










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkCiencias marinas
versão impressa ISSN 0185-3880
Cienc. mar vol.30 no.3 Ensenada Set. 2004
Artículos
Efecto de la radiación UV en la inactivación genética del esperma de botete diana Sphoeroides annulatus (Jenyns, 1842)
Effect of UV radiation on the genetic inactivation of sperm of the bullseye puffer Sphoeroides annulatus (Jenyns, 1842)
Lenin Arias-Rodríguez*, Luz Estela Rodríguez-Ibarra y Gabriela del Valle-Pignataro**
Laboratorio de Genética, Centro de Investigación en Alimentación y Desarrollo, A.C., Unidad Mazatlán Sábalo-Cerritos s/n, Estero del Yugo Apartado postal 711 Mazatlán, Sinaloa, México. *E-mail: larias@victoria.ciad.mx, **gvp@victoria.ciad.mx
Recibido en marzo de 2003;
aceptado en marzo de 2004.
Resumen
La inactivación genética (ADN) del esperma de peces se realiza mediante luz ultravioleta que, en irradiaciones crecientes, genera efectos paradójicos (efecto Hertwig) en los porcentajes de supervivencia. En este trabajo se diluyeron muestras de semen provenientes de diez machos de botete diana (Sphoeroides annulatus) en solución extendedora Cortland modificada en dilución 1:50, y se utilizaron para probar el efecto de nueve dosis de radiación ultravioleta (0.2-1.0 Jcm-2) sobre la duración de la motilidad en segundos, el índice de motilidad del esperma, y los porcentajes de supervivencia de embriones obtenidos de la fertilización de huevos provenientes de cinco hembras de la misma especie. Los tiempos de motilidad del esperma en las muestras irradiadas con 0.2-0.9 Jcm-2 resultaron estadísticamente no diferentes de los controles, mientras que las muestras irradiadas con 1.0 Jcm-2 fueron significativamente menores. El índice de motilidad (IM), por otra parte, permitió la diferenciación estadística de cuatro grupos en cuanto a su respuesta a diferentes dosis de radiación. El primer grupo se caracterizó por valores altos del IM, e incluyó a los controles y a las muestras irradiadas con 0.2-0.3 Jcm-2; el segundo agrupó a las irradiaciones de 0.4 a 0.7 Jcm-2, en las cuales se observó un descenso del IM; en el tercer grupo (0.8-0.9 Jcm-2) el IM aumentó nuevamente; y en el último (1.0 Jcm-2) se observó el menor IM. En cuanto a los porcentajes de supervivencia, se observó una curva descendente con valores altos de supervivencia en los controles y en las muestras irradiadas con 0.2 Jcm-2, con un descenso en los tratamientos de 0.3 a 0.4 Jcm-2, y una recuperación significativa a partir del tratamiento de 0.5 y hasta 0.8 J cm-2, observándose nuevamente un decremento en la supervivencia de los tratamientos de 0.9 a 1.0 Jcm-2. Ambos resultados, IM y supervivencia, indican que la dosis óptima para la producción de organismos haploides se encuentra cercana a 0.7 J cm-2. En el tratamiento de 0.7 J cm-2 se observaron larvas con las características típicas del síndrome haploide, indicando nuevamente que ésta puede ser la dosis más adecuada para producir organismos ginogenéticos.
Palabras clave: Sphoeroides annulatus, semen, ultravioleta, Hertwig.
Abstract
Genetic (DNA) inactivation of fish sperm with ultraviolet irradiation is generally accompanied by a paradoxical effect on survival rates (Hertwig effect). In the present study, sperm samples from ten male bullseye puffer fish (Sphoeroides annulatus) were diluted 1:50 using Cortland's extender solution and used to test the effect of nine ultraviolet doses (0.2-1.0 Jcm-2) on motility time in seconds, motility index, and embryo survival rate after fertilizing eggs from five bullseye puffer females. Motility time of sperm irradiated with 0.2-0.9 J cm-2 were not statistically different from the controls, but sperm irradiated with a dosage of 1.0 Jcm-2 dosage had significant lower motility time. Motility indices (MI) allowed for the statistical differentiation offour groups in relation to their response to different radiation doses: the first had high MI, and included the controls and 0.20.3 Jcm-2 treatments; the second had lower MI and included the 0.4-0.7 Jcm-2 treatments; the third showed recovery of MI and included the 0.8-0.9 Jcm-2 treatments; and the fourth showed the lowest MI with the 1.0 Jcm-2 treatment. Embryo survival was highest for the controls and 0.2 Jcm-2 treatment, decreasing in the 0.3-0.4 Jcm-2 treatments, increasing again in the 0.50.8 Jcm-2 treatments, until reaching lowest survival in the 0.9-1.0 Jcm-2 treatments. These results indicate that the best ultraviolet dosage to achieve genetic inactivation of sperm of this species is close to 0.7 Jcm-2, a dosage in which fish fry showed typical haploid syndrome characteristics.
Key words: Sphoeroides annulatus, sperm, ultraviolet, Hertwig.
Introducción
La fertilización externa en los peces facilita la manipulación de los gametos y las características genéticas que éstos portan (Purdom, 1983). Esto permite la inducción de la ginogénesis artificial, en la cual los huevos son activados por esperma que no contribuye genéticamente en el desarrollo de los embriones (Lasher y Rugh, 1962; Purdom, 1969; Thorgaard, 1983; Arai, 2000), produciéndose así haploides que no sobreviven más allá de la etapa de eclosión (Lasher y Rugh, 1962; Hussain et al., 1993). El restablecimiento de la diploidía en huevos fecundados con esperma cuyo complemento genético ha sido inactivado se logra mediante la supresión de la segunda meiosis por retención del segundo cuerpo polar o de la primera división mitótica, aplicando choques de temperatura y presión hidrostática en las primeras etapas posteriores a la fertilización (Mair, 1993; Arai, 2000).
