradas jerárquicamente. La descripción de los patrones de zonación vertical y horizontal nos permitirá conocer la variabilidad inherente a las poblaciones naturales de estas especies y la importancia de las distintas fuentes de heterogeneidad espacial en las mismas (Menconi et al., 1999; Benedetti-Cecchi, 2001), así como establecer hipótesis sobre los mecanismos ecológicos que gobiernan sobre estos organismos.
Material y métodos
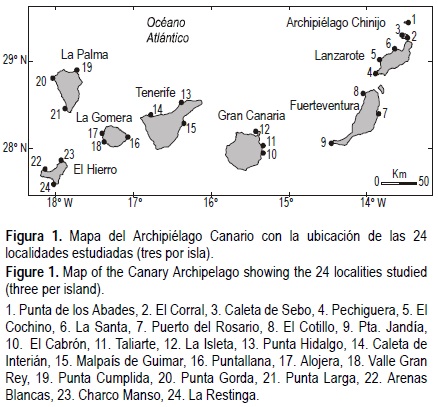
El presente estudio se realizó en las plataformas rocosas del medio intermareal de las Islas Canarias (noroeste del continente africano; 27-30° N, 13-19° W) durante marzo de 2003 (fig. 1), incluyendo un conjunto de islotes al norte de Lanzarote conocido como Archipiélago Chinijo (a partir de ahora, A. Chinijo). En general, el intermareal rocoso del Archipiélago Canario presenta un patrón de zonación bien definido (Navarro et al. , 2005) siguiendo los esquemas de zonación clásicos (Stephenson y Stephenson, 1949). Se estructura en tres bandas: intermareal alto o superior, intermareal intermedio e interma-real bajo o inferior. La banda superior se caracteriza por la existencia de cianofíceas, mientras que en la banda intermedia el crustáceo cirrípedo Chthamalus stellatus es el organismo prominente. La banda inferior está dominada por macroalgas corticadas erectas. Las zonas intermareales examinadas mostraron diferencias en cuanto a su longitud (medida perpendicular a la costa e incluyendo las tres bandas), de manera que éstas mostraron mayor amplitud en las islas orientales (64.66 ± 6.32 m, media ± EE) que en las islas occidentales (45.16 ± 8.51 m, media ± EE).
Diseño muestral
El muestreo se realizó en base a un diseño jerárquico con cuatro escalas espaciales: tres escalas de variabilidad horizontal y una vertical (Underwood, 1997; Kingsford y Battershill, 1998; Menconi et al., 1999). La perspectiva del estudio, utilizando escalas espaciales múltiples organizadas jerárquicamente para el estudio de poblaciones del medio intermareal, permite obtener una mejor interpretación de los patrones de distribución espacial de los organismos (Benedetti-Cecchi, 2001).
En cada una de las islas, separadas entre sí por decenas a centenas de kilómetros, se escogieron al azar tres localidades de sustrato rocoso separadas entre ellas entre 1 y 30 km. En cada localidad se seleccionaron aleatoriamente tres sitios separados por decenas de metros. En cada sitio se estimaron las abundancias de O. atrata y O. sauciatus mediante cuadrados de 50 x 50 cm (0.25 m2) distribuidos al azar en cada una de las tres bandas (n = 10). En cada cuadrado se contarón todos los individuos presentes y se midió su talla mediante el uso de vernieres de 0.1 mm de precisión.
