










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkCiencias marinas
versão impressa ISSN 0185-3880
Cienc. mar vol.30 no.1a Ensenada Fev. 2004
Artículos
Distribución, movimientos y tamaño de grupo del tursión (Tursiops truncatus) al sur de Bahía San Quintín, Baja California, México
Distribution, movements and group size of the bottlenose dolphin (Tursiops truncatus) to the south of San Quintín Bay, Baja California, Mexico
E. Morteo1, 2 3*, G. Heckel1, 3, R.H. Defran1, 4 y Y. Schramm1, 2
1 Investigación y Conservación de Mamíferos Marinos de Ensenada, A.C. (ICMME). Plácido Mata No. 2309, Departamento D-5 Condominios Las Fincas Ensenada, CP 22810, Baja California, México. *E-mail: e_morteo@hotmail.com
2 Facultad de Ciencias Marinas, Universidad Autónoma de Baja California. Km 103 carretera Tijuana-Ensenada Ensenada, CP 22860, México.
3 Centro de Investigación Científica y de Educación Superior de Ensenada. Km 107 carretera Tijuana-Ensenada, Ensenada, CP 22860, México.
4 Department of Psychology, San Diego State University (SDSU). San Diego, CA 92182, USA.
Recibido en octubre de 2002;
Aceptado en mayo de 2003.
Resumen
Entre julio de 1999 y junio de 2000 se realizaron 12 navegaciones de fotoidentificación a lo largo de la costa al sur de Bahía San Quintín, Baja California, México. Se recorrieron en total 276.8 km de costa, en 31.7 h. Durante 12.9 h de observación se encontraron 242 tursiones agrupados en 22 manadas. El tamaño promedio de grupo fue de 11 (DE = 8) delfines; sin embargo, es posible que las agrupaciones más comunes sean de menor tamaño. Los grupos con crías fueron significativamente más grandes (P < 0.05) y las manadas intercambiaron individuos constantemente. Los tursiones prefirieron la franja entre 250 y 500 m fuera de la costa (P < 0.05), con profundidades menores a 7 m y sustratos arenosos (P < 0.05). Se establecieron dos zonas con mayor frecuencia de avistamientos (P < 0.05) y el comportamiento alimentario fue más común cerca de la boca de la bahía. Se identificaron 169 animales mediante fotografías de las aletas dorsales y 124 de ellos fueron individuos diferentes. La costa al sur de San Quintín representó una zona de tránsito durante el periodo de estudio debido a que la mayoría de los tursiones (> 70%) fueron vistos en una sola ocasión o por corto tiempo. El catálogo de aletas dorsales desarrollado en este trabajo, en conjunto con el realizado en 1990 por Caldwell (1992), contiene a la fecha 220 animales; posiblemente estos delfines representan sólo una pequeña parte de la población. En la medida en que se desarrollen trabajos científicos sobre la biología poblacional de los tursiones en esta área geográfica y en otras adyacentes, se contribuirá a mejorar las estrategias de conservación y manejo de este recurso natural.
Palabras clave: distribución, movimientos, ecología, Tursiops truncatus, San Quintín.
Abstract
Twelve boat-based photoidentification surveys were carried out along the coast to the south of San Quintín Bay, in Baja California, Mexico, from July 1999 to June 2000; effort was 276.76 km and 31.7 h at sea. Twenty-two schools were encountered and 12.9 h of total observation were spent with 242 dolphins in these schools. The average school size was 11 (SD = 8) dolphins, although it is possible that groups are actually smaller; nursing groups were significantly larger (P < 0.05), with frequent membership exchanges occurring among schools. Dolphins preferred a coastal strip between 250 and 500 m offshore (P < 0.05), at depths below 7 m, and with sandy substrates (P < 0.05). Greater sighting frequencies occurred in two coastal zones (P < 0.05) and feeding was common around the mouth of the bay. Dorsal fin photographs led to the identification of 169 dolphins, and 124 were different individuals. During the study period, the coast south of San Quintín was a pass zone for transient dolphins, since most of the animals (>70%) were sighted one time or stayed for short periods. A total of 220 different dolphins have been identified in the San Quintín area when these data are combined with those gathered by Caldwell (1992) in 1990; these dolphins probably represent a small part of a larger population. More research on the population biology of the bottlenose dolphin in this and adjacent geographic areas is needed to develop better conservation and management strategies for this important natural resource.
Key words: distribution, movements, ecology, Tursiops truncatus, San Quintín.
Introducción
El tursión (Tursiops truncatus: Montagu, 1821) es considerado por muchos autores como uno de los cetáceos más conocidos y estudiados (Leatherwood y Reeves, 1990). El conocimiento popular de este delfín se debe principalmente a su distribución cosmopolita y hábitos costeros. Debido a ello, estos animales han sido aprovechados de formas muy diversas, como fuente de alimentación, por sus aceites y su piel (Gunter, 1942). También desde principios del siglo XX este animal ha sido extraído de su hábitat natural para ser entrenado y presentado en acuarios y delfinarios (Heckel, 1992). A pesar de esto, en muchos lugares aún se desconocen los aspectos más básicos de la ecología de estos animales; es por este motivo que la Unión Internacional para la Conservación de la Naturaleza (IUCN por sus siglas en inglés) considera el estado de riesgo de esta especie como desconocido. En México es necesario ampliar los límites geográficos y la duración de los estudios sobre tursiones para implementar programas adecuados de conservación, manejo y aprovechamiento (Morteo, 2002).
