










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCiencias marinas
versión impresa ISSN 0185-3880
Cienc. mar vol.30 no.1a Ensenada feb. 2004
Artículos
Dinámica de los nutrientes en el brazo oeste de Bahía San Quintín, Baja California, México, durante y después de El Niño 1997/1998
Nutrient dynamics in the west arm of San Quintín Bay, Baja California, Mexico, during and after El Niño 1997/1998
J. Martín Hernández-Ayón1, M. Salvador Galindo-Bect1, Víctor Camacho-Ibar1, Zaul García-Esquivel1* Marco A. González-Gómez1 y Francisco Ley-Lou2
1 Instituto de Investigaciones Oceanológicas, Universidad Autónoma de Baja California Km. 103 carretera Tijuana-Ensenada Ensenada, CP 22860, Baja California, México. *E-mail: sgarcia@uabc.mx
2 Facultad de Ciencias Marinas, Universidad Autónoma de Baja California Km. 103 carretera Tijuana-Ensenada Ensenada, CP 22860, Baja California, México.
Recibido en julio 2002;
Aceptado en agosto de 2003.
Resumen
Se realizaron series de tiempo de 8 h (mayo, septiembre y noviembre de 1997, y Marzo de 1998) y de 24 h (junio y septiembre de 1999, y febrero de 2000) en dos (1997-1998) y tres localidades (1999-2000) del brazo oeste de Bahía San Quintín (Bahía Falsa, BF), con el fin de describir la dinámica de los nutrientes durante y después del evento El Niño 1997/1998. La concentración de clorofila a también se evaluó en cada serie. Se estimaron los flujos no conservativos de fósforo inorgánico disuelto (ΔPID), nitrógeno inorgánico disuelto (ΔNID), nitratos (ΔNO3-) y amonio (ΔNH4+), utilizando un modelo de balance de nutrientes. Durante El Niño 1997/1998 se detectaron pulsos de NH4+ entre 18 y 45 µM, y concentraciones medias más altas que en 1999-2000. La concentración de NH4+ fue consistentemente menor de 8 µM en 1999-2000. Se observaron correlaciones significativas (r = 0.60 a 0.80) entre la altura de la marea y la concentración de NO3- en 1999-2000, y una relación inversa entre NO3- y salinidad, lo que reflejó un aporte oceánico de NID nuevo. La concentración de clorofila a fue hasta dos veces más alta en 1999-2000 que durante El Niño. No se observaron correlaciones significativas entre la marea y la concentración de nutrientes en 1997-1998, lo que se sugiere que: (a) las concentraciones altas de NH4+ en 1997-1998 se debieron a la remineralización in situ de la materia orgánica y posiblemente a la excreción de moluscos bivalvos presentes en BF, y (b) el aumento de clorofila a y NO3-observados en 1999-2000 se debieron principalmente a aportes nuevos desde el océano adyacente. La combinación de valores positivos de ΔPID (0.1 a 0.57) y negativos del metabolismo neto del ecosistema (MNE = -1 a -60 mmol m-2 d-1) durante 1997-2000, indican que BF exportó permanentemente PID hacia el océano y actuó como un sistema heterótrofo neto en presencia y ausencia de El Niño.
Palabras clave: Bahía San Quintín, surgencias, nutrientes, balances, El Niño, metabolismo neto.
Abstract
Time series of 8 h (May, September and November 1997, and March 1998) and 24 h (June and September 1999, and February 2000) were carried out at two (1997-1998) or three sites (1999-2000) of the west arm (Falsa Bay, FB) of San Quintín Bay, in order to assess the nutrient dynamics during and after the 1997/1998 El Niño event. Chlorophyll a concentrations were also deter- mined during each time series. Non-conservative fluxes of dissolved inorganic phosphorus (ΔDIP), dissolved inorganic nitrogen (ADIN), nitrates (ΔNO3-) and ammonium (ΔNH4+) were estimated using a nutrient budget model. High NH4+ peaks (18-45 µM) were detected during El Niño 1997/1998, and the mean concentration values were higher than those of 19992000. Ammonium concentration was consistently less than 8 µM during 1999-2000. Significant correlations (r = 0.60 to 0.80) were found between tidal height and NO3- concentration in 1999-2000, as well as an inverse relationship between nitrates and salinity, thus reflecting the input of new DIN from the ocean. Chlorophyll a concentration was up to two times higher in 19992000 than during El Niño. There were no significant correlations between tidal height and nutrient concentration during 19971998, suggesting that: (a) the high NH4+ concentrations found in 1997-1998 were due to in situ remineralization of the organic matter and, possibly, the excretion by cultured mollusc bivalves in FB, and (b) the increase of chlorophyll a and NO3- during 1999-2000 was mainly due to new inputs from the adjacent ocean. The positive ΔDIP values (0.1 to 0.57) and negative values of the net ecosystem metabolism (NEM = -1 to -60 mmol m-2 d-1) observed throughout 1997-2000 indicate that FB permanently exported DIP towards the ocean and behaved as a net heterotrophic system, with or without the presence of El Niño.