Para la inactivación genética del esperma se emplean diversas fuentes de radiación. Entre ellas se encuentran los rayos gamma, los rayos X yla radiación ultravioleta (UV) (Chourrout, 1982; Don y Avtalion, 1993; Galbusera et al., 2000; Gomelsky et al., 2000). Debido a que el empleo de radiaciones ionizantes como los rayos gamma y X generan individuos con características residuales de origen paterno (Chourrout, 1984), la inactivación completa del ADN de los espermatozoides se logra con luz UV (Jenneckens et al., 1999). Este tipo de radiación produce la fragmentación del ADN y la formación de dímeros de timina. A pesar de existir estudios sobre inactivación de ADN en esperma en múltiples especies de peces, el empleo y optimización de esta metodología requiere del ensayo de las dosis para definir cuáles generan porcentajes de supervivencia adecuados a las condiciones particulares de cada especie. En forma general se sabe que el incremento de las dosis genera una disminución en la viabilidad de los embriones, hasta que con dosis altas se presenta una mortalidad total. Asimismo, un fenómeno paradójico conocido en este tipo de experimentos es el llamado efecto de Hertwig, en el cual se observa que con ciertas dosis de radiación intermedia la supervivencia de los embriones muestra una ligera recuperación con respecto a dosis menores. Este fenómeno ha sido explicado por la supervivencia de espermatozoides cuyo ADN ha sido fragmentado por la radiación (y digerido por las nucleasas del huevo fertilizado), pero que conservan, no obstante, la motilidad necesaria para activar el desarrollo del huevo, que porta únicamente material genético de procedencia materna (Purdom, 1969, 1983).
Esta técnica tiene aplicaciones importantes, como son la generación de cultivos monosexuales, y la selección de líneas puras (clones) (Taniguchi et al., 1993) para caracteres de interés como resistencia a enfermedades, y para la mejora en el crecimiento de las especies manipuladas (Yamamoto, 1999; Arai, 2000).
La aplicación de estas técnicas en México es reciente, habiendo algunos reportes en peces marinos entre los que se encuentra un estudio de inactivación genética de esperma mediante luz UV en cabrilla, Paralabrax maculatofasciatus (Huerta, 1999) y otro en botete diana, Sphoeroides annulatus (Arias-Rodríguez, 2001). Esta última especie se consume en la costa noroeste de México desde los años setenta (Castellanos et al. , 1982), alcanzando alto precio en los mercados locales debido a la blancura y excelente textura y sabor de su carne. Además, especies de la misma familia, como Takifugu rubripes, también tienen un alto valor en el mercado en Japón, Corea y China (Kanazawa, 1991), por lo que el potencial de exportación del botete diana es alto. Su cultivo se inició en Mazatlán, México, en 1996 en el Centro de Investigación en Alimentación y Desarrollo (CIAD), lográndose avances significativos a la fecha (Abdo-de la Parra et al., 2001; Martínez-Palacios et al., 2002; Duncan et al., 2003).
El objetivo del presente trabajo fue evaluar el efecto de nueve dosis de radiación UV sobre el tiempo de movimiento (T) y el índice de motilidad (IM) de los espermatozoides del botete diana (S. annulatus), así como los porcentajes de supervivencia de los embriones a las 24, 48 y 72 h de incubación, como un primer paso en el establecimiento de las condiciones óptimas para producir organismos cromosómicamente manipulados.
Materiales y métodos
Los reproductores fueron capturados en la Bahía de Teacapan, municipio de Escuinapa (Sinaloa, México), durante los meses de reproducción (abril a junio de 2000), y transportados al CIAD, Unidad Mazatlán, donde fueron mantenidos bajo condiciones de cautiverio. Se seleccionaron diez machos, con longitud de 28.77 ± 2.32 cm y peso de 605.68 ± 143.04 g, y cinco hembras con 32.88 ± 3.56 cm de longitud y 1121.0 ± 402.91 g de peso, en promedio.
Para la recolección de los gametos los reproductores fueron anestesiados con 0.75 mLL-1 de 2-fenoxi-etanol, diluido en agua de mar. El esperma se recolectó en jeringas estériles de 5 mL, evitando la contaminación del semen con excretas u orina que debilitan la capacidad fecundante de los espermatozoides (Aas et al., 1991; Satterfield y Flickinger, 1995). Los gametos femeninos fueron recolectados en recipientes de plástico, aplicando ligera presión abdominal a hembras inyectadas con LHRHa (Sigma Chemical®) (Duncan y Rodríguez, 2001).
Radiación de esperma con UV
Se utilizó la solución Cortland modificada (SCM) extendedora de la motilidad (Geffen y Evans, 2000) para comparar el efecto de nueve dosis de radiación UV en el semen de los diez reproductores. La SCM se compone de solución A: KCl (0.9095 g), NaCl (0.2337 g), CaCl2 (0.0332 g), NaH2PO4H2O (0.0551 g), MgSO4 (0.0739 g), y solución B: NaHCO3 (0.4956 g), D-(+)-C6H12O6 (0.5045 g) (cada solución en 100 mL de H20 destilada), mezcladas en proporción 4:1 a pH 6.74 ± 0.11 y empleadas en fresco. En los experimentos se empleó la dilución 1:50 (semen:SCM).