El presente trabajo es una continuación de los estudios sobre el tursión realizados por el Laboratorio de Comportamiento de Cetáceos de la Universidad Estatal de San Diego (SDSU) desde 1981 en la costa del Océano Pacífico en California, EUA, y Baja California, México. El objetivo de la presente investigación fue describir y analizar la distribución y los movimientos del tursión en las aguas adyacentes a San Quintín, Baja California, mediante técnicas de fotoidentificación.
Materiales y métodos
Navegaciones
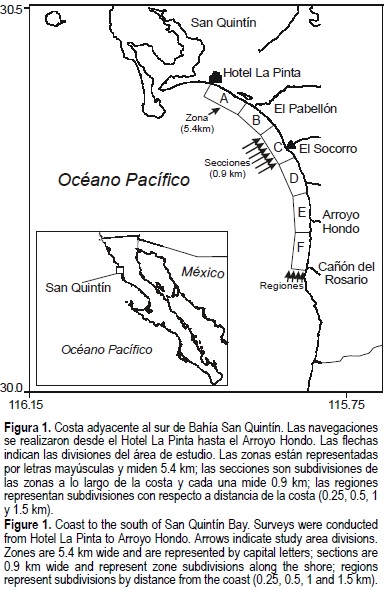
Entre julio de 1999 y junio de 2000 se realizaron navegaciones mensuales para fotografiar las aletas dorsales de los delfines y así identificarlos individualmente, según el método descrito por Defran et al. (1990). Se utilizó una panga de 5.5 ó 6.7 m (18 ó 22 pies) de eslora con un motor fuera de borda de 45 caballos de fuerza. Con el fin de realizar las navegaciones de manera sistemática, la costa se recorrió desde el Hotel La Pinta (30°23'24" N, 115°54'43.92' W; fig. 1) hacia el sur hasta el Arroyo Hondo, aproximadamente a 1.5 km de la costa (dependiendo del oleaje) y con una velocidad menor a 18 km h-1 (9.7 nudos). Las fechas de muestreo se agruparon por temporada de acuerdo con los máximos y mínimos de temperatura y el régimen de lluvia local (Hersh et al., 1990), por lo cual la primavera incluyó los meses de abril a junio, el verano de julio a septiembre, el otoño de octubre a diciembre y el invierno de enero a marzo. Todos los datos se recolectaron bajo condiciones del viento menores a Beaufort 3 (velocidad del viento <5.6 m s-1).
Tamaño y composición de los grupos
Para determinar el tamaño y la composición de los grupos se usó la definición de manada o grupo según Irvine et al. (1981). Las clases de edad (crías, jóvenes y adultos) fueron clasificadas según la literatura (Weller, 1991; Barco et al., 1999).
El tamaño de los grupos se revisó y corrigió por medio de los datos de fotoidentificación (Rossbach y Herzing, 1999). Se realizaron comparaciones por temporada del año mediante la prueba no paramétrica de Kruskal-Wallis (Neave y Worthington, 1988). Para determinar diferencias en el tamaño de grupo debido a la presencia de crías, se usó la prueba no paramétrica de Wilcoxon (Neave y Worthington, 1988). Los promedios reportados en este trabajo muestran entre paréntesis la desviación estándar (DE) seguida del tamaño de muestra.
Observaciones de comportamiento
Antes de obtener las fotografías, se registró el comportamiento de los delfines al aplicar el método ad libitum (Altman, 1974); para ello, un grupo se definió como el número máximo de delfines en asociación cercana, que exhibieran el mismo patrón de comportamiento. De acuerdo con Allen y Read (2000), el comportamiento fue clasificado como alimentación y no alimentación. Se utilizaron periodos de habituación y observación, ambos mayores a 10 min (Weller, 1998), y sólo se registró el comportamiento cuando al menos 20% del grupo se encontraba realizando la misma actividad (Hanson y Defran, 1993; Tepper, 1996).
Distribución espacial y uso del área
La localización geográfica de las manadas se registró por medio de un sistema de posicionamiento global (GPS). Para ello se consideró el punto más próximo al primer contacto con los delfines (Wilson et al., 1997).
El área de estudio se dividió en zonas de 5.4 km de extensión de acuerdo con Ballance (1992), desde el Hotel La Pinta hasta el Cañón del Rosario (fig. 1). Cada una de las seis zonas se subdividió posteriormente en seis secciones de 0.9 km a lo largo de la costa, y en cuatro regiones desde la costa hasta mar abierto (0.25, 0.5, 1.0 y 1.5 km) según Hanson y Defran (1993) (fig. 1). Para identificar el tipo de fondo presente, se determinó el tipo de sustrato con base en la literatura y las entrevistas a los pescadores locales, y se clasificó como arena, grava, roca, mantos de algas y desconocido (Hanson y Defran, 1993; Tepper, 1996; Morteo, 2002).