Key words: San Quintín Bay, upwelling, nutrients, budget, El Niño, net metabolism.
Introducción
El Sistema de la Corriente de California (SCC) se extiende sobre las costas de California (EUA) y Baja California (México), y controla en gran medida el funcionamiento de las surgencias y la oceanografía de la región (Ibarra-Obando et al., 2001). El SCC es sensible a los cambios climatológicos interanuales tales como los generados por El Niño (Schwing et al., 1997). El impacto de El Niño sobre el SCC incluye la reducción en la intensidad de las surgencias, el calentamiento de la capa de mezcla, el hundimiento de la nutriclina y el aumento en el nivel del mar (Lynn et al., 1995). En Bahía San Quintín se ha reportado la reducción en la concentración de nutrientes durante el evento de El Niño 1982/1983 (Torres-Moye y Álvarez-Borrego, 1985), así como de la productividad orgánica primaria (Torres-Moye y Álvarez-Borrego, 1985; Silva-Cota y Álvarez-Borrego, 1988). Recientemente se documentó también la disminución de biomasa en las poblaciones de macroalgas de las costas de Baja California durante El Niño 1997/1998 (Ladah et al., 1999).
A diferencia de otros eventos El Niño (e.g., 1982/1983), el de 1997/1998 se presentó más temprano de lo normal en las costas de Baja California. Imágenes de satélite mostraron que las anomalías positivas de temperatura superficial del océano adyacente iniciaron entre los meses de abril y julio de 1997, y tuvieron registros máximos en enero de 1998 (Durazo y Baumgartner, 2002). Asimismo, Ladah et al. (1999) reportaron el inicio de las anomalías positivas de temperatura en Bahía Tortugas, Baja California (~27.6°N), a partir de mayo de 1997. En julio de 1997 se observaron anomalías mayores de +1°C que persistieron por mas de ocho meses (Ladah et al., 1999). De acuerdo con Durazo y Baumgartner (2002), durante El Niño 1997/1998 se reportó agua más caliente y salada en los primeros 600 m de profundidad en las costas de Baja California, y el regreso a condiciones no Niño ocurrió entre julio y octubre de 1998.
Aunque se sabe que los eventos El Niño inducen cambios en las condiciones oceanográficas en las aguas costeras que interactúan con Bahía San Quintín, hasta ahora no existen estudios que describan los cambios interanuales en la dinámica de los nutrientes dentro de esta laguna costera. El presente estudio tuvo como objetivos: (a) identificar la variación, en periodos cortos, de la concentración de nutrientes en el brazo oeste (Bahía Falsa, BF) de Bahía San Quintín durante y después de un evento El Niño; (b) comparar los flujos no conservativos de nutrientes, calculados para ambas condiciones oceanográficas; y (c) comparar el metabolismo neto del sistema (MNE) para BF durante y después de un evento El Niño. Para ello, se midieron los cambios en la concentración de nutrientes en la columna de agua a partir de siete series de tiempo cortas (8 ó 24 h) realizadas durante el periodo 1997-2000 (97-00).
Materiales y métodos
En mayo, septiembre y noviembre de 1997, y marzo de 1998, se realizaron series de tiempo de 8 h en dos localidades de BF (sitios C y B, fig. 1), mientras que en junio y septiembre de 1999 y febrero de 2000 se realizaron series de 24 h en tres localidades (sitios C, B y Y, fig. 1). En cada sitio se recolectaron muestras superficiales de agua cada hora (1997-1998) o cada 2 h (1999-2000), con el fin de cuantificar las concentraciones de clorofila a y nutrientes (NO3-, NH4+ y PO4-3). Se filtraron muestras de 200 mL de agua, in situ, a través de filtros de fibra de vidrio GF/F y cada filtro se congeló para la determinación posterior de clorofila a, en el laboratorio. El agua del filtrado se recolectó en frascos de vidrio y se congeló para el posterior análisis de los nutrientes. La temperatura y la salinidad superficiales se registraron en forma continua con un CTD Sea Bird durante los muestreos de 1997-1998, mientras que en 1999-2000 las mediciones se realizaron con un salinómetro-conductímetro. A partir de marzo de 1997 se instaló en la estación B un termógrafo (± 0.2°C) construído en el Instituto de Investigaciones Oceanológicas (López, 1992), el cual se reemplazó en 2000 por un termistor submarino portátil de igual precisión (Onset Computer Corp.). Los termógrafos se instalaron a 1 m de profundidad y se programaron con una frecuencia de registro de 30 min. Los datos obtenidos con los termógrafos fueron tratados de acuerdo con el procedimiento estándar indicado por los fabricantes. Las clorofilas fueron extraídas en el laboratorio con acetona al 90% y analizadas mediante fluorometría (Strickland y Parsons, 1972), utilizando un fluorómetro Turner 112. La concentración de nutrientes se determinó con un analizador de flujo contínuo Lachat QuikChem 8000 en la Universidad de California en Santa Barbara. La precisión para cada uno de los análisis fue de ±0.05, ±0.03 y ±0.02 µM para NO3-, NH4+ y PO4-3, respectivamente. Por cada diez muestras se analizó un estándar de la curva de calibración para asegurar la calidad de las mediciones.

