










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkCiencias marinas
versão impressa ISSN 0185-3880
Cienc. mar vol.30 no.1a Ensenada Fev. 2004
Artículos
Nutrient and phytoplankton dynamics in a coastal lagoon strongly affected by coastal upwelling
Dinámica de nutrientes y fitoplancton en una laguna costera fuertemente afectada por surgencias costeras
Saúl Álvarez-Borrego
Departamento de Ecología, Centro de Investigación Científica y de Educación Superior de Ensenada (CICESE). Carretera Tijuana-Ensenada Km 103 Ensenada, CP 22860, Baja California, México. E-mail: alvarezb@cicese.mx
Recibido en septiembre de 2002;
Aceptado en agosto de 2003.
Abstract
San Quintín Bay is a coastal lagoon influenced by the California Current System (CCS) coastal upwelling. Upwelling brings nutrient-rich waters near the bay mouth and tidal currents propagate those waters throughout the bay. Upwelling intensification and relaxation events occur with a period of ~2 weeks, possibly due to the variability of circulation of the CCS. Off San Quintín, the CCS has an offshore component of the flux, causing upwelling intensification events as strong as off Point Conception (34.5°N), with high phytoplankton productivity. At the lagoon's mouth, upwelling is the main cause of variability for all physico-chemical properties except temperature. Semi-diurnal tides are the main cause of variability for temperature. Nutrient remineralization at the sediments and turbulence induced by tidal currents and wind waves increase nutrient concentrations in the lagoon. At the heads of the bay, phytoplankton abundance was ten-fold lower, productivity and chlorophyll concentrations were three-fold lower, chlorophyll content per cell was three-fold higher, and turbidity was higher than at the mouth. The few available data suggest the hypothesis that the effect of ENSO events on phytoplankton biomass depends on the interdecadal regime shifts in the northeastern Pacific. At the lagoon's mouth and adjacent ocean, summer salinities as low as 32.4 suggest the arrival of water parcels that originate in the north, possibly the Columbia River estuarine plume.
Key words: coastal lagoon, phytoplankton, nutrients, coastal upwelling, interannual variation.
Resumen
Bahia San Quintín es una laguna costera influenciada por surgencias costeras del Sistema de la Corriente de California (SCC). Las surgencias acarrean aguas ricas en nutrientes cerca de la boca de la bahía y las corrientes de marea las propagan a través de toda la bahía. Existen eventos de intensificación y relajamiento de surgencias con un periodo de ~2 semanas, posiblemente debidos a la variabilidad de la circulación del SCC. El SCC tiene un componente de flujo hacia mar adentro frente a San Quintín. Esto causa eventos de intensificación de surgencia tan fuertes como los que se dan frente a Point Conception (34.5°N), con alta productividad fitoplanctónica. En la boca de la laguna las surgencias son la causa principal de variabilidad de todas las propiedades físicas y químicas, con excepción de la temperatura que es afectada principalmente por las mareas semidiurnas. La remineralización de los nutrientes en los sedimentos, y la turbulencia inducida por las corrientes de marea y el oleaje incrementan las concentraciones de nutrientes en la laguna. En los extremos internos de la bahía hay hasta diez veces menos abundancia de fitoplancton, tres veces menos productividad y concentración de clorofila, tres veces más alto contenido de clorofila por célula, y mayor turbidez que en la boca de la bahía. Los pocos datos disponibles sugieren la hipótesis de que el efecto de los eventos de El Niño en la biomasa del fitoplancton depende de los cambios interdecadal es de régimen climático en el Pacífico nororiental. En la boca de la laguna y el océano adyacente, salinidades de verano tan bajas como 32.4 sugieren el arribo de parcelas de agua originadas en el norte, posiblemente de la lengüeta estuarina del Río Columbia.
Palabras clave: laguna costera, fitoplancton, nutrientes, surgencia costera, variación interanual.
Introduction
San Quintín Bay is a coastal lagoon where salinity increases from the mouth to the inner reaches due to the high evaporation rate and almost nil freshwater input from rainfall and surface runoff (Chávez-de-Nishikawa and Alvarez-Borrego, 1974; Alvarez-Borrego et al., 1975). According to Kjerfve's (1994) classification, it is a restricted lagoon, with a permanent single connection to the ocean and tides that co-oscillate with tides in the coastal ocean with little reduction of amplitude inside the lagoon. It is influenced by coastal upwelling of the California Current System (CCS), mainly in late spring and summer. The uniqueness of the oceanic area adjacent to San Quintín Bay is that the effect of the oceanic circulation is added to that of the winds to produce very intense upwelling. Off San Quintín, circulation not only has a component parallel to the coast and towards the equator, but it also has a strong offshore component to compensate partially for the onshore flux off Ensenada (fig. 1).