Como medida de esfuerzo se utilizó la distancia recorrida (kilómetros) durante la búsqueda de delfines (Reilly y Fiedler, 1994). Se realizó un balance de esfuerzo para reducir los sesgos ocasionados por sobremuestreos y navegaciones incompletas; se eliminaron los datos de una navegación cuando durante el mismo mes de muestreo se duplicó el esfuerzo sobre una misma zona, siempre y cuando la zona remuestreada fuera mayor a 1.5 km de longitud. También se eliminaron los datos cuando la zona no fue visitada al menos en el 85% de las navegaciones.
Para determinar la distribución espacial de los tursiones se graficaron las posiciones geográficas de las manadas sobre la zona de estudio (a priori). Las agrupaciones de avistamientos se compararon (a posteriori) mediante un análisis de escalamiento multidimensional (MDS por sus siglas en inglés, Kruskal y Wish, 1978). La distancia (kilómetros) entre avistamientos fue utilizada como índice de similitud para la prueba de MDS, y se usaron dos dimensiones según el modelo de Catell (Kruskal y Wish, 1978).
El uso del área fue establecido al comparar el número de animales presentes en cada división del área de estudio mediante pruebas de bondad de ajuste (Neave y Worthington, 1988). Para evitar la falacia estadística de combinación de datos (Machlis et al., 1985), se eliminaron los datos de las manadas que al moverse se ubicaron en una zona, región o tipo de sustrato diferente al inicial (Morteo, 2002).
Distribución temporal
Los movimientos de los delfines se dividieron en de mediano y de largo plazo. Estos datos se obtuvieron con base en la fotoidentificación; es decir, la mejor fotografía de cada animal se comparó con el resto de los individuos, y cuando la fotografía permitió identificar al mismo individuo en avistamientos diferentes se consideró una recaptura. Si no se encontraron empates se consideró un animal nuevo. De acuerdo con lo anterior, los movimientos a mediano plazo se establecieron al comparar las recapturas a lo largo de un año y los movimientos a largo plazo se determinaron al comparar las fotografías de este estudio contra las del catálogo realizado por Caldwell (1992) en la misma zona.
Abundancia relativa
Se calculó el número de delfines presentes con respecto a la distancia recorrida (kilómetros) y se multiplicó por un ancho de banda de 0.8 km de acuerdo con las condiciones de visibilidad y la velocidad del viento (< Beaufort 3) según Barco et al. (1999). Se comparó la abundancia relativa (delfines km-2) entre temporadas del año mediante la prueba de Kruskal-Wallis (Neave y Worthington, 1988).
Resultados
Se realizaron 12 navegaciones a lo largo de un año, en las que se recorrió un total de 276.8 km de costa en 31.7 h. Se observaron 242 delfines agrupados en 22 manadas durante un periodo de 12.9 h. Se tomaron 2297 fotografías y el 41.18% fueron útiles para identificar individualmente a los delfines.
Tamaño y composición de los grupos
El tamaño de grupo varió entre 2 y 36 delfines, con un promedio de 11 animales (DE = 8, n = 22 manadas). Los grupos más frecuentes tuvieron entre 4-6 y 13-15 delfines (fig. 2). Los grupos fueron más grandes durante el verano ( = 12.47, DE = 8.91, n = 15), seguido del otoño (= 10.5, DE = 6.36, n = 2), invierno (= 7.75, DE = 4.35, n = 4) y primavera (3, n = 1); sin embargo, no se encontró diferencia por temporada del año (Kruskal-Wallis, H = 6.11, P > 0.05, g.l. = 3, n = 22). Al combinar los datos de todo el año en relación con la presencia o ausencia de crías, se encontró que los grupos con cría (= 13.92, DE = 9.27, n = 12) resultaron ser más grandes en comparación con los grupos sin crías (= 7.50, DE = 4.42, n = 10) y la diferencia fue significativa (Wilcoxon, T = 86, P < 0.05, n = 22). De los 242 delfines avistados, el 8.5% fueron crías y casi un 1.5% fueron jóvenes.
= 12.47, DE = 8.91, n = 15), seguido del otoño (= 10.5, DE = 6.36, n = 2), invierno (= 7.75, DE = 4.35, n = 4) y primavera (3, n = 1); sin embargo, no se encontró diferencia por temporada del año (Kruskal-Wallis, H = 6.11, P > 0.05, g.l. = 3, n = 22). Al combinar los datos de todo el año en relación con la presencia o ausencia de crías, se encontró que los grupos con cría (= 13.92, DE = 9.27, n = 12) resultaron ser más grandes en comparación con los grupos sin crías (= 7.50, DE = 4.42, n = 10) y la diferencia fue significativa (Wilcoxon, T = 86, P < 0.05, n = 22). De los 242 delfines avistados, el 8.5% fueron crías y casi un 1.5% fueron jóvenes.

Distribución espacial, uso del área y comportamiento
Después de eliminar los recorridos en zonas remuestradas y poco visitadas, el tamaño de muestra se redujo de 22 a 16 avistamientos, sin que la distancia recorrida por zona fuera diferente (Kruskal-Wallis, H = 2.12, P > 0.05, g.l. = 4, n = 11; tabla 1).