Con los datos de salinidad y nutrientes se calculó el metabolismo neto de BF utilizando el modelo de cajas propuesto por Gordon et al. (1996) y descrito para Bahía San Quintín por Camacho-Ibar et al. (1999). Las variables del modelo fueron el volumen del sistema (BF), los flujos de agua dulce aportados por ríos (VQ), agua subterránea (VG), precipitación pluvial (VP) y evaporación (VE). Se requirieron además los aportes de fósforo inorgánico disuelto (PID) y nitrógeno inorgánico disuelto (NID) asociados con los flujos de agua dulce, así como los gradientes de salinidad, PID y NID, entre el sistema y el mar adyacente. El área de BF se consideró de 9 x 106 m2, con una profundidad media de 1 m. Se consideró un valor promedio de VE = 36 x 103 m3 d-1 y un valor de VP = 0.9 x 103 m3 d-1, restringido este último a los meses de invierno (Camacho-Ibar et al., 1999). Para VQ y VG se consideraron valores de cero, ya que durante el periodo de muestreos no hubo escurrimientos superficiales ni probablemente tampoco aportes subterráneos (Aguirre-Muñoz et al., 2001).
Los balances de materiales conservativos (agua y sal) y no conservativos (nutrientes) y la estimación del metabolismo neto del ecosistema (MNE) se realizaron en tres pasos:
(a) Para el balance de agua y sal se supuso que el volumen de BF permaneció constante a escalas mayores de un ciclo de marea (dVSyst/dt = 0 = VQ + VP + VE + VG + VO +VR), de tal forma que el volumen residual (VR) compensó las pérdidas o ganancias netas de agua dulce, de acuerdo con la ecuación:
Se consideró además que las sales disueltas fueron removidas o adicionadas a BF solamente por procesos de mezcla (VX) y advección (VR) de acuerdo con la ecuación (Smith y Hollibaugh, 1997):
donde SOcn es la salinidad del océano y SSyst la salinidad del sistema (BF). Los datos obtenidos durante los flujos de marea en la estación B se consideraron como valores "oceánicos" para 1997-1998, mientras que para 1999-2000 se utilizó el promedio de los valores obtenidos en las estaciones B y Y durante los flujos de marea. La estación C se utilizó para referir la salinidad del sistema. Con la información anterior, se estimó el tiempo de residencia (τ), mediante la ecuación:
(b) Para estimar el flujo no conservativo de nutrientes se utilizó una ecuación similar a la del balance de agua y sal, excepto que se incluyó el término no conservativo (ΔY), el cual indica si el sistema es fuente o sumidero neto de estos materiales:
(c) El MNE (produccion - respiración) y el balance entre la fijación de nitrógeno y la desnitrificación (nfix-denit) se estimaron estequiométricamente a partir de los balances de PID y de NID, suponiendo una relación de Redfield para la razón carbono:fósforo (C:P) = 106:1, y para la razón nitrógeno:fósforo (N:P) = 16:1, como se indica a continuación:
donde ΔNIDobs es el flujo no conservativo observado de NID, ΔNIDexp es el flujo esperado no conservativo de NID calculado con la relación de Redfield (= 16 x ΔPID), y ΔPID es el flujo no conservativo observado de PID.


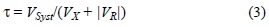


Resultados
Variables conservativas
La salinidad en la cabeza de BF fue generalmente mayor que en la parte externa, con un valor promedio para todos los muestreos de 34.6 y 34.2, respectivamente (tabla 1). El tiempo de residencia del agua calculado para la mayoría de los muestreos fluctuó entre 1 y 6 días, con excepción de noviembre de 1997 y febrero de 2000 cuando el tiempo de residencia calculado fue de alrededor de 11 días (tabla 1).