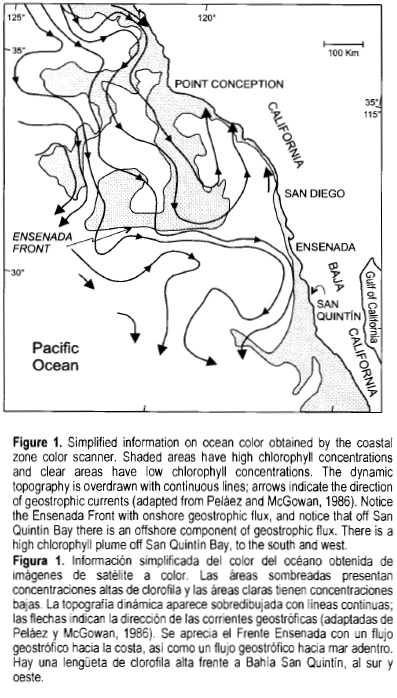
Using ocean color satellite imagery, Peláez and McGowan (1986) defined a sharp zonal boundary, the Ensenada Front, about 160-500 km offshore. It is a persistent characteristic of the region, with colder and higher chlorophyll water towards the north of the front. It clearly coincides with an intense onshore flux of the CCS. When meeting the coast, this flux divides into two: a northward flux, which is the oligotrophic intrusion of the Southern California Bight, and a southward flux; the latter flux turns offshore off San Quintín in a similar manner as the offshore flux off Point Conception. These offshore fluxes produce similarly intense coastal upwelling, as is described later. Upwelling affects even the internal extremes of San Quintín Bay because of the tidal currents that renew its waters (Lara-Lara et ah, 1980; Alvarez-Borrego and Alvarez-Borrego, 1982). Upwelling occurs in a sequence of intensification and relaxation events with a ~2-week period, the most intense events occurring in July. During an upwelling intensification event the lagoon has greater water exchange with the open ocean and this can be detected even in the inner arms (Alvarez-Borrego and Alvarez-Borrego, 1982).
The first studies of San Quintín Bay were carried out in 1960/1961, mainly on its sedimentology and benthic biota (Barnard, 1962; Dawson, 1962; Gorsline and Stewart, 1962). Researchers from the University of Baja California began working at the lagoon in 1973, and their main motivation was the development of aquaculture, which lead to nutrient and phytoplankton studies (Chávez-de-Nishikawa and Alvarez-Borrego, 1974; Álvarez-Borrego and López-Álvarez, 1975; Lara-Lara and Alvarez-Borrego, 1975). Macronutrients are not limiting to phytoplankton photosynthesis in San Quintín Bay (Álvarez-Borrego and Chee-Barragán, 1976; Silva-Cota and Álvarez-Borrego, 1988). Besides upwelling and tidal currents, other natural fertilization mechanisms occur in these coastal lagoons, such as nutrient remineralization in the water column, and in the sediments due to bacteria, with input to the water column by turbulence (Álvarez-Borrego and Chee-Barragán, 1976; Camacho-Ibar and Álvarez-Borrego, 1988).
The objective of this contribution is to review existing data to describe nutrient and phytoplankton dynamics in San Quintín Bay over a wider time scale spectrum than those of the individual contributions, from hours to decades, and within the spatial scale of events in the CCS. Phenomena that were not well-known earlier, such as interannual variations due to El Niño and La Niña events (Torres-Moye and Álvarez-Borrego, 1987; Silva-Cota and Álvarez-Borrego, 1988) and inter-decadal variations produced by "cold and warm regimes" in the North Pacific (Chávez et al, 2003), are taken into account.
Physical description
San Quintín Bay is a K-shaped lagoon of approximately 42 km2. The western arm is named Falsa Bay and the eastern arm is named San Quintín Bay (fig. 2). The organic carbon and nitrogen contents in sediments increase broadly towards the bay heads, paralleling the increase in clay content. The lagoon is quite shallow, about 85% of the eastern arm lying in depths of 1.8 m or less at mean high water. Depths greater than this occur in channels that are strongly differentiated from the shallow bay flats by sharp depth changes. In general, the channels have depths of 5-7 m, with one record of -10 m. Except during the few years of heavy rainfall, there are no flowing streams with surface freshwater input to the bay (Barnard, 1962). The mean annual precipitation is 15 cm and the mean annual evaporation is 140 cm; rainfall is restricted to the period from November to March (Camacho-Ibar et al, 2003). As evidenced by saline intrusion, over-extraction of groundwater for agriculture in recent years has induced a reversal of the normal groundwater flow, making coastal aquifers unlikely sources of nutrients to the lagoon (Aguirre-Muñoz et al, 2001). Domestic waste is not a significant source of nutrients to the lagoon (Camacho-Ibar et al, 2003).