La localización geográfica de estos 16 avistamientos se graficó con respecto a la división por zonas (fig. 3). Las zonas A y B presentaron sustrato arenoso; la zona C tuvo en su mayoría arena, con una pequeña playa de gravas a la altura de El Socorro; la zona D tuvo una transición entre mantos de algas fuera de la costa y gravas en la región de la playa; y la zona E consistió en su mayoría de sustrato rocoso, con gravas y mantos de alga en la frontera con la zona D. En el 40.9% de los casos se determinó que los delfines se alimentaron durante algún momento y se observó una mayor tendencia de alimentación en la parte norte del área de estudio, particularmente en la zona A (fig. 3). El 99% de estos avistamientos se encontraron en profundidades menores a 7 m. La comparación de esta distribución con el análisis MDS (estrés = 0.026, fig. 4) fue consistente en el 93.75% de los casos, es decir, se formaron los mismos grupos utilizando ambos métodos, con excepción del avistamiento número 11, el cual a priori se ubicó en la zona C y el MDS lo agrupó en la zona D.
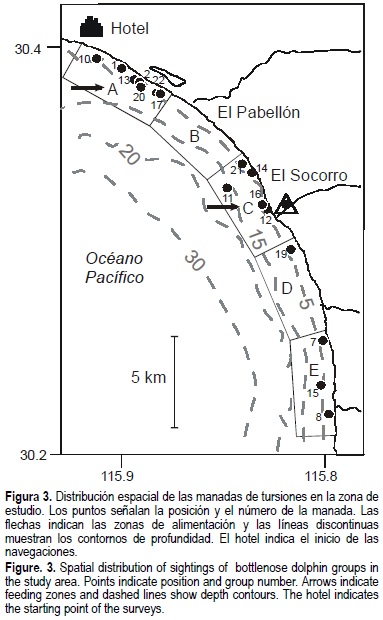

Con respecto al uso del área, después del balance de esfuerzo y de eliminar los avistamientos que incurrieron en la falacia estadística de combinación de datos, la zona A tuvo 99 delfines, la zona C tuvo 38, la zona D tuvo 6, la zona E tuvo 24 y no se observaron tursiones en la zona B. Se encontró una diferencia en el número de tursiones presentes por zona del área de estudio (χ2 = 188, P < 0.05, g.l. = 4, n = 167 delfines); de igual forma, hubo 137 delfines presentes en sustrato arenoso, 4 en grava, 14 en mantos de algas y ninguno fue observado en zonas con sustrato rocoso o desconocido, por lo que el sustrato arenoso tuvo una mayor cantidad de animales (χ2 = 457, P < 0.05, g.l. = 4, n = 155 delfines). Asimismo, la región más cercana a la costa (0-250 m) tuvo 11 animales, la región ente 250 y 500 m de la costa presentó 79 delfines, 14 fueron vistos en la región entre 500 y 1000 m y ninguno se observó de manera consistente más allá de los 1000 m; estas diferencias fueron estadísticamente significativas (χ2 = 148, P < 0.05, g.l. = 3, n = 104 delfines).
Distribución temporal
(a) Mediano plazo
A lo largo de un año, el número de animales presentes por temporada (n = 167 después del balance de esfuerzo) mostró un aumento significativo durante el verano (χ2 = 201, P < 0.05, g.l. = 3, n = 167 delfines; tabla 2).

Se identificaron 124 individuos diferentes de 169 aletas dorsales fotografiadas. El 72% de estos animales fueron fotografiados (capturados) una sola vez, 22% dos veces y el 9% fueron recapturados en tres ocasiones. Sin embargo, el 52% y el 70% de los delfines recapturados en dos y tres ocasiones, respectivamente, fueron avistados en días consecutivos a su primera recaptura. La acumulación de individuos identificados mostró un incremento constante, ya que en cada navegación un promedio del 75% de los animales avistados fueron individuos nuevos. También se encontró que de los 45 individuos recapturados, la mayoría (86%) se avistaron durante los meses de verano.
(b) Largo plazo
La comparación del catálogo fotográfico de este estudio con el de 1990 (Caldwell, 1992) resultó en 220 delfines diferentes. De lo anterior se obtuvo que sólo cuatro animales habían sido identificados en el área 10 años antes, la mayoría durante el verano. Cabe destacar que algunos de los animales identificados originalmente por Caldwell (1992) fueron identificados juntos en la misma manada, y reavistados días o años más tarde, en las mismas fechas, pero en manadas diferentes.
Abundancia relativa
La abundancia relativa de tursiones a lo largo del estudio promedió 1.00 delfines km-2 (DE = 0.9, n = 10 meses). Se observa una mayor abundancia relativa en los meses de verano (= 2.06 delfines km2, DE = 0.25, n = 3 meses; fig. 5).

Discusión
Las navegaciones realizadas durante el presente estudio (n = 12) son comparables con las realizadas por Caldwell (1992) 10 años antes (n = 8); sin embargo, el estudio de 1990 cubrió sólo la mitad del área de estudio y únicamente primavera y verano. No obstante, en ambos trabajos los resultados fueron similares en cuanto al número, tamaño, composición y movimientos de los grupos de tursiones.