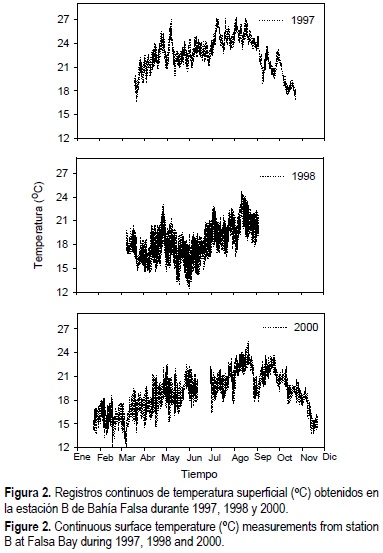
Las temperaturas más altas de todos los muestreos se registraron en 1997, con valores máximos de 27.4°C y mínimos de 16.6°C (fig. 2). La máxima temperatura registrada para el periodo de abril a agosto se observó en 1997 (27.3°C), seguida de las de 2000 (25.6°C) y de 1998 (24.7°C), mientras que las temperaturas mínimas de ese mismo periodo fueron 16.6°C, 14.6°C y 12.3°C, respectivamente. Los valores mínimos de temperatura se presentaron con las surgencias de primavera (mayo a junio) de 1998 y marzo de 2000, con registros mínimos de 12.3°C y 12.0°C, respectivamente. En general, durante los tres años se observaron pulsos de temperaturas bajas con una frecuencia aproximada de 15 días, los cuales están probablemente relacionados con eventos de surgencias.
Variables no conservativas
En las series de 1997-1998 no se observó una relación clara entre la altura de la marea y la concentración de nutrientes (fig. 3), excepto para el amonio que presentó picos máximos mayores de 25 µM en las series de primavera (mayo de 1997 y marzo de 1998), durante la fase de velocidad máxima de la corriente de marea (fig. 3 a-b, g-h). El PO4-3 y el NO3- también tendieron a mostrar picos máximos durante el flujo y reflujo de las series, aunque con menor consistencia (fig. 3). En contraste, durante los tres muestreos del periodo 1999-2000 se presentaron correlaciones positivas significativas (P < 0.05) entre la altura de la marea y la concentracion de nitratos en la estación Y, con valores de coeficiente de correlación (r) entre 0.68 y 0.87 (fig. 4, tabla 2). La clara covariación entre la altura de la marea y los nitratos observada en las series de la estación Y, y en una serie de la estación B en febrero de 2000 (fig. 4) indica la ocurrencia de aportes de NO3-desde el océano hacia el interior de la bahía durante el flujo de marea en años no Niño. Al igual que los nitratos, la clorofila a mostró valores muy bajos de r con relación a la altura de la marea durante 1997-1998, mientras que en 1999-2000 se presentaron varios valores significativos (tabla 2). En general, los valores promedio de clorofila a fueron menores en 1997-1998 (aprox. 1 mg m-3) que en 1999-2000 (>2 mg m-3). Es de notar que la concentración de clorofila a fue generalmente mayor en la boca (hasta dos veces mayor) que en la parte interna (fig. 5).

El PID mostró de manera consistente valores más altos en la estación C, con valores promedio entre 1.1 y 5.3 µM (tabla 3). En la boca de BF los valores promedio fluctuaron entre 0.6 y 1.9 µM. El único muestreo en el cual no se observó un gradiente claro de PID fue el de septiembre de 1999 (tabla 3). Durante el periodo 1997-1998 se detectaron mayores concentraciones medias de NH4+ (4 a 17 µM) que en 19992000 (1.8 y 4.6 µM). En 1997-1998 los valores más altos se observa- ron generalmente hacia el interior de la bahía, mientras que en 1999-2000 no se observó un gradiente claro entre sitios de muestreo (tabla 3). Por otra parte, durante 1997-1998 la concentración media de NO3- fue generalmente más alta en la cabeza (1.0 a 3.3 µM) que en la boca de BF (0.8 a 2.0 µM), pero dicho patrón se invirtió en 1999-2000 (tabla 3). Es notable que al igual que el PO4-3 y el NH4+, la concentración media de NO3- fue similar en ambas localidades en septiembre de 1999 (fig. 4, tabla 3).
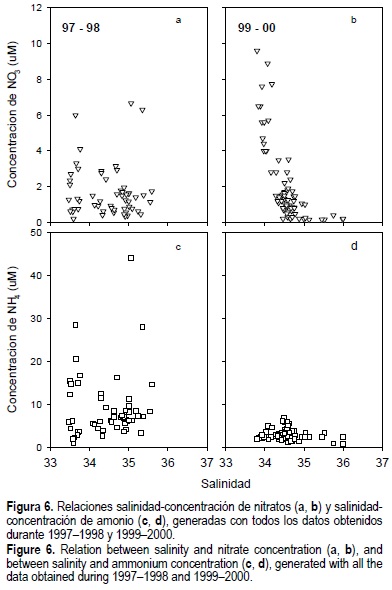
Durante el periodo 1997-1998 no se observó una relación clara entre la concentración de las especies nitrogenadas y la salinidad (fig. 6a-d), aunque hubo una tendencia del amonio a aumentar con el incremento de salinidad a concentraciones menores de 10 µM (fig. 6c). En contraste, los datos para junio de 1999 y febrero de 2000 mostraron una alta correlación inversa (r = -0.95) entre NO3- y salinidad (fig. 6b).