El experimento consistió de dos controles (A y B) y del tratamiento C, cada uno con tres réplicas: A = 0.2 mL de semen sin diluir en SCM por réplica, B = semen + SCM, y C = semen + SCM + radiación UV. Para B y C se mezclaron 1.8 mL de semen en 88.2 mL de SCM (1:50) de los que se colocaron 10 mL en cada una de tres cajas petri de vidrio de 8.8 cm de diámetro interior, lo que permitió mantener una película de 1.6 mm (h = V/πr2 = 10 mL/3.1416 [4.4 cm]2 = 0.16 cm). En el tratamiento C se aplicaron nueve dosis de radiación UV de 0.2 a 1.0 J cm-2 con intervalos de 0.1 J cm-2. Debido al escaso volumen de semen recolectado, sólo se pudieron hacer por duplicado las dosis 0.6, 0.8 y 0.9.
Las muestras fueron irradiadas en una cámara de UV con lámpara germicida de 15 W (General Electric, modelo G15T8) y agitador orbital (Felisa, FE-315) de 100 rpm.
Los tiempos de exposición requeridos para estas dosis se calcularon aplicando el modelo sugerido por Christensen y Tiersch (1994):

donde E es la exposición total en J cm-2, n es el número de lámparas (1), w es el rendimiento de la lámpara a 254 nm (3.3 W), t es la duración de la exposición en segundos (x), d2 es la distancia de la muestra en relación con la lámpara (10 cm), y π2 (3.14162). Por ejemplo, para alcanzar una dosis de 0.2 J cm-2 fue necesario aplicar 60 s de exposición a la luz UV.
Calidad y motilidad de esperma
Se verificó la calidad del esperma observando su tipo de movimiento, de acuerdo con la escala gradual (4, 3, 2, 1, 0) señalada por Menkveld y Kruger (1996). El grado 4 indica el máximo desplazamiento de los espermatozoides, el grado 3 describe el descenso de su desplazamiento y el incremento de movimientos laterales, el grado 2 representa poco o nulo desplazamiento y la disminución de los movimientos laterales, el grado 1 indica muy poco movimiento de la cabeza de los espermatozoides, junto con el decremento del movimiento del flagelo, y el grado 0 representa nulo desplazamiento de los espermatozoides.
Simultáneamente fue examinado el porcentaje de células activas empleando el criterio sugerido por Aas et al. (1991), en una escala de 0% a 100%, con intervalos de 10 (p.e., 10%, 20%, 30%, 40%, etc.), y el tiempo del movimiento en segundos (T) de los espermatozoides desde la activación con agua de mar hasta el grado 3 de movimiento de la escala descrita. Únicamente fueron consideradas útiles para los experimentos las muestras de semen provenientes de machos con movimiento de grado 4 en la escala descrita y con un 90% a 100% de células espermáticas activas (Aas et al., 1991).
A partir de dichos parámetros se estimó el IM, multiplicando la graduación del movimiento por el porcentaje de células activas (p.e., 4 x 100 igual a un IM de 400). Un IM de 100% es considerado como el mínimo necesario para fertilizar un óvulo (Satterfield y Flickinger, 1995).
Los parámetros de calidad del semen (T, porcentaje de células activas y tipo de movimiento) fueron evaluados después de activar 50 de semen de cada réplica con una gota de agua de mar (tratada con luz UV y filtrada con cartuchos de 10 y 1 nm de diámetro de poro).
Fertilización artificial e incubación
Las hembras empleadas en los experimentos fueron inducidas al desove con LHRHa (Sigma Chemical®), siguiendo las recomendaciones de Duncan y Rodríguez (2001). Para la fertilización se utilizaron 1.67 ± 0.47 g (media ± desviación estándar) de huevos por cada réplica. La fertilización artificial se llevó a cabo mezclando los espermatozoides y los huevos suavemente, y activándolos con agua de mar. Las muestras (réplicas) de semen A y B fueron activadas con agua de mar en dilución 1:100, con 19.8 mL, y las del tratamiento C con 10 mL (Duncan y Rodríguez, 2001).
Debido a que los huevos de esta especie se adhieren y el vidrio resultó ser una superficie adecuada, se utilizaron tres portaobjetos por réplica. La incubación se realizó en cubetas de plástico oscuro con capacidad de 8 L, con flujo continuo de agua tratada con luz UV y filtros de cartucho de 20 y 5 de diámetro de poro. El fotoperiodo fue controlado a 11 h de luz y 13 h de oscuridad.
En cada portaobjetos se hicieron tres conteos de huevecillos vivos y muertos en campos seleccionados al azar, de acuerdo a las características señaladas por Yamahira (1997), con ligeras modificaciones conforme a lo siguiente: los huevecillos vivos son esféricos y transparentes, con pequeños glóbulos de aceite en el citoplasma, muestran movimientos esporádicos a las 24 h de incubación, y los muertos son blanquecinos y opacos, en algunos casos con el saco de vitelo fragmentado.
Análisis estadísticos
Las variables consideradas fueron el IM (Satterfield y Flickinger, 1995), la duración del movimiento de las células espermáticas en segundos (T) y los porcentajes de supervivencia de los embriones a las 24, 48 y 72 h de incubación.
La supervivencia (S) fue calculada de acuerdo a la fórmula:

donde Ev es el número de embriones vivos por conteo a las 24, 48 y 72 h; Em, los embriones muertos por conteo a las 24, 48 y 72 h; y 100, para obtener conversión porcentual.