Tamaño y composición de los grupos
El tamaño promedio de grupo en este estudio (> = 11, DE = 8, n = 22 manadas) resultó menor al reportado para las costas de San Diego (Weller, 1991; Tepper, 1996; Ward, 1998) y similar a lo encontrado en las costas de Ensenada y San Quintín (Espinosa, 1986; Sandoval, 1987; Caldwell, 1992; Guzón, 2002; tabla 3). Se ha observado que en sistemas semi-cerrados (bahías, lagunas y canales costeros), el tamaño de grupo generalmente es menor a seis delfines (Irvine et al., 1981; Scott et al., 1990; Weller, 1998); por el contrario, en sistemas abiertos (como la costa al sur de San Quintín) este valor generalmente supera los 15 animales (Würsig, 1978; Ross et al., 1989).

Würsig (1979) propuso que existe un equilibrio en el número de delfines necesarios para la defensa contra depredadores, la alimentación eficiente, las interacciones sociales y reproductivas, y la sobrevivencia de las crías. Se ha encontrado que la abundancia de peces es menor en el exterior de Bahía San Quintín y que los principales depredadores del tursión (tiburones) generalmente se encuentran alejados de la costa (Rosales-Casián, 1996). Esto podría explicar el que los grupos de tursiones sean de menor tamaño, ya que deben dispersarse para localizar a sus presas y no necesitan formar grupos grandes debido a la poca presencia de depredadores. Cabe destacar que la mayoría de los modelos depredador-presa no consideran las capacidades de ecolocalizacón de los delfines o la comunicación interespecífica como mecanismo de localización de presas y de prevención contra depredadores; por ejemplo, en regiones oceánicas donde el alimento es más escaso, se ha observado que los delfines se agregan en manadas de más de 100 individuos para localizar a sus presas al usar de manera conjunta su sistema de ecolocalización (Richardson et al., 1995).
En lo que respecta a la distribución bimodal del tamaño de grupo, Caldwell (1992) propuso que la formación de grupos pequeños en San Quintín se debe a la baja profundidad de la zona, lo que se refleja en la alta frecuencia de grupos de 4-6 animales. Por otra parte, la formación de grupos de 13-15 animales puede explicarse si se considera que generalmente los grupos se agregan en zonas de alimentación (Hanson y Defran, 1993) formando manadas de mayor tamaño (Rossbach y Herzing, 1999); esto fue observado particularmente en las zonas A y C. Cabe destacar que tanto el promedio del tamaño de grupo reportado por Caldwell (1992) como el de este trabajo se encuentran sesgados por valores extremos derivados de una distribución bimodal. Por lo tanto, el promedio del tamaño de grupo reportado hace 10 años ( = 14.28) y el encontrado en este trabajo ( = 11) no reflejan el tipo de agregación más común en la zona de estudio.
En diversos trabajos se ha planteado el posible papel social del tamaño y la composición de los grupos de delfines (Scott et al. , 1990); se ha propuesto que la presencia de crías puede determinar el número de animales agregados en una manada (Würsig, 1979; Scott et al., 1990; Weller, 1991, 1998). Se ha sugerido que este incremento presenta alguna ventaja social (aprendizaje y/o entrenamiento) o cierta tendencia hacia la protección de las crías, lo cual implica la existencia de micro-núcleos sociales dentro de las poblaciones (Irvine et al., 1981; Weller, 1991, 1998). Es factible que esto ocurra en el área de estudio, ya que se encontró un mayor número de animales en los grupos con crías; adicionalmente, a pesar del intercambio de individuos entre las manadas observadas, algunos de los delfines que cambiaron de grupo permanecieron relativamente cerca entre sí, debido a que fueron observados durante los mismos días.
Se ha reportado que en poblaciones cerradas la proporción de crías puede variar entre 2% y 6% (Leatherwood y Reeves, 1983); en poblaciones abiertas puede encontrarse hasta un 14% de crías (Irvine et al., 1981; Espinosa, 1986; Caldwell, 1992; Weller, 1998). Por lo tanto, con base en la proporción de crías, San Quintín se comporta como un sistema semicerrado. Cabe destacar que en ocasiones la asignación de los animales en una categoría de edad es diferente entre estudios; sin embargo, muchos autores reconocen al menos las tres categorías utilizadas en este trabajo (Leatherwood y Reeves, 1983; Caldwell, 1992; Weller, 1998).
Distribución espacial, uso del área y comportamiento
Es conocido que la distribución de presas del tursión generalmente es discontinua a lo largo de la costa (Defran et al., 1999); por este motivo la distribución de los tursiones tiende más a ser discreta que homogénea. En las costas del Pacífico los tursiones generalmente prefieren zonas estuarinas, ambientes rocosos y mantos de algas (Ballance, 1992; Hanson y Defran, 1993; Tepper, 1996; Ward, 1998). Se ha sugerido que los mantos de algas y las zonas rocosas presentan una mayor variedad de microhábitats, además de que proveen de refugio a los peces; esto explica que en San Quintín los delfines usan zonas arenosas para alimentarse, ya que posiblemente sus presas no pueden esconderse sobre este tipo de fondo (Würsig y Würsig, 1979; Tepper, 1996).