Todos los valores de ΔPID calculados fueron positivos (+0.01 a +0.57 mmol m-2 d-1), lo cual indica que el sistema produjo PID y que pudo funcionar como una fuente para el océano (tabla 4). En consecuencia, los valores del MNE fueron negativos (-1 a -60 mmol m-2 d-1), lo que indica que BF se comportó como un sistema heterótrofo neto en todas las fechas en las que se realizaron los muestreos. Por otra parte, todos los valores de ΔNID fueron positivos durante el periodo 1997-1998 (+0.14 a +3.71 mmol m-2 d-1), indicando que el sistema exportó NID al océano durante la condición El Niño. En contraste, los valores de ΔNID calculados para 1999-2000 fueron negativos (entre -0.30 y -0.80 mmol m-2 d-1), indicando que el sistema importó NID durante ese periodo (tabla 4). Los valores de (nfix-denit) fueron negativos en todas las series (-0.5 a -6.9 mmol N m-2 d-1), indicando que BF actuó como un sumidero de nitrógeno tanto en condiciones de El Niño como en condiciones no Niño.

Para comprobar que los cálculos de los balances de nutrientes no estuvieran afectados por promediar observaciones de nutrientes en flujo y reflujo de marea, se calcularon los flujos no conservativos para ambas condiciones por separado. Los datos en la tabla 5 muestran que, para cada serie, el signo de los flujos no conservativos fue similar durante el flujo y reflujo de la marea. La tabla 5 también muestra que, con excepción de la serie de septiembre de 1997, el flujo no conservativo de NID estuvo dominado por los aportes internos de NH4+, presentándose valores de ΔNH4+ entre +0.17 y +5.91 mmol m-2 d-1. En contraste, los flujos no conservativos de NID en 1999-2000 fueron ligeramente dominados por el consumo de NO3-, con valores de balances de ΔNO3 entre -0.28 y -0.68 mmol m-2 d-1 (tabla 5).

Discusión
Las altas concentraciones de NID, principalmente de amonio, observadas en 1997-1998, combinadas con la falta de correlación entre las especies nitrogenadas y la salinidad o la altura de la marea, sugieren que durante el evento El Niño la concentracion de NID en BF fue controlada por procesos biogeoquímicos in situ más que por aportes físicos externos. Esto incluyó la remineralización de la materia orgánica, y la excreción de amonio por parte de ostiones y almejas presentes en BF. Los valores consistentemente negativos de (nfix-denit) hallados en el presente estudio (tabla 4) sugieren que BF actuó como un sumidero neto de nitrógeno en presencia y ausencia de El Niño, y que la desnitrificación fue un proceso que ayudó a controlar la disponibilidad de NO3-. Anteriormente se reportaron concentraciones de NH4+ por arriba de 1900 µM en aguas intersticiales de sedimentos someros de Bahía San Quintín (Camacho-Ibar y Álvarez-Borrego, 1988). La dominancia de los aportes de NID por remineralización en 1997-1998 sería análogo a lo reportado para la Bahía de Monterey durante El Niño 1992/1993, donde se observó una disminución substancial de la producción nueva y el aumento de nutrientes regenerados (Kudela y Chávez, 2000). El empobrecimiento de la concentración de nitratos en las aguas costeras frente a Bahía San Quintín durante el periodo de El Niño 1997/1998 es consistente con las anomalías negativas en la concentración de nitratos (hasta de -5 µM) reportadas para la zona central de la costa de California (Chávez et al., 2002). En contraste, la alta correlación positiva del nitrato con la marea en 1999-2000 (tabla 2), y su relación inversa con la salinidad, reflejaron el aporte oceánico de nitratos y producción nueva hacia el interior de la bahía. Anteriormente, Farfán y Álvarez-Borrego (1983) observaron que los flujos de NID desde el océano hacia Bahía San Quintín estaban asociados con la intensidad de las surgencias. Pennington y Chávez (2000) también reportaron valores más altos de nitratos en aguas superficiales costeras de la Bahía de Monterey, del orden de 7.5 a 20 µM durante mayo y junio, periodo en el cual las surgencias fueron más intensas. Aunque se ha reportado un calentamiento de las aguas locales del SCC y una disminución en las surgencias costeras durante El Niño (Durazo y Baumgartner, 2002), los mínimos periódicos de temperatura observados en el presente estudio sugieren que los eventos de surgencias estuvieron presentes en las costas de Bahía San Quintín aun durante El Niño. Sin embargo, el agua subsuperficial que entró a BF durante ese periodo era cálida y pobre en nitratos, mientras que en 1999-2000 entró agua más rica (fig. 6a, b). Podría suponerse que las diferencias en la concentración de NID entre los periodos 1997-1998 y 1999-2000 se debieron a que los muestreos se llevaron a cabo durante etapas diferentes de eventos de surgencias. Sin embargo, el análisis del registro contínuo de temperaturas, verificado con el índice de Bakun (http://upwell.pfeg.noaa.gov) correspondiente a cada muestreo, indicó que sólo el muestreo de noviembre de 1997 se realizó en ausencia de surgencias; los de septiembre de 1997 y marzo de 1998 se ubicaron en la etapa de relajación de la surgencia, y el de mayo de 97 se realizó durante una surgencia fuerte. Por otro lado, los muestreos del 99-00 se realizaron en las etapas de relajación y de surgencia fuerte, en las cuales se pudieron observar pulsos de nitratos, de acuerdo con lo esperado para condiciones distintas a El Niño (fig. 4). La falta de pulsos de nitratos durante la fase más intensa de las surgencias de 1997-1998 refuerzan la hipótesis de que durante esos años los flujos de NID se dieron principalmente a partir de la remineralización en el interior de BF.