Análisis estadístico
Los datos obtenidos se analizaron mediante análisis de la varianza (ANOVA). Se intentó determinar si las dos especies de Osilinus mantienen abundancias diferentes en las bandas del intermareal siguiendo un patrón determinado para las escalas espaciales del estudio. Se consideraron (1) el factor "Isla" como fijo (ocho niveles correspondientes a las ocho islas); (2) el factor "Banda" como fijo, ortogonal al anterior, y con tres niveles correspondientes a las tres bandas del intermareal (alto, medio y bajo); (3) el factor "Localidad" (aleatorio y anidado en la interacción "Isla x Banda"), y finalmente (4) el factor "Sitio" (aleatorio y anidado en la interacción "Localidad (Isla x Banda)" (Underwood, 1997). El modelo de las fuentes de variabilidad responde, consecuentemente, a la siguiente fórmula: X = µ + Isla + Banda + Isla x Banda + Localidad (Isla x Banda) + Sitio (Localidad (Isla x Banda)) + Residual.
Mediante la prueba de Cochran se detectó una fuerte heterogeneidad de varianzas incluso transformando los datos, por lo que se realizó el análisis empleando un nivel de significancia a = 0.01, en vez de 0.05 (Underwood, 1981). En contrastes que disponen de un volumen de datos considerables, debido a la robustez del ANOVA, las probabilidades asociadas al estadístico F no resultan muy afectadas por la heterogeneidad de varianzas, resultando suficiente elegir un a más pequeño y conservador (Underwood, 1997). Cuando se encontraron diferencias significativas para el factor "Isla" o "Banda" (como interacción o efecto principal), se realizó un contraste a posteriori mediante la prueba de Student-Newman-Keuls (SNK).
Para definir la estructura poblacional de cada especie se obtuvo un diagrama de frecuencias de tallas para la totalidad de los individuos registrados. Contrastamos las diferencias para la talla mediante intervalos de confianza al 95% (a = 0.05) (Underwood, 1997).
Resultados
Osilinus atrata
Esta especie fue la más abundante de las dos, con un total de 2556 individuos censados. La abundancia media para la totalidad del estudio fue de 4.93 ± 0.41 ind m-2 (media ± EE). Los valores mínimos se registraron en la banda inferior (0 ind m-2, en diversas localidades), mientras que los máximos se registraron en la banda intermedia (70.40 ± 8.59 ind m-2 ± EE y 43.73 ± 5.68 ind m-2 ± EE, en Pechiguera y La Santa, respectivamente) de Lanzarote (tabla 1).
Esta especie presentó diferencias significativas para la interacción "Isla x Banda" (F = 4.62, P < 0.001; tabla 2), lo que supone diferencias en abundancia entre islas y, a su vez, una distribución heterogénea entre las bandas de una isla a otra (tabla 1). Las pruebas SNK mostraron que Lanzarote presentaba mayor abundancia que el resto de islas, con diferencias significativas entre las tres bandas estudiadas (tabla 1). Por otro lado, La Gomera y El Hierro no presentaron diferencias entre sí, pero si sus bandas, siendo la banda inferior diferente a la intermedia y superior en ambas islas. El A. Chinijo, Fuerte-ventura, Gran Canaria, Tenerife y La Palma no presentaron diferencias significativas entre ellas, al igual que las bandas dentro de cada una de estas islas. Además, O. atrata mostró una importante variabilidad espacial a pequeña escala, con diferencias significativas entre sitios (F(IxBxL) = 9.29, P < 0.001; tabla 2).