Se ha propuesto que las zonas con mayor número de avista-mientos (y de animales) pueden ser consideradas como zonas preferidas (Ross et al., 1989) y que este incremento puede relacionarse con la concentración de recursos alimentarios (Hui, 1985). En este trabajo las mayores frecuencias de avistamientos y comportamiento alimentario se observaron en las zonas A y C. No es inusual que esto suceda en dichas zonas debido a su cercanía con la boca de la Bahía San Quintín, la cual tiene una alta productividad (Álvarez-Borrego et al., 1975); por ejemplo, Estrada-Ramírez (2002) reportó la presencia de tursiones en el interior de Bahía San Quintín y Morteo et al. (2002) encontraron que algunos de ellos se alimentan en esta zona y permanecen en el lugar por más de 2 h.
Cabe destacar la ausencia de avistamientos en la zona B, aún cuando ésta fue una de las más recorridas. En la zona B se encuentra la localidad de El Pabellón y en ella se concentra la mayor cantidad de lanchas de la flota pesquera local; el tránsito de lanchas (principalmente pangas) es frecuente en esta zona aunque a baja escala (Morteo, 2002). Se ha documentado la conducta evasiva de los tursiones hacia embarcaciones de todo tipo; sin embargo, los tursiones generalmente se habitúan a su presencia después de cierto tiempo (Irvine et al., 1981; Ballance, 1992; Weller, 1998). La respuesta de éstos y otros cetáceos hacia las embarcaciones puede estar influenciada por una gran cantidad de variables, tales como la velocidad de la embarcación, el ángulo de acercamiento, el número de lanchas presentes, el espectro sónico emitido, la frecuencia audible de los animales, el tiempo de contacto, la profundidad de la zona, el tamaño y la composición de los grupos e incluso sus propias actividades (Richardson et al., 1995; Allen y Read, 2000; Heckel et al., 2001). Por lo tanto, la ausencia de avistamientos en la zona B podría implicar una tendencia a evitar el transitar en la zona B por periodos prolongados.
Distribución temporal a mediano y largo plazo
Se han realizado una gran cantidad de estudios sobre los movimientos de los tursiones a lo largo del año y algunos de ellos muestran poblaciones residentes, aunque una pequeña proporción de los individuos tienden a aparecer y desaparecer durante ciertas épocas (Würsig, 1978; Schramm, 1993). Se ha propuesto que la permanencia de los tursiones se relaciona principalmente con una estabilidad en la producción de recursos alimentarios a lo largo del tiempo (Wilson et al., 1997), lo cual podría resultar en áreas más propicias para actividades de alimentación y reproducción (Irvine et al., 1981) o para evitar a los depredadores (Scott et al., 1990).
Los tursiones exhibieron una baja fidelidad al sitio a mediano plazo, es decir, la mayor parte (72%) se fotografiaron una sola vez y los que regresaron al área lo hicieron durante periodos cortos (principalmente en verano). Por tal motivo, es posible que durante el periodo de estudio San Quintín representara un área de paso para los tursiones. Esta tendencia ha sido reportada en otros sistemas abiertos (Ballance, 1992; Caldwell, 1992; Weller, 1998) y es posible que esta característica favorezca el intercambio de individuos entre las manadas, reforzando la variabilidad genética de las poblaciones (Irvine et al., 1981; Scott et al., 1990).
El intervalo de tiempo entre las navegaciones realizadas para la presente investigación (aproximadamente 30 días) implica una baja continuidad en la escala temporal, por lo que la determinación del tiempo de residencia de los tursiones no es concluyente. Sin embargo, el estudio de Caldwell (1992) fue más puntual e intensivo y mostró la misma tendencia, por lo cual los movimientos de los delfines son consistentes a mediano y largo plazo.
Abundancia relativa
En diversos estudios se ha observado que la abundancia relativa comúnmente se encuentra entre 0 y 3 delfines km-2 (Ross et al., 1989; Heckel, 1992; Schramm, 1993; Barco et al., 1999). Debido a las variaciones estacionales de las características oceanográficas (relacionadas principalmente con las surgencias), es posible que en efecto exista un aumento en la cantidad de animales durante el verano.
La presencia casi permanente de tursiones en el área (Morteo, 2002) y la alta tasa de animales nuevos, sugieren que la población de animales que frecuentan las costas de San Quintín es mayor que sólo los 220 animales identificados por Caldwell (1992) y este estudio. Sin embargo, es necesario realizar estudios detallados para estimar de manera precisa el tamaño de la población de delfines en San Quintín, ya que debido al bajo número de recapturas no fue posible utilizar algún modelo para la estimación de la abundancia.
Agradecimientos
Agradecemos a la Secretaría del Medio Ambiente, Recursos Naturales y Pesca (ahora SEMARNAT) y al Instituto Nacional de Ecología (INE) las facilidades otorgadas a esta investigación (Permiso Pesca de Fomento No. 080699-231-03 y Of. No. DOO 02.-2481, respectivamente). Los mapas fueron elaborados con la ayuda de Tonatiuh Mendoza. Agradecemos también a los socios y voluntarios de ICMME, en especial a Gabriela Athié y Oscar Guzón, así como a los estudiantes del Laboratorio de Comportamiento de Cetáceos Aimeé Lang y Karine Viaud, su ayuda durante el trabajo de campo y laboratorio. También agradecemos a Horacio de la Cueva y a otros dos revisores anónimos sus comentarios hacia este trabajo.