En el presente estudio no se evaluó la contribución de amonio por excreción de moluscos bivalvos. Sin embargo, se ha estimado que en primavera y otoño el ostión Crassostrea gigas cultivado en la Bahía de Morlaix, Francia, excreta entre 2.5 y 6.7 µmoles NH4+ g-1 tejido seco h-1 (Boucher y Boucher-Rodoni, 1988). Aun cuando la biomasa del ostión C. gigas cultivado en BF disminuyó hasta en un 60% debido a mortalidades inusuales ocurridas en 1998, durante ese periodo se introdujeron almejas del Golfo de California (García-Esquivel et al., este volumen), alcanzando una biomasa combinada de ostiones y almejas de aproximadamente 2000 t (Aguirre-Muñoz et al., 2001; Camacho-Ibar et al., 2003). Utilizando las relaciones alométricas reportadas por García-Esquivel et al. (este volumen), y suponiendo que las 2000 t fueron ostiones, se estima que la biomasa anterior equivale a 38 x 106 ostiones adultos. Si consideramos una excreción de 4.6 µmoles NH4 g-1 tejido seco h-1, en promedio, y un peso promedio de 1.9 g de tejido seco para ostiones de talla comercial cultivados en BF (Garcia-Esquivel et al., este volumen), lo anterior se traduce en aproximadamente 210 µmoles NH4 excretados por ostión por día, o 7.98 x 106 mmoles d-1 para el total de ostiones/almejas presentes en BF. Al dividir el valor anterior entre la superficie de BF (9 x 106 m2) encontramos que los moluscos presentes durante el periodo 1997-1998 podrían haber excretado hasta 0.9 mmol NH4 m2 d-1. Este valor representa entre el 24% y >100% de los flujos observados de NID para ese periodo (tablas 4, 5), lo cual sugiere que la excreción por moluscos bivalvos es un componente que debe ser considerado en los balances de nitrógeno de BF.
El gradiente de clorofila a, entre la boca y la cabeza de BF se mantuvo aun en presencia de El Niño, por lo cual es de suponer que el tiempo de residencia del agua y/o el pastoreo jugaron un papel crítico sobre la variabilidad espacial de la materia orgánica particulada en BF, especialmente durante El Niño, cuando la concentración media de clorofila importada del oceáno se redujo en más de la mitad al interior de BF (fig. 5a). Los tiempos de residencia del agua estimados para BF (tabla 1) a partir de la salinidad fueron similares a los reportados por Juárez-Villarreal (1982) y la tendencia estacional también coincidió con lo reportado para toda la Bahía San Quintín (Camacho-Ibar et al., 2003). Anteriormente, estos autores habían sugerido que los largos tiempos de residencia en invierno se debían a la menor intensidad de los vientos del noroeste durante esa época del año. Aun cuando el papel del viento sobre el tiempo de residencia no se pudo confirmar, los resultados del presente estudio mostraron que la evacuación del agua es consistentemente más dinámica durante los meses de primavera y verano.
Balance de nutrientes
Los valores positivos de ΔPID estimados para BF durante y después del periodo de El Niño (tabla 4) indican que BF exportó permanentemente PID hacia el océano. Álvarez-Borrego y Chee-Barragán (1976) y Zertuche-González y Álvarez-Borrego (1978) reportaron la misma tendencia en estudios previos, pero ésta es la primera ocasión que se incluyen muestreos consecutivos durante y después de El Niño. La magnitud de los flujos no conservativos de PID encontrados en el presente estudio (+0.01 a +0.57 mmol m-2 d-1) son consistentes con los valores reportados para la misma zona por Camacho-Ibar et al. (2003) y con los flujos reportados para Bahía Tomales, California (Smith y Hollibaugh, 1997).