Los IM estimados de cada tratamiento y los porcentajes de supervivencia a las 24, 48 y 72 h de incubación fueron transformados a la función arcoseno con el propósito de lograr la aditividad de efectos (Zar, 1984). Los datos de IM, T y los porcentajes de supervivencia fueron comparados por análisis de varianza de dos vías realizados por separado y de acuerdo con el tipo de variable. Se estableció el valor de significancia a P ≤ 0.05. Se aplicaron las pruebas de comparación múltiple de Tukey y Dunn (Zar, 1984; Danield, 1990).
Resultados
Al analizar las muestras irradiadas se encontraron diferencias significativas en la duración del movimiento en segundos, T (P = 0.0017). La comparación múltiple indicó que dichas diferencias se deben solamente al valor T del tratamiento C irradiado con la dosis de 1.0 J cm-2 (12.0 ± 20.7 s; media ± desviación estándar). Las T mínima y máxima (21.0 ± 0.0 y 40.0 ± 0.0 s, respectivamente) de los grupos controles A y B no mostraron diferencias significativas entre sí. Las muestras irradiadas con las dosis de 0.2 a 0.9 J cm-2 mostraron valores aparentemente altos (25.6 ± 2.3 y 42.3 ± 4.0 s, respectivamente) comparados con los de las muestras control, aunque no hubo diferencias significativas (fig. 1).
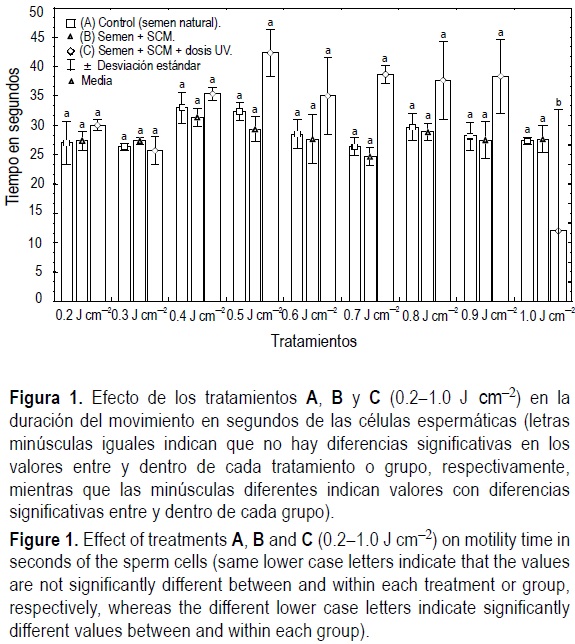
En todos los tratamientos se observó esperma con grado 4 de movimiento al ser activado. Los IM (fig. 2) presentaron diferencias significativas entre grupos y entre tratamientos para el grupo tratado (P = 0.001). La comparación múltiple de Tukey mostró que el grupo C se caracterizó por cuatro respuestas en el IM observado. El índice de motilidad más alto correspondió a esperma sometido a las dosis de 0.2 a 0.3 J cm-2, con un IM no estadísticamente diferente de los controles. En un segundo grupo se encontraron los IM de esperma sometido a las dosis de 0.4 a 0.7 J cm-2, con IM no diferentes entre sí, pero significativamente menores que los controles. La tercer respuesta correspondió a los IM de las dosis de 0.8 a 0.9 J cm-2, observándose un aumento en los IM, pero siendo aún significativamente diferente de los controles. Por último, el cuarto grupo, en que el esperma fue sometido al tratamiento con una dosis de 1.0 Jcm-2, resultó significativamente diferente a los controles (fig. 2).

Los porcentajes de supervivencia mostraron una curva con cuatro fases a las 24, 48 y 72 h de incubación. El análisis de varianza indicó diferencias significativas (P = 0.001) sólo en el grupo C y entre periodos de incubación (24, 48 y 72 h) (P = 0.001). Los controles A y B no mostraron diferencias significativas entre sí, por lo que fueron considerados como un solo grupo en la figura 3 y de acuerdo con los periodos de incubación. La comparación múltiple de Tukey mostró que la primera fase de la curva de supervivencia corresponde a los embriones de los grupos controles A y B, y a los embriones del grupo C sometidos a la dosis de 0.2 J cm-2, con porcentajes de supervivencia que variaron de 100 ± 0.0 (a las 24 h) a 89.7 ± 4.20 % (a las 72 h), sin diferencias significativas entre sí. La segunda fase de la curva muestra un descenso en la supervivencia de los embriones en las dosis de 0.3 a 0.4 J cm-2, con intervalos de 76.6 ± 17.8 (a las 24 h) a 17.1 ± 12.8 % (a las 72 h). En la tercera fase se observó un incremento significativo en la supervivencia correspondiente a embriones fertilizados con esperma UV irradiado con dosis de 0.5 a 0.8 J cm-2, con porcentajes de 99.8 ± 0.35 (a las 24 h) a 26.3 ± 19.8 (a las 72 h) (fig. 3). La cuarta fase, que corresponde a las dosis de 0.9 a 1.0 Jcm-2, mostró un descenso significativo en la supervivencia de embriones, con valores de 90.1 ± 3.18 (a las 24 h) a 0.0 % (a las 72 h) en la dosis de 1.0 J cm-2 (fig. 3).