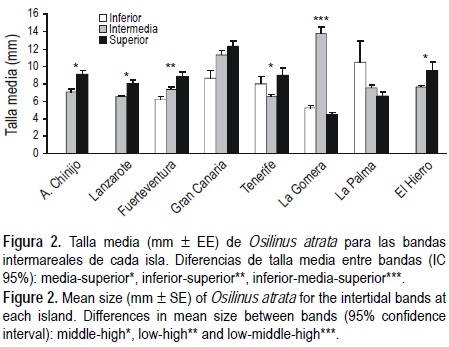
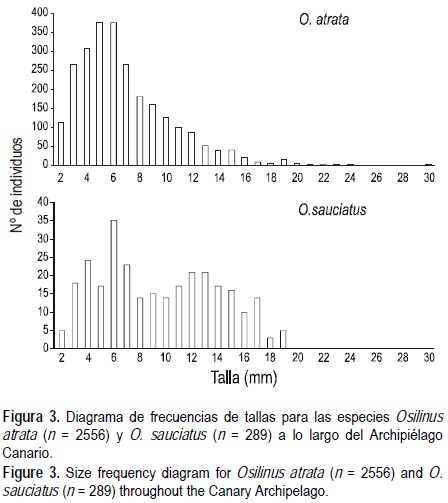
La talla media de O. atrata para el Archipiélago Canario fue de 6.88 ± 0.07 mm (media ± EE), mientras que su talla máxima fue de 30 mm. Los intervalos de confianza obtenidos mostraron las siguientes diferencias y similitudes para las tallas medias: La Gomera ≈ El Hierro < Lanzarote ≈ Tenerife ≈ La Palma ≈ Fuerteventura ≈ A. Chinijo < Gran Canaria. Por otro lado, en cuatro de las islas (A. Chinijo, Lanzarote, Tenerife y El Hierro) la talla media fue mayor en la banda intermareal superior (fig. 2). Igualmente sucedió en Fuerteventura, pero en ella la banda superior no se diferenció de la banda intermedia. La Gomera presentó su mayor talla media en la banda intermedia, mientras que Gran Canaria y La Palma no presentaron diferencias de tallas entre bandas (fig. 2). El diagrama de frecuencias de tallas presenta una clara asimetría positiva (fig. 3), mostrando una cola prolongada en donde se sitúan los individuos de mayor talla, que son los menos abundantes.
Osilinus sauciatus
Esta especie fue menos abundante, habiéndose censando un total de 289 individuos, con un valor medio de abundancia para el conjunto del archipiélago de 0.55 ± 0.11 ind m-2 (media ± EE). Únicamente se encontró en las islas orientales (A. Chinijo, Lanzarote, Fuerteventura y Gran Canaria), en donde alcanzó valores máximos en la banda superior del A. Chinijo (13.73 ± 4.86 ind m-2, El Corral) (tabla 1). Se detectaron diferencias significativas entre islas (F(I) = 3.85, P < 0.01; tabla 2), siendo el A. Chinijo la isla con mayor abundancia y diferente al resto, las cuales no difirieron significativamente entre sí (prueba SNK, P > 0.01). No se hallaron diferencias significativas entre bandas (F(IxBxL) = 1.77; tabla 2); aun así, la distribución de O. sauciatus varió enormemente entre la banda inferior (no se encontró ningún individuo) y la banda superior, donde se hallaron el 91.35% del total de individuos censados (tabla 1). Se detectó, al igual que ocurrió con O. atrata, una fuerte variabilidad a pequeña escala; es decir, diferencias entre sitios separados por decenas de metros (F(IxBxL) = 6.53, P < 0.001; tabla 2).
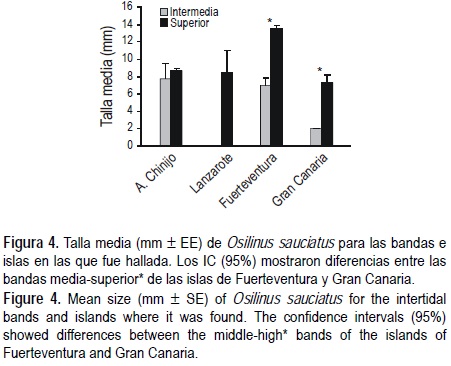
Esta especie presentó una talla media superior a la de O. atrata para todo el estudio de 9.72 ± 0.25 mm (media ± EE), pero una menor talla máxima (20 mm). Fuerteventura presentó una talla media superior que el A. Chinijo y Gran Canaria. A su vez, en Fuerteventura y Gran Canaria la mayor talla media se halló en la banda superior (fig. 4). Debido al escaso número de individuos registrados en Lanzarote, lo que supone intervalos de confianza demasiado amplios, no se encontraron diferencias entre bandas. Además, esta especie presentó una distribución de frecuencias de tallas distinta a la anterior (O. atrata), mostrando una población estructurada en torno a dos clases de tallas (= 2 modas, fig. 3). La primera moda se sitúa entre 5 y 6 mm, al igual que ocurría con O. atrata, y la segunda moda se aprecia sobre 12 mm (fig. 3).