Referencias
Allen, M.C. and Read, A. (2000). Habitat selection of foraging bottlenose dolphins in relation to boat density near Clearwater, Florida. Mar. Mamm. Sci., 16: 815-24. [ Links ]
Altman, J. (1974). Observational study of behavior sampling methods. Behaviour, 49: 227-67. [ Links ]
Álvarez-Borrego, S., Ballesteros, G. y Chee, A. (1975). Estudio de algunas variables fisicoquímicas superficiales en la Bahía de San Quintín en verano, otoño e invierno. Ciencias Marinas, 2: 1-9. [ Links ]
Ballance, L.T. (1992). Habitat use patterns and ranges of bottlenose dolphin in the Gulf of California, Mexico. Mar. Mamm. Sci., 8: 262-74. [ Links ]
Barco, S.G., Swingle, W.M., McLellan, W.A., Harris, R.N. and Pabst, D.A. (1999). Local abundance and distribution of bottlenose dolphins (Tursiops truncatus) in the nearshore waters of Virginia Beach, Virginia. Mar. Mamm. Sci., 15: 394-408. [ Links ]
Caldwell, M.J. (1992). A comparison of bottlenose dolphins identified in San Quintín and the Southern California Bight. M.Sc. thesis, San Diego State University, San Diego, CA, USA, 58 pp. [ Links ]
Defran, R.H., Shultz, G.M. and Weller, D.W. (1990). A technique for the photographic identification and cataloging of dorsal fins of the bottlenose dolphin (Tursiops truncatus). Rep. Int. Whal. Commn., 12: 53-55. [ Links ]
Defran, R.H., Weller, D.W., Nelly, D.L and Espinosa, M.A. (1999). Range characteristics of Pacific coast bottlenose dolphins (Tursiops truncatus) in the Southern California Bigth. Mar. Mamm. Sci., 15: 381-393. [ Links ]
Espinosa, M.A. (1986). Biología poblacional del delfín costero Tursiops truncatus en la costa noroccidental de Baja California, México. Tesis de licenciatura, Facultad de Ciencias Marinas, Universidad Autónoma de Baja California, Ensenada, México, 54 pp. [ Links ]
Estrada-Ramírez, A. (2002). Avistamientos de ballena gris Eschrichtius robustus en la Bahía de San Quintín, Baja California. Resúmenes del Congreso Bahía San Quintín en el 2002. Universidad Autónoma de Baja California, Instituto de Investigaciones Oceanológicas - Facultad de Ciencias Marinas, 17 19 de abril, Ensenada, Baja California, México, p. 7. [ Links ]
Gunter, G. (1942). Contributions to the natural history of bottlenose dolphin, Tursiops truncatus (Montague), on the Texas coast, with particular reference to food habits. J. Mamm., 23: 267-76. [ Links ]
Guzón, O. (2002). Distribución y movimientos del tursión (Tursiops truncatus) en la Bahía de Todos Santos, Baja California, México (Cetacea: Delphinidae). Tesis de licenciatura, Facultad de Ciencias Marinas, Universidad Autónoma de Baja California, Ensenada, Baja California, México, 156 pp. [ Links ]
Hanson, M.T. and Defran, R.H. (1993). The behaviour and feeding ecology of the Pacific coast bottlenose dolphin, Tursiops truncatus. Aqua. Mamm., 19: 127-142. [ Links ]
Heckel, G. (1992). Fotoidentificación de tursiones Tursiops truncatus (Montagu, 1821) en la Boca de Corazones de la Laguna de Tamiahua, Veracruz, México (Cetacea: Delphinidae). Tesis de licenciatura, Facultad de Ciencias, Universidad Nacional Autónoma de México, México, D.F., 164 pp. [ Links ]
Heckel, G., Reilly, S.B., Sumich, J.L and Espejel, I. (2001). The influence of whalewatching on the behavior of migrating gray whales (Eschrichtius robustus) in Todos Santos Bay and surrounding waters, Baja California, Mexico. J. Cetacean Res. Manage., 3: 227-237. [ Links ]
Hersh, S.L., Odell, D.K. and Asper, E.D. (1990). Bottlenose dolphin mortality patterns in the Indian/Banana River system of Florida. In: S. Leatherwood and R. Reeves (eds.), The Bottlenose Dolphin. Academic Press, San Diego, California, pp. 155-164. [ Links ]
Hui, C.A. (1985). Undersea topography and the comparative distribution of two pelagic dolphins. Fish. Bull., 83: 472-75. [ Links ]
Irvine, A.B., Scott, M.D., Wells, R.S. and Kaufmann, J.H. (1981). Movements and activities of the Atlantic bottlenose dolphin, Tursiops truncatus, near Sarasota, Florida. Fish. Bull., 79: 671-688. [ Links ]
Kruskal, J.B. and Wish, M. (1978). Multidimensional scaling. Sage University Paper Series on Quantitative Applications in the Social Sciences. Sage Publ., Beverly Hills, California, and London, pp. 7-11. [ Links ]