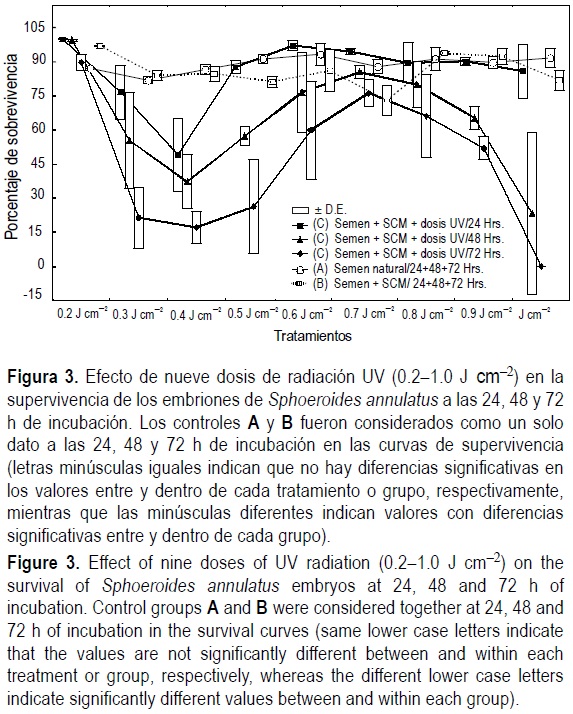
Se observó un descenso significativo en la supervivencia de 24 a 48 h y de 48 a 72 h, el cual fue causado principalmente por el incremento en mortalidad de los embriones del grupo C, observándose los efectos más drásticos cuando se considera la dosis de 1.0 J cm-2 (fig. 3).
El análisis del desarrollo morfológico de los embriones fotografiados permitió identificar un patrón de desarrollo relativamente homogéneo con las dosis de radiación intermedia (0.5 a 0.8 J cm-2), el cual se caracterizó por la presencia de una columna vertebral doblada, cola reducida, saco vitelino grande en relación con el tamaño de los embriones, y ojos acromáticos (fig. 4a), características comúnmente observadas en embriones haploides. Los embriones diploides (controles) se presentan en la figura 4b, en la que se observa la columna vertebral recta y larga, saco vitelino proporcional al tamaño de los embriones (los glóbulos de aceite son aparentemente más pequeños y en mayor cantidad) y ojos cromáticos. Con las dosis menores a 0.4 J cm-2 y mayores a 0.8 J cm-2 el desarrollo morfológico de los embriones fue anormal y desordenado. Estas observaciones morfológicas coinciden con los porcentajes más altos de supervivencia a las 72 h, los cuales se obtuvieron utilizando una dosis de 0.7 J cm-2. Las larvas resultantes (fig. 4c) se caracterizaron por la presencia de una columna vertebral doblada y corta, ojos acromáticos, saco vitelino distorsionado y tubo digestivo atrofiado, esto es, con el llamado "síndrome haploide". Comparativamente, las larvas de los controles mostraron columna vertebral recta y saco vitelino proporcional al tamaño de las larvas y los ojos mostraron pigmentación y tubo digestivo con desarrollo normal (fig. 4d).
Discusión
Los resultados de la duración del movimiento en segundos (T) con todas las dosis de radiación evaluadas (a excepción de 1.0 J cm-2) no mostraron diferencias significativas con relación a los controles experimentales (A y B), lo que difiere de lo reportado en otros estudios. Se ha identificado que la radiación UV reduce el tiempo del movimiento de las células espermáticas (Don y Avtalion, 1993; Huerta, 1999), y en algunos casos (P. maculatofasciatus) a valores menores que los registrados en los grupos control (Huerta, 1999).
En Oreochromis niloticus, Don y Avtalion (1993) identificaron que la dosis de 0.09 J cm-2 no genera daños en las mitocondrias espermáticas, mientras que las dosis de 0.27 a 0.57 J cm-2 sí, observándose como cápsulas vacías. En las dosis de radiación menores a 0.2 J cm-2 la motilidad de las células espermáticas no fue afectada significativamente, mientras que en dosis superiores a 0.9 J cm-2 las células espermáticas fueron lesionadas en su totalidad siendo característica la pérdida del flagelo. En el presente trabajo, los valores extremadamente bajos de T en la dosis de 1.0 J cm-2 (fig. 1) permiten inferir que los espermatozoides fueron dañados, aunque no se lograron hacer observaciones directas de los mismos.
El hecho de no encontrar un efecto sobre la motilidad de los espermatozoides en el presente estudio (dosis de 0.2 a 0.9 J cm-2) puede ser explicado con base en resultados previos en trucha y carpa, en las que se encontró que el movimiento de los espermatozoides no depende totalmente de la energía (ATP) proveniente de las mitocondrias (Billard y Cosson, 1992; Perchec et al., 1995). En estas especies se ha identificado que el nivel de ATP intracelular disminuye en la fase activa de los espermatozoides, pero éste se incrementa gradualmente antes de que la motilidad se detenga (Billard y Cosson, 1992), en parte porque es almacenado en el citoplasma previamente a su utilización (Perchec et al., 1995). La concentración de cationes como el Mg2+ (de la SCM) reducen la acción inhibitoria de la motilidad por parte del K+ como ha sido probado en Takifugu niphobles (Takai y Morisawa, 1995). También se ha demostrado que el fosfato de creatinina en semen de trucha ayuda a mantener la motilidad por un tiempo superior a los 30 s (Cosson et al., 1991).
Con respecto a los IM, en el primer grupo con dosis de 0.2 y 0.3 J cm-2, 100% y 93.3% de las células activas, respectivamente, presentaron valores más altos que los encontrados por Christensen y Tiersch (1994) en Ictalurus punctatus (77.0% y 64.0%, respectivamente). Las diferencias encontradas indican que la susceptibilidad de las células espermáticas a la radiación UV varía dependiendo de la especie y de las condiciones experimentales. Christensen y Tiersch (1994) emplearon testículos macerados de bagre en combinación con una solución extendedora, lo que no favoreció un decremento en la penetración de la radiación en los espermatozoides y causó que los porcentajes de células activas fueran bajos.