Discusión
Nuestro estudio detectó diferencias en los patrones de distribución, abundancia y estructura de tallas de las dos especies de gasterópodos del género Osilinus presentes en el medio intermareal rocoso de las Islas Canarias. Estas diferencias son más patentes a escalas pequeñas (sitios separados por decenas de metros) y entre bandas (variabilidad vertical, sensu Benedetti-Cecchi, 2001), que a escalas mayores (variabilidad horizontal a gran escala: diferencias entre islas y localidades dentro de cada isla). Consecuentemente, nuestras observaciones revelaron una importante variabilidad espacial, tal y como registraron Menconi et al. (1999) para la especie Osilinus turbinatus en el Mediterráneo y Underwood y Chapman (1996), Benedetti-Cecchi (2001) y Navarro et al. (2005) para otras especies de invertebrados gasterópodos del intermareal rocoso de costas de mares templados. Para explicar estos cambios espaciales hay que entender el efecto de multitud de procesos y factores que afectan a los organismos presentes en el medio intermareal (Thompson et al., 2002).
Los resultados hallados muestran que O. atrata fue más abundante a lo largo del intermareal rocoso del Archipiélago Canario que O. sauciatus, tal y como Núñez (1995) observó en la isla de Fuerteventura. Esta especie presentó mayor abundancia en la banda intermedia, mientras que O. sauciatus se centró mayoritariamente en la banda superior, si bien para esta última especie no encontramos diferencias significativas. Respecto a la talla, O. sauciatus fue mayor que O. atrata, aunque ambas especies mostraron una tendencia hacia tallas mayores en la banda superior. Estas diferencias encontradas en el patrón vertical podrían explicarse en base a la arquitectura del hábitat, la orientación del sustrato, incidencia del oleaje, etc., que influyen sobre la distribución de organismos intermareales forzando una importante variabilidad a esta escala espacial (Dayton, 1971; Todgham et al., 1997; Menconi et al., 1999; Chapman, 2000; Benedetti-Cecchi, 2001; Rilov et al., 2005). En el caso particular del efecto del oleaje, se sabe que los gasterópodos intermareales de zonas expuestas tienen conchas más delgadas y lisas con amplias aberturas (mayor anchura) que los presentes en zonas protegidas (Boulding, 1990). En nuestro caso, esto coincide con las morfologías diferenciadas de ambas especies de burgados, lo que unido a sus distribuciones en las diferentes bandas y tallas obtenidas, podrían reflejar el efecto del oleaje a lo largo del eje vertical. Osilinus atrata, debido a su menor talla, podría refugiarse en grietas y huecos y ser menos vulnerable al desalojo por el oleaje. A su vez, y gracias a su mayor abertura oral, dispone de un mayor pie con el que se adherirse al sustrato. Por otro lado, O. sauciatus, de mayor altura, se vería más afectado por el oleaje, dificultándole su introducción en grietas. Esta morfología le permitiría, sin embargo, retener más agua evitando la pérdida con su menor abertura y soportando así mejor la desecación durante los largos periodos de emersión (Bertness, 1999).