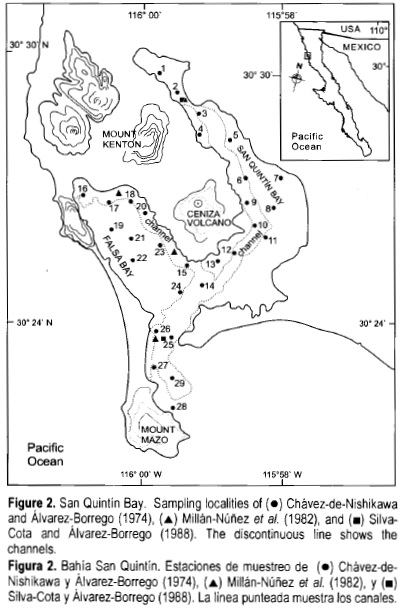
Tides are the main cause of water circulation and explain up to 97% of measured currents. Bottom friction and wind forcing affect the residual circulation, causing transverse currents and eddies that affect water exchange with the open ocean. Tides are predominantly semidiurnal, with ranges of 2.5 and 1.0 m for spring and neap tides, respectively, throughout the lagoon (Martori-Oxamendi, 1989). Tidal currents of up to 100 cm s_1 occur in the region near the mouth, decreasing toward the inner reaches to maximum velocities of -60 cm s-1 due to volume continuity (Ocampo-Torres, 1980). Vertical distribution of water properties is homogeneous because of turbulence caused by tidal currents (Martori-Oxamendi, 1989).
Spatial and seasonal variability
Horizontal gradients and seasonal variations of the lagoon's water properties were first defined from monthly samplings in 1973 and 1974 (Chávez-de-Nishikawa and Alvarez-Borrego, 1974; Alvarez-Borrego et al, 1975; Alvarez-Borrego and López-Alvarez, 1975; Lara-Lara and Alvarez-Borrego, 1975; Álvarez-Borrego and Chee-Barragán, 1976). Results based on single instantaneous samplings carried out during 1973/1974 revealed that the lagoon water properties had patchy distributions due, among other things, to the irregular bathymetry, and that there was a great local temporal variation due to tidal currents. Later experiments in which simultaneous time series were generated by sampling every 2 h during several days, confirmed that some properties have clear horizontal gradients. For example, chlorophyll and phytoplankton abundance were in general higher for the mouth than for the inner reaches of the lagoon during the summer of 1979, a non-El Niño year (Millán-Núñez et al, 1982). With relation to the 1973/1974 sampling, only January and July data were used to show the main seasonal and spatial variations, from three regions: the lagoon's mouth and the two internal extremes, Falsa Bay and San Quintín Bay.
In summer, water temperature increased from the mouth to the inner reaches, with highest temperatures in the eastern arm. During the winter of 1973, this gradient reversed, with temperature decreasing from the mouth to the internal extremes because of heat exchange with a cold atmosphere (fig. 3a). Salinity showed a similar pattern in summer and winter of 1973, increasing from the mouth to the inner reaches, with highest values in the eastern arm (up to >35) (fig. 3b). This is an indication of a longer water residence time in San Quintín Bay than in Falsa Bay, both in summer and winter. The greater horizontal salinity gradient for July than for January was due to a higher evaporation rate during summer. In the eastern arm, salinities >37 have been recorded for June, July, August and November (Álvarez-Borrego et al, 1975; Millán-Núñez et al, 1982; Camacho-Ibar et al, 2003), possibly due to sampling during upwelling relaxation periods and neap tides that produce high water residence time in the inner reaches of the lagoon.
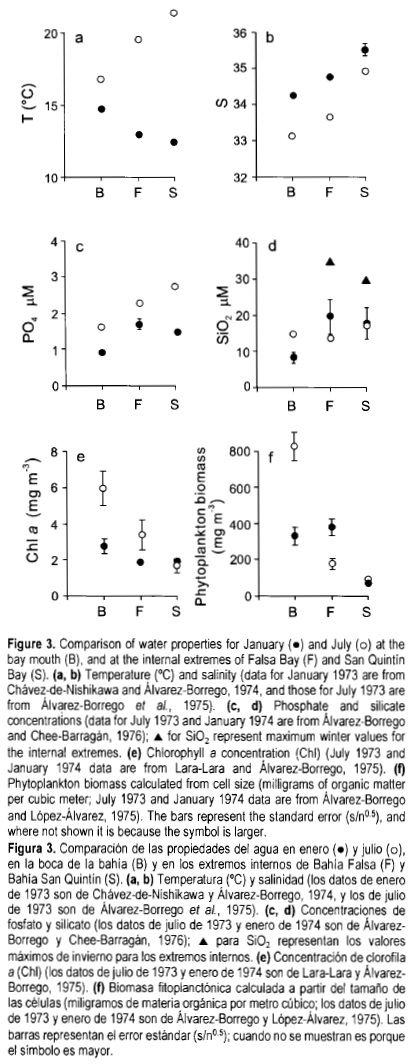
Summer salinity values were lower than those for winter throughout the lagoon in 1973/1974 (fig. 3b), because of the seasonal variation in the CCS circulation and the possible effect of the Columbia River estuarine plume, as will be discussed later. The arrival of low salinity water parcels from the north is perceived through low summer salinity values off San Quintín Bay and at its mouth, with flood flow. Typical salinity values for the CCS surface waters off northwestern Baja California are 33.4-33.6 throughout the year (e.g., CalCOFI Data Report, 1984), but eddies may carry parcels of water with lower salinity. At the mouth of San Quintín Bay, salinity was 33.00 in July 1973 during high tide and, according to the temperature values, within an upwelling relaxation period. The origin of high oceanic winter salinities is to the south of this area. Surface salinities of the CCS increase towards the south to >34 off the tip of the Baja California peninsula, and during winter, inshore surface water flows northwards (Reid, 1960; Hickey, 1979). However, salinity values >34 may be found any time of the year at the lagoon's mouth and adjacent oceanic area with ebb flow, and during spring and summer with upwelling intensification events even with flood flow (e.g., data in Lara-Lara et al, 1980; Camacho-Ibar et al, 2003).