En el segundo grupo (0.4 a 0.7 J cm-2) se identificó un decremento en los IM, con células activas que mostraron valores de 83.3% a 43.3%, también superiores a los obtenidos por Christensen y Tiersch (1994), con un máximo (con 0.4 J cm-2) y mínimo (con 0.7 J cm-2) de 30% y 20% de células activas, respectivamente. En el tercer grupo (0.8 a 0.9 J cm-2) se observó el aumento de los IM y del porcentaje de células activas de 61.6% a 51.6%, valores que fueron altos en relación con la lisis de células espermáticas reportada por Don y Avtalion (1993) en O. niloticus. Los resultados con la dosis de 1.0 J cm-2 fueron característicos del cuarto grupo, coincidiendo con los valores de 5.0% de células activas obtenidos por Christensen y Tiersch (1994).
Los porcentajes de supervivencia en los periodos de incubación (24, 48 y 72 h) muestran el patrón clásico del efecto Hertwig que se caracterizó por cuatro fases (fig. 3):
La primera mostró porcentajes de supervivencia altos en los controles A y B, y con la dosis de 0.2 J cm-2 del grupo C. De acuerdo con Don y Avtalion (1993), las muestras de semen que son irradiadas de 0.09 a 0.27 J cm-2 muestran una deficiente condensación de la cromatina espermática y destrucción parcial de la membrana plasmática y nuclear, pero sin daño del flagelo y con reducido número de mitocondrias dañadas, lo que produce un alto potencial de fertilización de los espermatozoides.
En la segunda fase (0.3 a 0.4 J cm-2) hubo mortalidades masivas durante los periodos de incubación, probablemente por los efectos resultantes de la ruptura parcial del genoma paterno y por los efectos derivados de fragmentos grandes de dímeros de pirimidina; esto puede ocasionar la expresión inapropiada de genes paternos esenciales en los primeros estadios de desarrollo (Egami y Ijiri, 1979; Regan y Carrier, 1982; Don y Avtalion, 1993).
Adicionalmente a los efectos físicos directos de la radiación, Regan y Carrier (1982) y Don y Avtalion (1993) han señalado la posible interferencia de la cromatina, desorganizada por causa de la radiación UV, en la eficiencia de mecanismos de reparación existentes en los huevos de las especies manipuladas. Asimismo, los genes mutantes o los cromosomas anormales que se formen a consecuencia de la radiación o por una reparación incorrecta, pueden propiciar la aparición de productos letales.
Fue en la tercera fase (0.5 a 0.8 Jcm-2) en la cual se identificó un incremento en la supervivencia (fig. 3). Esto puede ser explicado sobre la base de las conclusiones de Don y Avtalion (1993). Estos autores encontraron que en dosis de 0.57 a 0.9 J cm-2, las células espermáticas de O. niloticus presentan defectos en la membrana citoplásmica y nuclear, observándose cromatina dispersa y fragmentada. La ruptura de la membrana nuclear resulta en la exposición de la cromatina de origen paterno (del esperma radiado) a ser digerida por las nucleasas en el citoplasma del huevo, resultando esto en embriones que sólo expresan el complemento cromosómico materno (no irradiado), esto es, son haploides "completos" (Egami y Ijiri, 1979; Regan y Carrier, 1982; Don y Avtalion, 1993). El aumento en la viabilidad de los embriones en esta fase (periodo de eclosión 72 h de incubación) puede entonces ser el resultado de un incremento en los porcentajes de cigotos haploides (Don y Avtalion, 1993).
La cuarta fase (0.9 y 1.0 J cm-2) (fig. 3), donde se observó la menor supervivencia en el presente estudio, puede ser explicada por el daño causado a las células espermáticas a consecuencia de la alta dosis de radiación. La motilidad fue la menor, lo que permite inferir que están totalmente lisadas y sin flagelo, de acuerdo con lo observado por Don y Avtalion (1993). Esto resultaría en muy baja eficiencia de fertilización y altas mortalidades en las primeras horas de incubación (Don y Avtalion, 1993; Li et al., 2000), y por tanto en una muy baja supervivencia.
Con respecto a las larvas que eclosionaron, la dosis de 0.7 J cm-2 produjo organismos con las características típicas del síndrome haploide, como son retraso en el desarrollo, saco vitelino distorsionado, columna vertebral doblada, cola reducida y ojos acromáticos (fig. 4c), características indicativas de la inactivación o de la destrucción total del genoma paterno (Suwa et al., 1994; Valcárcel et al., 1994). Comparando los embriones haploides, generados mediante las dosis de radiación intermedias como la de 0.7 J cm-2, con los posibles aneuploides (embriones en los que el número de cromosomas no es múltiplo exacto del juego haploide típico) probablemente generados con las dosis de radiación bajas y los huevos no fecundados de las altas, es claro que el mayor daño sobre la generación de embriones haploides está dado por las dosis de radiación bajas, ya que los embriones no sobreviven a pesar de haber ocurrido la fertilización, ni eclosionan al final del período de incubación (72 h). Por el contrario, los embriones haploides eclosionan, sobreviviendo hasta las 24 h. Estas observaciones coinciden con las presentadas por Fujioka (1993) , Sumantadinata et al. (1990), Mair (1993), Valcárcel et al. (1994), Gomelsky et al. (2000) y Galbusera et al. (2000) para otras especies de peces.