Se han observado diferencias importantes en el ámbito regional (diferencias entre islas), como es la distribución restringida de O. sauciatus en las islas orientales citada previamente por Pérez-Sánchez y Moreno (1991) y Núñez (1995). Además, ambas especies presentaron sus máximas abundancias en las islas más orientales del A. Chinijo (O. sauciatus) y Lanzarote (O. atrata). Como factores potenciales que podrían explicar esta diferencia en distribución y abundancia de las especies de Osilinus podríamos citar (1) la superior productividad de las aguas de las islas orientales (Llinás et al., 1994; Davenport et al., 2002), que permitiría un aumento de la disponibilidad de alimento y recursos nutritivos (sensu Menge, 2000), y (2) la mayor antigüedad de las islas orientales respecto a las occidentales (Afonso, 1988). La presencia de plataformas intermareales más extensas en las islas orientales (tal y como apreciamos) favorecería la disipación de la energía del oleaje y un mayor espacio para el asentamiento de larvas y desarrollo de ambas especies.
Finalmente, y respecto a la estructura demográfica, se observan modelos dispares para ambas especies. Así, O. atrata presenta un claro desplazamiento de la curva hacia las tallas de menor tamaño. Considerando que las especies de Osilinus han sido consumidas desde la antigüedad (Batista, 2001) hasta la actualidad (Aguilera et al., 1994; Núñez, 1995), que en la mayoría de las islas los individuos de mayor talla se sitúan en la banda superior (más accesible al marisqueo) y, observando lo que sucede con otras especies de gasterópodos intermareales de la zona (Navarro et al., 2005), detectamos un considerable efecto de la recolección (marisqueo) sobre la estructura poblacional de O. atrata. Este efecto no es observable en O. sauciatus debido, suponemos, a su menor abundancia y distribución más restringida en el Archipiélago Canario.
Agradecimientos
Este estudio se realizó dentro del proyecto "Canarias, por una Costa Viva" del Ministerio de Medio Ambiente (Secretaría General de Costas) y la Universidad de Las Palmas de Gran Canaria (http://www.canariasporunacostaviva.org). Los autores agradecen la participación de todos los colaboradores y voluntarios que han intervenido en las campañas de investigación. También queremos agradecer a los dos revisores anónimos de la revista por sus contribuciones para la mejora del presente artículo.
Referencias
Afonso, L. (ed.) (1988). Geografía de Canarias. Interinsular, Santa Cruz de Tenerife. [ Links ]
Aguilera, F., Brito, A., Castilla, C., Díaz, A., Rodríguez, A., Sabaté, F. y Sánchez, J. (1994). Canarias: Economía, Ecología y Medio Ambiente. Francisco Lemus Editor, Islas Canarias, 361 pp. [ Links ]
Batista, C. (2001). El marisqueo en la prehistoria de Gran Canaria. Vector Plus, 18: 67-76. [ Links ]
Benedetti-Cecchi, L. (2001). Variability in abundance of algae and invertebrates at different spatial scales on rocky sea shores. Mar. Ecol. Prog. Ser., 215: 79-92. [ Links ]
Bertness, M.D. (1999). The Ecology of Atlantic Shorelines. Sinauer Associates, 417 pp. [ Links ]
Boulding, E.G. (1990). Are the opposing selection pressures on exposed and protected shores sufficient to maintain genetic differentiation between gastropod populations with high intermigration rates? Hydrobiologia, 193: 41-52. [ Links ]
Chapman, M.G. (2000). A comparative study of differences among species and patches of habitat on movements of three species of intertidal gastropods. J. Exp. Mar. Biol. Ecol., 244: 181-201. [ Links ]
Davenport, R., Never, S., Helmke, P., Pérez-Moreno, J. and Llinás, O. (2002). Primary productivity in the northern Canary Islands region as inferred from Seawifs imagery. Deep-Sea Res. II, 49: 3481-3496. [ Links ]