In general, reactive phosphate (PO4) and dissolved silicon (H4SiO4, symbolized here as SiO2) concentrations increased from the lagoon's mouth to the internal extremes in 1973/1974 (Alvarez-Borrego and Chee-Barragán, 1976) (fig. 3c, d). In summer, PO4 values clearly increased inward, with highest values in the eastern arm, whereas in winter the gradient was weaker and there was no significant difference between Falsa Bay and San Quintín Bay (fig. 3c). A survey carried on in 1995/1996 by Camacho-Ibar et al. (2003) confirms that this general tendency for the PO4 spatial change remains essentially the same. Summer SiO2 values were similar throughout the bay, about 15 µM, while in winter the SiO2 values were both low and very high (up to >30 µM) in the inner reaches, but the means for the three regions were not significantly different (fig. 3d).
Barnard (1962) hypothesized an input of underground freshwater to explain the high SiO2 values from the eastern arm. However, high SiO2 values in Falsa Bay would have to be explained in terms of water exchange between the two arms of the bay, which would be complicated. Besides, as Aguirre-Muñoz et al (2001) indicated, over-extraction of groundwater for agriculture in recent years has induced a reversal of the normal groundwater flow. New winter SiO2 data are needed to test Barnard's (1962) hypothesis. Maximum SiO2 values for the eastern arm for August 1995 and 1996 were ~20 µM, associated with salinities >35 (Ibarra-Obando et al, 2001), and they are similar to the 1973 data. Without the influence of the aquifer, no high SiO2 values (>30 µM) should be detected for the eastern arm if Barnard (1962) is correct. In January, the effect of coastal upwelling, if any, is very low. An alternative explanation for the high SiO2 values at the internal extremes (fig. 3d) is the dissolution of exoskeletons of organisms such as diatoms and silicoflagellates deposited in the sediments. This dissolution is much slower than organic matter degradation (Libes, 1992). The intense production of diatoms during spring and summer upwelling events, their introduction to the lagoon's interiors by tidal currents, and their sedimentation was not immediately followed by clear large increments of SiO2 values in the water column.
Throughout the lagoon, higher PO4 values for July 1973 than for January 1974 (fig. 3c) were due to summer upwelling in the adjacent ocean, although as mentioned above, the July sampling was done within a relaxation period. Possibly, there was a larger import of organic matter into the lagoon during summer than during winter. Open Pacific Ocean deep waters have maximum PO4 concentrations of 3.3 µM (Libes, 1992), but the highest PO4 values reported for the eastern arm of San Quintín Bay are > 4 µM for November (Álvarez-Borrego and Chee-Barragán, 1976) and August (Camacho-Ibar et al, 2003). The origin of these high values cannot be the adjacent ocean. Álvarez-Borrego and Chee-Barragán (1976) suggested that the high PO4 and SiO2 values for the internal extremes are a result of the organic matter enrichment of the sediments and its degradation. Tidal currents transport particulate organic matter, mainly phytoplankton, from the adjacent oceanic area into the lagoon's arms, which is deposited on the surface sediments where it is decomposed; however, both pelagic and benthic nutrient regeneration may occur (Knoppers, 1994). The contribution to regeneration by the dissolved organic fraction has been noted to be of significant importance in shallow water sediments (Teague et al, 1988). Álvarez-Borrego and Chee-Barragán (1976) indicated that high PO4 values in the internal extremes of San Quintín Bay suggest that more organic matter is being consumed than photosynthesized in the lagoon. Smith and Hollibaugh (1997) reported a similar situation for Tómales Bay in California. Lara-Lara et al (1980) reported large PO4 spikes at the mouth of San Quintín Bay caused by intensified winds stirring the bay-bottom sediments into the water column. This is an indication that the sediment released during wind mixing events is a source of inorganic phosphate for the whole lagoon. Besides the direct effect of winds on mixing, a most effective mechanism is wind waves. Using a steady-state three-box model and data from three 2-day samplings within the main channels and with neap tides (August 1995, February 1996 and August 1996), Camacho-Ibar et al. (2003) concluded that the lagoon has a net heterotrophic condition and that it is a net exporter of inorganic phosphate, but they mentioned that a high frequency (<2-week) time course study of nutrients is required to evaluate the potential short-term variations in nonconservative nutrient fluxes of this lagoon, affected by the biweekly upwelling cycles during spring and summer.