En conclusión, los resultados de supervivencia obtenidos, en los que se observa un marcado decremento con las dosis de radiación de 0.3 a 0.4 J cm-2, un incremento significativo con las de 0.5 a 0.8 J cm-2 y un nuevo decremento con la de 1.0 J cm-2, demuestran en este estudio el comportamiento clásico del efecto Hertwig, de acuerdo con Purdom (1969, 1983), Egami e Ijiri (1979), Chourrout (1982) y Valcárcel et al. (1994) .
Además, puede concluirse que estos resultados (aunados a los obtenidos con el IM) indican que la dosis óptima para la producción de larvas con las características típicas del síndrome haploide se encuentra cercana a 0.7 J cm-2, indicando nuevamente que ésta puede ser la dosis más adecuada para producir organismos ginogenéticos.
Agradecimientos
Este trabajo fue realizado con financiamiento del Centro de Investigación en Alimentación y Desarrollo, A.C., Unidad Mazatlán (proyecto fiscal 2000-6250-1) y del proyecto CONACYT J-28342B, cuyo responsable es Neil J. Duncan, a quien los autores agradecen su participación en el manejo reproductivo de los organismos. Fue también importante para el buen desempeño de este trabajo la valiosa participación del grupo de reproducción y nutrición. Se agradecen también las correcciones sugeridas por Ana María Ibarra Humphries, quien tuvo a bien revisar este documento.
Referencias
Aas, G.H., Refstie, T. and Gjerde, B. (1991). Evaluation of milt quality of Atlantic salmon. Aquaculture, 95: 125-132. [ Links ]
Abdo-de la Parra, M.I., García-Ortega, A., Martínez-Rodríguez, I., González-Rodríguez, B., Velasco, G. and Duncan, N.J. (2001). Larval rearing of the Mexican bullseye puffer Sphoeroides annulatus under hatchery conditions. Larvi 2001, 3rd Fish and Shellfish Larviculture Symposium, Gent, Belgium, 663 pp. [ Links ]
Arai, K. (2000). Chromosome manipulation in aquaculture: Recent progress and perspective. Suisanzoshoku, 48: 295-303. [ Links ]
Arias-Rodríguez, L. (2001). Inactivación genética de esperma e inducción de ginogénesis y de triploidía en el botete diana Sphoeroides annulatus (Jenyns, 1842). Tesis de maestría, Centro de Investigación en Alimentación y Desarrollo, Mazatlán, Sinaloa, México, 202 pp. [ Links ]
Billard, R. and Cosson, M.P. (1992). Some problems related to the assessment of sperm motility in freshwater fish. J. Exp. Zool., 261: 122-131. [ Links ]
Castellanos, R.S., García R., J.L., Guevara O., J.L. y Franco A., F.C. (1982). Aportación al conocimiento de la especie Sphoeroides annulatus (Jenyns). Tesis de licenciatura, Universidad Autónoma de Sinaloa, México, 42 pp. [ Links ]
Chourrout, D. (1982). Gynogenesis caused by ultraviolet irradiation of salmonid sperm. J. Exp. Zool., 223: 175-181. [ Links ]
Chourrout, D. (1984). Pressure-induced retention of second polar body and suppression of first cleavage in rainbow trout: Production of all-triploids, all-tetraploids, and heterozygous and homozygous diploid gynogenetics. Aquaculture, 36: 111-126. [ Links ]
Christensen, J.M. and Tiersch, T.R. (1994). Standardization of ultraviolet irradiation of channel catfish sperm. J. World Aquacult. Soc., 25: 571-575. [ Links ]
Cosson, M.P., Cosson, J. and Billard, R. (1991). Synchronous triggering of trout sperm is followed by an invariable set sequence of movement parameters whatever the incubation medium. Cell Motil. Cytoskel., 20: 55-68. [ Links ]
Danield, W.W. (1990). Applied Nonparametric Statistics. 2nd ed. PWS Kent Publishing Co., Boston, 635 pp. [ Links ]
Don, J. and Avtalion, R.R. (1993). Ultraviolet irradiation of tilapia spermatozoa and the Hertwig effect: Electron microscopic analysis. J. Fish Biol., 41: 1-14. [ Links ]
Duncan, N.J. and Rodríguez, M. de O.G.A. (2001). Induced spawning of the bullseye puffer Sphoeroides annulatus using LHRHa. In: The International Triennial Conference and Exposition of World Aquaculture Society. Book of abstracts. Florida, 754 pp. [ Links ]
Duncan, N.J., Rodríguez, M. de O., Alok, D. and Zohar, Y. (2003). Effects of controlled and acute injections of LHRHa on bullseye puffer fish (Sphoeroides annulatus) spawning. Aquaculture, 218: 625-635. [ Links ]
Egami, N. and Ijiri, K.-I. (1979). Effects of irradiation on germ cells and embryonic development in teleosts. Int. Rev. Cytol., 59: 195-248. [ Links ]
Fujioka, Y. (1993). Induction of gynogenetic diploids and cytological studies in Honmoroko Gnathopogon caurulescens. Nippon Suisan Gakkaishi, 59: 493-500. [ Links ]
Galbusera, P., Volckaert, F.A.M. and Ollevier, F. (2000). Gynogenesis in the African catfish Clarias gariepinus (Burchell, 1822). III. Induction of endomitosis and the presence of residual genetic variation. Aquaculture, 185: 25-42. [ Links ]
Geffen, A.J. and Evans, J.P. (2000). Sperm traits and fertilization success of male and sex-reversed female rainbow trout (Oncorhynchus mycosis). Aquaculture, 182: 61-72. [ Links ]