Los cálculos de ΔPID obtenidos con el modelo de LOICZ, traducidos a valores de MNE, indican que BF se comportó como un sistema heterotrófico neto, con valores más altos en verano y bajos en invierno (tabla 4). El hecho de que esta condición heterotrófica neta se haya mantenido en presencia de El Niño sugiere que durante ese periodo existió aporte externo de materia orgánica desde la boca hacia BF en cantidades suficientes para que la respiración (r) excediera la fotosíntesis (p). Esto no parece ser una condición inusual, ya que Smith y Hollibaugh (1997) reportaron que en la Bahía Tomales el MNE no presentó valores anómalos durante el evento de El Niño 1992/1993, y éstos se mantuvieron en el intervalo de 0 a -30 mmol C m-2 d-1 a lo largo de 8 años de observaciones, con excepción de algunos valores extremos que alcanzaron -60 mmol C m-2 d-1. En BF, todos los valores calculados hasta ahora se mantienen entre -1 y -28 mmol m-2 d-1, con excepción de un valor extremo de -60 mmol m-2 d-1 (tabla 4). Los resultados del cálculo del MNE para junio de 1999 indican que la fotosíntesis y la respiración en BF estuvieron casi balanceadas, lo cual podría considerarse como anómalo si consideramos que otros autores reportaron mayores valores de MNE en primavera y verano que en invierno (Smith y Hollibaugh, 1997; Camacho-Ibar et al., 2003). No obstante, el gradiente de concentración de nitratos observado entre la cabeza y la boca de BF para esa fecha sugieren que hubo un consumo intenso por parte de productores primarios. En dicho muestreo se observaron, en general, las concentraciones más altas de clorofila a (> 2 mg m3), incluso en el interior de BF (fig. 5b). Estas observaciones sugieren que durante este periodo se presentó un evento de alta producción primaria, lo cual balanceó los valores comúnmente elevados de respiración resultando en un valor de MNE cercano al cero.
En resumen, los resultados del presente estudio indican que BF es un sistema heterotrófico neto, tanto en presencia como en ausencia del fenómeno El Niño, por lo que exporta permanentemente PID que se produce internamente a partir de la remineralización de materia orgánica importada. Durante El Niño 1997/1998 hubo fuertes pulsos de amonio en la columna de agua de BF, mayores que los que ocurrieron en el periodo no Niño. Se sugiere que dicha acumulación se debió a a la remineralización bentónica de la materia orgánica y a la excreción de ostiones y almejas presentes en BF. Aunque los procesos de generación interna de amonio siguieron presentes durante los años no Niño, el agua del mar adyacente abasteció de nitrógeno nuevo a Bahía San Quintín con cada flujo de la marea. Este abasto promovió la producción nueva en el interior de la laguna (evidenciada por las mayores concentraciones de clorofila a en 1999-2000), favoreciendo el consumo biológico del NID y limitando la acumulación del mismo en la columna de agua.
Agradecimientos
El presente estudio fue realizado con financiamiento del Sistema de Investigación del Mar de Cortés, a través del proyecto Estimación del Potencial Productivo y Capacidad de Carga Ostrícola en Bahía San Quintín, Baja California (ref. SIMAC 980106020). Se agradece el apoyo logístico y en especie de la Empresa Integradora Acuacultores de San Quintín, S.A. de C.V. Nuestro agradecimiento especial a Conrado Rivera, Dulce Partida, Jesus Serrano y Sergio Jiménez por su ayuda durante los muestreos. El manuscrito mejoró notablemente con las sugerencias de Francisco Delgadillo-Hinojosa y otro revisor anónimo.
Referencias
Aguirre-Muñoz, A., Buddemier, R.W., Camacho-Ibar, V., Carriquiry, J.D., Ibarra-Obando, S.E., Massey, B.W., Smith S.V. and Wulff, F. (2001). Sustainability of coastal resource use in San Quintín, Mexico. AMBIO, 30: 142-149. [ Links ]
Álvarez-Borrego, S. y Chee-Barragán, A. (1976). Distribución superficial de fosfatos y silicatos en Bahía de San Quintín, BC. Cienc. Mar., 3: 51-61. [ Links ]
Boucher, G. and Boucher-Rodoni, R. (1988). In situ measurement of respiratory metabolism and nitrogen fluxes at the interface of oyster beds. Mar. Ecol. Prog. Ser., 44: 229-238. [ Links ]
Camacho-Ibar, V.F. and Álvarez-Borrego, S. (1988). Nutrient concentration in pore waters of intertidal sediments in a coastal lagoon: Patchiness and temporal variations. Sci. Total Environ., 75: 325-339. [ Links ]