The lagoon is a net exporter of ammonia (Farfán and Álvarez-Borrego, 1983) and phosphate (Camacho-Ibar et al, 2003) due to the intense remineralization in the system, but it imports nitrite, nitrate, and particulate organic carbon and nitrogen (mostly in the form of diatoms) during upwelling events (Farfán and Álvarez-Borrego, 1983). Nitrogen fixation minus denitrification indicates that San Quintín Bay is a net sink of nitrogen (Camacho-Ibar et al, 2003), but this was the result for only 2-day sampling periods. Usui et al. (2001) measured nitrification and denitrification processes in a Japanese estuary using 15N as a tracer, and reported that the rate of these processes changed with time, sometimes one exceeding the other and vice versa. Ammonia concentration in San Quintín Bay changes irregularly both temporally and spatially, with a range of < 1 to ~5 µM, and exceptionally up to ~8 µM, and with no clear difference between winter and summer values (Farfán and Álvarez-Borrego, 1983; Osorno-Velázquez, 2000). Nitrite concentration ranged from 0 to 0.42 µM, with a mean of 0.18 µM in June and July 1979, at the mouth (Farfán and Álvarez-Borrego, 1983).
Chlorophyll values were higher at the mouth than in the internal extremes in 1973/1974, and at the mouth they were higher in July than in January, due to the effect of summer upwelling (Lara-Lara and Álvarez-Borrego, 1975) (fig. 3e). In July 1973, high chlorophyll values (up to >8 mg m-3) in the mouth region indicate that upwelling was in a relaxation period, as also indicated by the relatively high temperature and low salinity values (fig. 3a, b). Phytoplankton populations had already grown when sampling was done. The spatial chlorophyll range was larger for July than for January, but at the internal extremes the chlorophyll means were not significantly different between summer and winter, or between the two extremes for the same season. Mean primary productivity values, estimated with the light-chlorophyll method, were also highest for the mouth and for July (1.7 gC n-3 d-1), with values of -0.5 gC n-3 d-1 for all other cases (not shown) (Lara-Lara and Álvarez-Borrego, 1975); however, these latter authors did not take into consideration the effect of the lagoon's water turbidity on light attenuation and their values might have been overestimated. Primary productivity values generated with the 14C method (Lara-Lara et al, 1980; Millán-Núñez et al, 1982) were lower by up to a factor of two. Barnard (1962) reported Secchi disk readings for 1960/1961 of 2-2.7 m for the mouth, 1.2-1.5 m for San Quintín Bay, and ~1 m for Falsa Bay. Álvarez-Borrego et al. (1977b) reported Secchi disk readings of 2.2-1.0 m for the mouth, for spring and summer of 1975. Osorno-Velázquez (2000) reported 3-3.5 m for the eastern arm and 1-3 m for Falsa Bay, for September 1999 and January 2000; this author explained that the low Secchi readings for Falsa Bay were due to the effect of increased turbidity because of oyster cultures in that arm. However, some of Barnard's (1962) values for this arm, obtained long before any aquacul-ture project, were as low as those of Osomo-Velázquez (2000). High turbidity in Falsa Bay is caused by the effect of wind waves on the shallow bathymetry. The volcanoes that separate the two arms of the lagoon protect the eastern arm and allow for much smaller waves in San Quintín Bay. These Secchi disk readings are very low and imply strong light attenuation, in some cases up to 50% in the first 0.5 m depth.
On 6 July 1977, phytoplankton abundance decreased from the mouth to the eastern extreme (not shown). Diatoms had the strongest decrease, from >400 cells mL-1 at the mouth to <20 cells mL-1 at the eastern extreme. Diatom abundance at Falsa Bay was intermediate, between the other two. Dinoflagellate abundance was similar for the three places (>80 cells mL-1), although slightly higher for the mouth, and microflagellate abundance was highest for the mouth (~110 cells mL-1), decreasing to similarly low values for the two internal extremes (<20 cells mL-1) (Álvarez-Borrego and Nájera-de-Muñoz, 1979). These values were for samples taken with flood flow and the high diatom abundances were due to an upwelling intensification event during sampling. Minimum surface temperature at the mouth on 6 July 1977 was 12°C, indicative of intense upwelling (Lara-Lara et al, 1980). These phytoplankton abundances do not take into consideration cells <3 µM because the inverted microscope technique did not detect them. Phytoplankton biomass, in milligrams of organic matter per cubic meter, was also clearly higher at the mouth than in the internal extremes in July 1973, with a minimum value in the eastern arm. Values for January 1974 were lower, with similar values for the mouth and Falsa Bay and a minimum value for San Quintín Bay (fig. 3f).