Dayton, P.K. (1971). Competition, disturbance and community organization: The provision and subsequent utilization of space in a rocky intertidal community. Ecol. Monogr., 41: 351-389. [ Links ]
Hawkins, S.J., Corte-Real, H.B.S.M., Pannacciulli, F.G., Weber, L.C. and Bishop, J.D.D. (2000). Thoughts on the ecology and evolution of the intertidal biota of the Azores and other Atlantic islands. Hydrobiologia, 440: 3-17. [ Links ]
Harley, C. and Helmuth, B. (2003). Local- and regional-scale effects of wave exposure, thermal stress and absolute versus effective shore level on patterns of intertidal zonation. Limnol. Oceanogr., 48: 1498-1508. [ Links ]
Kingsford, M. and Battershill, C. (1998). Studying Marine Temperate Environments: A Handbook for Ecologists. Canterbury Univ. Press, Christchurch, New Zealand, 335 pp. [ Links ]
Llinás, O., Rueda, M.J. y Pérez-Martell, E. (1994). Características termohalinas y nutrientes en aguas de las plataformas insulares canarias a finales de primavera. Bol. Inst. Esp. Oceanogr., 10(2): 177-189. [ Links ]
Menconi, M., Benedetti-Cecchi, L. and Cinelli, F. (1999). Spatial and temporal variability in the distribution of algae and invertebrates on rocky shores in the northwest Mediterranean. J. Exp. Mar. Biol. Ecol., 233: 1-23. [ Links ]
Menge, B.A. (2000). Top-down and bottom-up community regulation in marine rocky intertidal habitats. J. Exp. Mar. Biol. Ecol. 250: 257-289. [ Links ]
Moro, L., Martín, J.L., Garrido, M.J. y Izquierdo, I. (eds). (2003). Lista de Especies Marinas de Canarias (Algas, Hongos, Plantas y Animales). Consejería de Política Territorial y Medio Ambiente del Gobierno de Canarias, 248 pp. [ Links ]
Navarro, P.G., Ramírez, R., Tuya, F., Fernández-Gil, C., Sánchez-Jerez, P. and Haroun, R.J. (2005). Hierarchical analysis of spatial distribution patterns of patellid limpets in the Canary Islands. J. Molluscan Stud., 71: 67-73. [ Links ]
Núñez, J. (1995). Estudio Ecológico del Mejillón y Otros Moluscos en la Isla de Fuerteventura. Departamento de Biología, Universidad de La Laguna, 150 pp. [ Links ]
Pérez-Sánchez, J.M. y Moreno, E. (1991). Invertebrados Marinos de Canarias. Cabildo Insular de Gran Canaria, 335 pp. [ Links ]
Rilov, G., Gasith, A. and Benayahu, Y. (2005). Effect of disturbance on foraging: Whelk activity in wave-exposed rocky shores with minimal tidal range. Mar. Biol., 147: 421-428. [ Links ]
Stephenson, T.A. and Stephenson, A. (1949). The universal features of zonation between tide-marks on rocky coasts. J. Ecol., 37: 289-305. [ Links ]
Thompson, R.C., Crowe, T.P. and Hawkins, S.J. (2002). Rocky intertidal communities: Past environmental changes, present status and predictions for the next 25 years. Environ. Conserv., 29: 168-191. [ Links ]
Todgham, A., Boulding, E. and Megill, W.M. (1997). Wave exposure effects on the density, habitat selection and size distribution of intertidal limpets species of the Pacific coast of British Columbia. In: W.M. Megill (ed.), Annual Scientific Report of the Coastal Ecosystems Research Foundation. 1997. <http://cerf.bc.ca/pubs/asr97> [ Links ].
Underwood, A.J. (1981). Techniques of análisis of variance in experimental marine biology and ecology. Oceanogr. Mar. Biol. Annu. Rev., 19: 513-60. [ Links ]
Underwood, A.J. (1997). Experiments in Ecology: Their Logical Design and Interpretation Using Analysis of Variance. Cambridge Univ. Press, Cambridge, UK. [ Links ]
Underwood, A.J. and Chapman, M.G. (1996). Scales of spatial patterns of distribution on intertidal invertebrates. Oecologia, 107: 212-224. [ Links ]