Camacho-Ibar, V.F., Carriquiry, J.D. and Smith, S.V. (1999). Bahía San Quintín, Baja California: N/P budgets within compartments in a coastal lagoon. In: Mexican and Central American Coastal Lagoon Systems: Carbon, Nitrogen and Phosphorus Fluxes (Regional Workshop II). LOICZ Reports and Studies No. 13, LOICZ, Texel, Netherlands, pp. 65-78. [ Links ]
Camacho-Ibar, V.F., Carriquiry, J.D. and Smith, S.V. (2003). Non-conservative P and N fluxes and net ecosystem production in San Quintín Bay, Mexico. Estuaries, 26(5): 1220-1237. [ Links ]
Chávez, F.P., Pennington, J.T., Castro, C.G., Ryan, J.P., Michisaki, R.P., Schlining, B., Walz, P., Buck, K.R., McFadyen, A. and Collins, C. (2002). Biological and chemical consequences of the 1997-1998 El Niño in central California waters. Prog. Oceanogr., 54: 205-232. [ Links ]
Durazo, R. and Baumgartner, T.R. (2002). Evolution of oceanographic conditions off Baja California: 1997-1999. Prog. Oceanogr., 54: 7-73. [ Links ]
Farfán, B.C. and Álvarez-Borrego, S. (1983). Variablility and fluxes of nitrogen and organic carbon at the mouth of a coastal lagoon. Estuar. Coast. Shelf Sci., 17: 599-612. [ Links ]
García-Esquivel, Z., González-Gómez, M.A., Ley-Lou, F. y Mejía-Trejo, A. (2003). Potencial ostrícola del brazo oeste de Bahía San Quintín: Biomasa actual y estimación preliminar de la capacidad de carga. Cienc. Mar., 30(1): 71-84. [ Links ]
Gordon, D.C.J.R., Boudreau, P.R., Mann, K.H., Ong, J.E., Silvert, W.L., Smith, S.V, Wattayakorn, G., Wulff, F. and Yanagi, T. (1996). LOICZ Biogeochemical Modelling Guidelines. LOICZ Report and Studies No. 5. 2nd ed. LOICZ, Texel, Netherlands, vi + 96 pp. [ Links ]
Ibarra-Obando, S.E., Camacho-Ibar, V.F., Carriquiry, J.D. and Smith, S.V. (2001). Upwelling and lagoonal ecosystems of the dry Pacific coast of Baja California. In: U. Seeliger and B. Kjerfve (eds.), Coastal Marine Ecosystems of Latin America. Ecological Studies. Vol. 144. Springer-Verlag, Berlin, pp. 315-330. [ Links ]
Juárez-Villarreal, M.M. (1982). Modelo de dispersión para un estuario ramificado y su aplicación a Bahía de San Quintín. Tesis de maestría, Centro de Investigación Científica y Educación Superior de Ensenada, Baja California, 79 pp + apéndice. [ Links ]
Kudela, R.M. and Chávez, F.P. (2000). Modeling the impact of the 1992 El Niño on new production in Monterey Bay, California. Deep-Sea Res., II 47: 1055-1076. [ Links ]
Ladah, L.B., Zertuche-González, J.A. and Hernández-Carmona (1999). Giant kelp (Macrocystis pyrifera, Phaeophyta) recruitment near its southern limit in Baja California after mass disappearance during ENSO 1997-1998. J. Phycol., 35: 1106-1112. [ Links ]
López, O.R. (1992). Termógrafo Digital Submarino. Rev. Instrumentación y Desarrollo, SOMI, 3, 2. [ Links ]
Lynn R., Schwing, F.B. and Hayward, T.L. (1995). The effect of the ENSO on the California Current System. CalCOFI Rep., 36: 57-71. [ Links ]
Pennington, J.T and Chávez, F.P. (2000). Seasonal fluctuations of temperature, salinity, nitrate, chlorophyll and primary production at station H3/M1 over 1989-1996 in Monterey Bay, California. Deep-Sea Res., 47: 947-973. [ Links ]
Schwing, F.B., Hayward, T.L., Sakuma, K.M., Murphree, T., Mascarenhas, A.S., Larios-Castillo, S.I., Mantyla, A.W., Cummings, S.L., Chávez, F.P., Baltz, K. and Ainley, D.G. (1997). The state of the California Current, 1996-1997: Mixed signals from the tropics. CalCOFI Rep., 38: 22-47. [ Links ]
Silva-Cota, S. and Álvarez-Borrego, S. (1988). The "El Niño" effect on the phytoplankton of a northwestern Baja California coastal lagoon. Estuar. Coast. Shelf Sci., 27: 109-115. [ Links ]
Smith, S.V. and Hollibaugh, J.T. (1997). Annual cycle and interannual variability of ecosystem metabolism in a temperate climate embayment. Ecol. Monogr., 67: 509-533. [ Links ]
Strickand, J.D. and Parsons, T.R. (1972). A Practical Handbook of Seawater Analysis. 2nd ed. Bull. Fish. Res. Bd. Canada, 167, 311 pp. [ Links ]
Torres-Moye, G. y Álvarez-Borrego, S. (1985). Efectos de El Niño en los nutrientes y el fitoplancton de verano de 1983, en aguas costeras de Baja California occidental. Cienc. Mar., 11(3): 107-113. [ Links ]
Zertuche-González, J.A. y Álvarez-Borrego, S. (1978). Series de tiempo de variables fisicoquímicas en las bocas de dos antiestuarios de Baja California. Cienc. Mar., 5(1): 91-103. [ Links ]