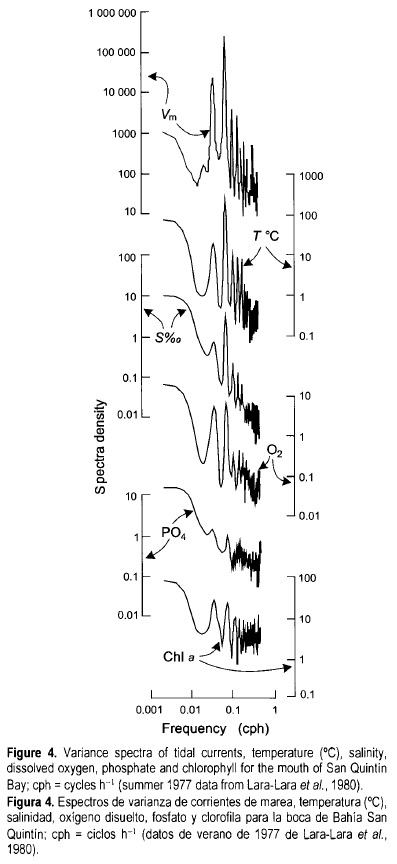
In 1975, time series were generated at the mouth to characterize the short-period variability, sampling every hour during 24 h. Results showed that properties such as chlorophyll change as much as one order of magnitude in only 2 h, but the causes of this variability were not clear (Álvarez-Borrego et al, 1977b). Longer time series of water properties were generated in June and July 1977, at the mouth, sampling every hour during 18 days (Lara-Lara et al, 1980). These data allowed for time series analysis to characterize the main sources of variation. Alternation of upwelling intensification and relaxation events were characterized as a factor causing a 2-week period variation of physical, chemical and biological water properties. Alternation of upwelling events was the main cause of variability for all properties except temperature. Semidiurnal tides were the main cause of variability for temperature. Conservative variables such as temperature and salinity had a semidiurnal component greater than the diurnal one. Most non-conservative variables (e.g., chlorophyll and PO4) had equal diurnal and semidiurnal variability components, due to biological processes that strongly depend on the solar radiation cycle (fig. 4). This type of variation was confirmed with the results from the June/July 1979 sampling (Millán-Núñez et al, 1982), and with data from continuous recording thermographs operated during a whole year in 1979 and 1980 (Álvarez-Borrego and Álvarez-Borrego, 1982). The 1979 time series was 10 days long, sampling every 2 h, and it covered four sampling points simultaneously: the mouth, the Y vertex and the two internal extremes.
The effect of the sequence of upwelling intensification and relaxation events on seawater properties at the lagoon's mouth was very clear in the June/July 1979 data (fig. 5). With intense upwelling, temperature was as low as ~11°C (fig. 5a), nitrate concentration (NO3) was >12 µM, and diatoms dominated the phytoplankton community (fig. 5b, d). With upwelling relaxation, the water warms up due to an increase in residence time off the bay mouth, presenting ideal conditions for phytoplankton growth; nutrients are used more efficiently and are not being input as fast from deeper waters to the surface, and there is weaker vertical advection and turbulence. After about four days, nutrients such as NO3 decreased to <1.0 µM, and phytoplankton composition changed from diatoms to dinoflagellates (fig. 5b, d). Chlorophyll concentration had a general tendency to increase from the period of intense upwelling to that of relaxation (fig. 5c), but with a very patchy behavior.
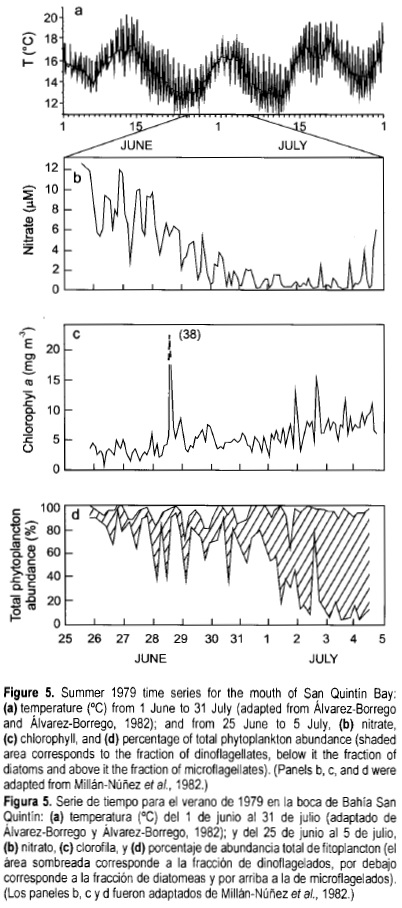
The very low temperature values and very high NO3 values during upwelling intensification events off San Quintín Bay (fig. 5) are similar to those reported for July 1979 by Simpson (1985) for waters off Point Conception. During summer, off Point Conception the CCS has a flux with equatorward and off-shore components (fig. 1). The plume of cold water off Point Conception is a feature formed by offshore advection of CCS water from the north augmented by coastal upwelling (Fiedler, 1984) . Simpson (1985) reported surface temperature values as low as 12°C and NO3 values up to 14 µM. There has been no report of this kind of intense upwelling for geographic points between Point Conception and San Quintín Bay. Consequently, it would be interesting to test the hypothesis that the sequence of upwelling events off San Quintín is due to intensification and relaxation events of the CCS circulation in this oceanic area. Cross-correlation coefficients between coastal surface water temperature and wind velocity time series (1978-1979) for two geographic locations, one between San Quintín Bay and Todos Santos Bay and the other off Todos Santos Bay (200 km to the north), were very low (Barton, 1985) , which means that the Ekman drift cannot be considered as the only generator of coastal upwelling off northwestern Baja California. Barton (1985) also reported that, in general, wind and current series were not significantly correlated; this author described equatorward pulses of currents over several days at the location closer to San Quintín Bay, coinciding with drops in sea level and not produced by any unusual change in either local or large-scale winds. There are no wind intensification events with a ~2-week period for the northwestern Baja California coast (Barton, 1985) that could explain the sequence of upwelling as described by Alvarez-Borrego and Alvarez-Borrego (1982) for the area off San Quintín.