Leatherwood, S. and Reeves, R.R. (1983). Abundance of bottlenose dolphins in Corpus Christi Bay and coastal southern Texas. Contrib. Mar. Sci., 26: 179-99. [ Links ]
Leatherwood, S. and Reeves, R.R. (1990). The Bottlenose Dolphin. Accademic Press, San Diego, California, 653 pp. [ Links ]
Machlis, L., Dood, P.W.D. and Fentress, J.C. (1985). The pooling fallacy: Problems arising when individuals contribute more than one observation to the data set. Z. Tierpsychol., 68: 201-14. [ Links ]
Morteo, E. (2002). Distribución y movimientos del tursión (Tursiops truncatus; Montagu, 1821) en las aguas adyacentes a San Quintín, Baja California, México (Cetacea: Dephinidae). Tesis de licenciatura, Facultad de Ciencias Marinas, Universidad Autónoma de Baja California, Ensenada, Baja California, México, 146 pp. [ Links ]
Morteo, E., Olivera, L.D. y de la Cueva, H. (2002). Estudio prospectivo sobre el estado de Tursiops truncatus en la Bahía de San Quintín. Centro de Investigación Científica y de Educación Superior de Ensenada, Baja California, México, 23 pp. [ Links ]
Neave, H.R. and Worthington, P.L. (1988). Distribution-free Tests. Unwin Hyman, London, 430 pp. [ Links ]
Reilly, S.B. and Fiedler, P.C. (1994). Interannual variability of dolphin habitats in the eastern tropical Pacific. I: Research vessel surveys, 1986-1990. Fish. Bull., 92: 434-50. [ Links ]
Richardson, W.J., Greene, C.G., Malme, C.I. and Thomson, D.H. (1995). Marine Mammals and Noise. Academic Press, San Diego, California, 575 pp. [ Links ]
Rosales-Casián, J.A. (1996). Ictiofauna de la Bahía de San Quintín, Baja California, México y su costa adyacente. Ciencias Marinas, 22: 443-58. [ Links ]
Ross, G.J., Cockroft, V.G., Melton, D.A. and Butterworths, D.S. (1989). Population estimates for bottlenose dolphins Tursiops truncatus in Natal and Transkei waters. Afr. J. Mar. Sci., 8: 119-29. [ Links ]
Rossbach, K.A. and Herzing, D.L. (1999). Inshore and offshore bottlenose dolphin (Tursiops truncatus) communities distinguished by association patterns near Grand Bahama Island, Bahamas. Can. J. Zool., 77: 581-92. [ Links ]
Sandoval, A. (1987). Movimientos y comportamiento del delfín nariz de botella (Tursiops truncatus) en la Bahía de Todos Santos, BC, México. Tesis de licenciatura, Facultad de Ciencias Marinas, Universidad Autónoma de Baja California, Ensenada, México, 100 pp. [ Links ]
Schramm, Y. (1993). Distribución, movimientos, abundancia e identificación del delfín Tursiops truncatus (Montagu, 1821), en el sur de la Laguna de Tamiahua, Ver. y aguas adyacentes (Cetacea: Delphinidae). Tesis de licenciatura, Escuela de Biología, Universidad Autónoma de Guadalajara, Guadalajara, Jalisco, México, 174 pp. [ Links ]
Scott, M.D., Wells, R.S., Irvine, A.B. and Mate, B. (1990). Tagging and marking studies on small cetaceans. In: S. Leatherwood and R. Reeves (eds.), The Bottlenose Dolphin, Academic Press, San Diego, California, pp. 489-513. [ Links ]
Tepper, E.M. (1996). Feeding duration in the Pacific coast bottlenose dolphin (Tursiops truncatus). M.Sc. thesis, San Diego State University, California, USA, 81 pp. [ Links ]
Ward, B.G. (1998). Movement patterns and feeding ecology of the Pacific coast bottlenose dolphin (Tursiops truncatus). M.Sc. thesis, San Diego State University, California, USA, 57 pp. [ Links ]
Weller, D.W. (1991). The social ecology of Pacific coast bottlenose dolphins. M.Sc. thesis, San Diego State University, California, USA, 78 pp. [ Links ]
Weller, D.W. (1998). Global and regional variation in the biology and behavior of bottlenose dolphins. Ph.D. thesis, University of Texas A & M, USA, 142 pp. [ Links ]
Wilson, B., Thompson P.M. and Hammond P.S. (1997). Habitat use by bottlenose dolphins: Seasonal distribution and stratified movement patterns in the Moray Firth, Scotland. J. Appl. Ecol., 34: 1365-74. [ Links ]
Würsig, B. (1978). Occurrence and group organization of Atlantic bottlenose porpoises (Tursiops truncatus) in Argentina Bay. Biol. Bull., 154: 348-359. [ Links ]
Würsig, B. (1979). Dolphins. Sci. Am., 240: 136-48. [ Links ]
Würsig, B. and Würsig, M. (1979). Behavior and ecology of the bottlenose dolphin, Tursiops truncatus, in the south Atlantic. Fish. Bull., 77: 399-412. [ Links ]