Gomelsky, B., Mims, S.D., Onders, R.J., Shelton, W.L., Dabrowski, K. and García-Abiado, M.A. (2000). Induced gynogenesis in black crappie. N. Am. J. Aquacult., 62: 33-41. [ Links ]
Huerta, B.M.A. (1999). Inactivación genética de esperma de cabrilla arenera Paralabrax maculatofasciatus (Steindachnert, 1868) utilizando luz UV. Tesis de maestría, Centro de Investigación en Alimentación y Desarrollo, Mazatlán, Sinaloa, México, 89 pp. [ Links ]
Hussain, M.G., Penman, D.J., McAndrew, B.J. and Johnstone, R. (1993). Suppression of first cleavage in the Nile tilapia, Oreochromis niloticus L.: A comparison of the relative effectiveness of pressure and heat shocks. Aquaculture, 111: 263-270. [ Links ]
Jenneckens, I., Müller-Belecke, A., Hörstgen-Scwark, G. and Meyers, J.-N. (1999). Proof of the successful development of Nile tilapia (Oreochromis niloticus) clones by DNA fingerprinting. Aquaculture, 173: 377-388. [ Links ]
Kanazawa, A. (1991). Puffer fish, Fugu rubripes. In: R.P. Wilson (ed.), Handbook of Nutrient Requirements of Finfish. CRC Press, Boca Raton, Florida, pp. 123-130. [ Links ]
Lasher, R. and Rugh, R. (1962). The "Hertwig effect" in teleost development. Biol. Bull., 123: 582-588. [ Links ]
Li, Q., Osada, M., Kashihara, M., Hirohashi, K. and Kijima, A. (2000). Effects of ultraviolet irradiation on genetical inactivation and morphological structure of sperm of the Japanese scallop, Patinopecten yessoensis. Aquaculture, 186: 233-242. [ Links ]
Mair, G.C. (1993). Chromosome-set manipulation in tilapia, techniques, problems and prospects. Aquaculture, 111: 227-244. [ Links ]
Martínez-Palacios, C.A., Chávez-Sánchez, M.C., Papp, G.S., Abdo-de la Parra, I. and Ross, L.G. (2002). Observations on spawning, early development and growth of the puffer fish Sphoeroides annulatus (Jenyns, 1843). J. Aquacult. Trop., 17 (1), 59-66. [ Links ]
Menkveld, R. and Kruger, T.E. (1996). Basic semen analysis. In: A.A. Acosta and T.F. Kruger (eds.), Human Spermatozoa in Assisted Reproduction. 2nd ed. Parthenon Publishing Group, New York, pp. 53-72. [ Links ]
Perchec, G., Jeulin, C., Cosson, J., André, F. and Billar, R. (1995). Relationship between sperm ATP content and motility of carp spermatozoa. J. Cell Sci., 108: 747-753. [ Links ]
Purdom, C.E. (1969). Radiation-induced gynogenesis and androgenesis in fish. Heredity, 24: 431-444. [ Links ]
Purdom, C.E. (1983). Genetics engineering by the manipulation of chromosomes. Aquaculture, 33: 287-300. [ Links ]
Regan, J.D. and Carrier, W.L. (1982). Photoreactivation in two closely related marine fishes having different longevities. Mech. Ageing Dev., 18: 59-66. [ Links ]
Satterfield, J.R. and Flickinger, S.A. (1995). Factors influencing storage potencial of preserved walleye semen. Prog. Fish-Cult., 57: 175-181. [ Links ]
Sumantadinata, K., Taniguchi, N. and Sugama, K. (1990). The necessary conditions and the use of ultraviolet irradiated sperm from different species to induce gynogenesis of Indonesian common carp. In: R. Hirano and I. Hanyu (eds.), 2nd Asian Fisheries Forum, Asian Fisheries Soc., Philippines, pp. 539-542. [ Links ]
Suwa, M., Arai, K. and Suzuki, R. (1994). Suppression of the first cleavage and cytogenetic studies on the gynogenetic loach. Fish. Sci., 60: 673-681. [ Links ]
Takai, H. and Morisawa, M. (1995). Change in intracellular K+ concentration caused by external osmolality change regulates sperm motility of marine and freshwater teleosts. J. Cell Sci., 108: 1175-1181. [ Links ]
Taniguchi, N., Han, H.S. and Tsujimura, A. (1993). Use of chromosome manipulated fish in aquaculture and related problems of conservation of wild stock. In: K.L. Main and E. Reynolds (eds.), Selective Breeding of Fishes in Asia and USA. Proc. of a workshop in Hawaii, Oceanic Institute, pp. 68-80. [ Links ]
Thorgaard, G.H. (1983). Chromosome set manipulation and sex control in fish. In: W.S. Hoar, D.J. Randall and E.M. Donaldson (eds.), Fish Physiology. Vol. IX, part B. Academic Press, New York, pp. 405-434. [ Links ]
Valcárcel, A., Guerrero, G. and Maggese, M.C. (1994). Hertwig effect caused by UV-irradiation of sperm of the catfish, Rhamdia sapo (Pisces, Pimelodidae), and its photoreactivation. Aquaculture, 128: 21-28. [ Links ]
Yamahira, K. (1997). Hatching success affects the timing of spawning by the intertidally spawning puffer Takifugu niphobles. Mar. Ecol. Prog. Ser., 155: 239-248. [ Links ]
Yamamoto, E. (1999). Studies on sex-manipulation and production of cloned populations in hirame, Paralichthys olivaceus (Temminck et Schlegel). Aquaculture, 173: 235-246. [ Links ]
Zar, J.H. (1984). Biostatistical Analysis. 2nd ed. Prentice Hall, Englewood Cliffs, New Jersey, 718 pp. [ Links ]