Primary productivity (PP) measured with 14C incubations varied considerably following chlorophyll variability. In both 1977 and 1979, there were large temporal variations in the time series for the bay mouth, even with as little as 3 h difference, due to the patchy phytoplankton distribution (Lara-Lara et al, 1980; Millán-Núñez et al, 1982) (not shown). In general, data from both years are similar for the mouth, with a mean for noon and surface waters of 27 mgC m-3 h-1, or 122 mgC m -2 h-1 integrated productivity for the water column. At the mouth, PP values decreased to an order of magnitude less at 6 m compared to surface values, although more often they decreased 25-40%. In both internal extremes, while both chlorophyll and PP were about one third compared to values for the bay mouth, total phytoplankton abundance was only one tenth. This indicates that in the internal extremes, chlorophyll cell content was about triple that of the mouth. This is the result of acclimation of phytoplankton to a lower irradiance regime in the internal extremes due to greater turbidity. It is well known that phytoplankton responds to a lower irradiance regime by increasing its photosynthetic pigment content (Falkowski, 1980). Lara-Lara et al. (1980) and Millán-Núñez et al. (1982) indicated that PP values for the mouth of San Quintín Bay (up to >40 mgC m-3 h-l) are two to three times greater than values for the Gulf of California, for other upwelling areas off the west coast of Baja California and off Oregon, and they mentioned that PP at the bay mouth during summer is similar to the highest PP found in narrow bands of the nearshore Oregon upwelling system.
Based on the high turbidity of the inner arms, relatively high concentrations of inorganic macronutrients, and relatively low PP and chlorophyll values, Millán-Núñez et al. (1982) suggested that light may be the main limiting factor for phytoplankton photosynthesis. However, the Colorado River estuary waters are much more turbid than those of San Quintín Bay, and Millán-Núñez et al. (1999) reported chlorophyll and PP values of >10 mg m-3 and >40 mgC m-3 h-1, respectively, for the former. This indicates that light may not limit PP in very turbid waters if turbulence allows for some exposure of phytoplankton cells to high light levels near the water surface. An alternative explanation for low chlorophyll and PP, and high macronutrients in the internal extremes of San Quintín Bay, may be low available iron, as proposed by Martin (1992) and Anderson (2003) for the eastern equatorial Pacific waters and the Southern Ocean. Possibly other nutrient-like metals may also have very low concentration. Lares et al. (2002) studied the monthly variability of cadmium concentration in soft tissues of mussels (Mytilus californianus) at a pristine rocky shore off San Quintín Bay, and they reported that cadmium was maximum during the upwelling season and minimum in February. The ratio between the maximum and minimum concentrations was 4.6. Cadmium is a nutrient-like trace metal (10-9 mol L-1 in rich ocean water) and in the open ocean it has a high correlation with macronutrients (Libes, 1992). Higher cadmium levels during the upwelling season is a clear indication of the high concentrations of nutrient-like metals in upwelled waters off San Quintín Bay. During an upwelling intensification event, nutrient-like metals such as iron are available to induce high PP values and diatom dominance in the mouth region, but due to scavenging by adsorption onto sinking particles under oxic conditions, iron is rapidly removed and made unavailable for photosynthesis, thus limiting PP. Possibly, in the inner reaches of the lagoon there is little iron available in the water column, causing low chlorophyll values despite relatively high values of inorganic nitrogen and PO4.
The ranges in temperature for the summer 1979 and January 1980 time series were very large and greater for the mouth (fig. 6a), because of the more direct influence of the adjacent oceanic area during flood flow and of the lagoon's interiors during ebb flow. A more detailed analysis of the summer time series for the mouth reveals that, in general, the lowest temperature values correspond to high tide and upwelling water, and the highest values correspond to low tide and water from the inner reaches (Millán-Núñez et al, 1982). A comparison between the January 1980 temperature means for the inner reaches (>15°C) and those from January 1973 (<13°C) (figs. 3a, 6a) suggest an interannual variability due to meteorological variation.
The June/July 1979 salinity values ranged from a minimum of 33.4 for the mouth to a maximum of 37.6 for the eastern extreme (fig. 6a); however, unlike temperature, the larger salinity range was that of the eastern extreme. Practically all the salinity range for the eastern extreme was above that for the mouth, and it had little overlap with the one for Falsa Bay (fig. 6a), which is indicative of the much larger water residence time in the eastern extreme
The June/July 1979 NO3 mean and range for the mouth were much larger than those for the eastern arm of the lagoon, with intermediate values for Falsa Bay (fig. 6b). During upwelling relaxation periods, many relatively high NO3 values are found in the internal extremes, between 1.0 and 2.0 µM, at the same time that values at the mouth are often <1.0 µM. This implies that ammonia oxidation is a source of nitrate in the internal extremes. Ammonia reaches >7 µM in the water column of the eastern extreme, and it reaches ~3 µM at the mouth and in Falsa Bay (Farfán and Alvarez-Borrego, 1983; Osorno-Velázquez, 2000).
The June/July 1979 PO4 range was greater for the mouth than for the internal extremes, but the means were statistically the same for the three regions of the lagoon (fig. 6b). The larger number of PO4 values >1.0 µM in Falsa Bay than in the eastern extreme (fig. 5 of Millán-Núñez et al, 1982) may be due to greater turbulence caused by more intense and/or persistent waves in the former. This is consistent with Secchi disk readings at Falsa Bay, which are lower than those of the eastern arm. The June/July 1979 silicate values are consistent with those from 1973/1974 (figs. 3d, 6b).
The June/July 1979 ranges and means of chlorophyll concentration and total phytoplankton abundance were larger at the mouth than in the internal extremes, consistent with the 1973/1974 data (figs. 3e, f.6c, d). Minimum values of the three locations are similar, but at the mouth maximum values were much larger than those of the internal extremes (fig. 6c, d). Sampling every 2 h during 10 days in June and July 1979, the spatial chlorophyll distribution was clearly very patchy (fig. 5c), but even in the eastern extreme values were often >3 mg m-3.
Large period variations
Data from San Quintín Bay are very scarce. However, evidence suggests some hypothesis about interannual and interdecadal changes, and about the influence of phenomena at a mesoscale (hundreds of kilometers) and large scale (103 km). During summer, oceanic phenomena are detected at the mouth with minimum temperature and salinity values (fig. 7a, b). The most interesting data are the minimum and maximum values. Comparing the June/July data from 1973 through 1984 for the mouth and for oceanic waters near the mouth (8 km) (fig. 7), it can be seen that the minimum temperature and salinity values were very different, for example, between 1979, a non-El Niño year, and 1984, an El Niño year (fig. 7a, b).
El Niño events are detected off Baja California one and two years after their manifestation in the equatorial zone. The ENSO event that started to impact the equatorial Pacific at the beginning of 1982 was weakly manifested off southern California in December 1982 and early 1983, and in July 1983, positive temperature anomalies of up to 4°C were detected (Fiedler, 1984). In 1983 and 1984, minimum temperature values at the mouth of San Quintín Bay and 8 km off the bay were between 3.5°C and 5.0°C higher than the 1979 minimum, a non-El Niño year (fig. 7a). Minimum salinities in 1983 and 1984 had a behavior opposite to that of minimum temperatures, fluctuating between 32.4 and 32.65 in 1984 compared to 33.4 in 1979 (fig. 7b). These low minimum salinities for 1983 and 1984 are a clear indication of the suppressed upwelling during El Niño. Maximum nutrient values decreased in 1983 and 1984 to less than half the 1979 values (fig. 7c), and maximum chlorophyll values (fig. 7d) and diatom abundance decreased in 1983 and 1984 one order of magnitude relative to those for 1977 and 1979 (Torres-Moye and Alvarez-Borrego, 1985, 1987; Silva-Cota and Álvarez-Borrego, 1988).
The 1972 and 1976 ENSO events had a very strong negative impact on Peru's anchovy fishery (Barber et al, 1985). These events may have affected the San Quintín Bay area in 1973 and 1977, but the few data from the mouth do not reveal the effect with clarity. Only the July 1973 salinity (33.0) tends to be similar to the 1984 minimum salinities. Based on the salinity values, it may be deduced that the relatively high temperature values from the mouth in July 1973 were taken during flood flow (salinity and temperature samples were taken simultaneously). The 1977 salinity and temperature minima are indicative of intense upwelling, very similar to that of 1979. The 1973 and 1977 chlorophyll values (fig. 7d) represent healthy and abundant phytoplankton communities, and this contrasts with the 1983/1984 values. A possible explanation for the lack of effect from the 1973 and 1977 ENSO events on the area off San Quintín Bay is that in 1972 and 1976 the North Pacific had a "cold regime" that lasted up to 1976, after which there was a change to positive temperature anomalies throughout 1999 (Stephens et al, 2001). Rainfall generally increases in Baja California as a result of ENSO events; however, the 1972/1973 event did not cause a significantly higher rainfall than the long-term mean (S. Reyes-Coca, CICESE, pers. comm.), which also shows the effect of interdecadal variations on other geophysical variables. In other words, in this geographic area, the effect of ENSO events on primary production possibly depends on the North Pacific interdecadal oscillation.
Water parcels at the mouth with salinity <33.0 (down to 32.4) in 1984 (fig. 7b), have their origin to the north of this oceanic area. Petersen et al. (1986) studied the physical and biological characteristics of the waters over the Southern California Bight shelf (isobaths 8-75 m) for the period from June 1978 to December 1984, and reported surface salinities as low as 33.2 for 1982, 32.8 for 1983, and 33.4 for 1984, and they indicated that this low salinity waters had to come from the north. The subarctic water that feeds the CCS has surface salinities from a little more than 33.0 to 33.5 in June (Park, 1967), and from a little less than 33.6 to 34.0 during spring (Álvarez-Borrego, 1970). Interannual variation may occur due to the variability of precipitation, but its study is out of the scope of this contribution. One feasible source for these low salinities (<33.0) in the area adjacent to San Quintín Bay is a mixture of subarctic water and the Columbia River estuarine plume (Columbia River mouth is at 46°10'N). Due to the Coriolis effect, this plume should turn north, but the CCS transports it to the south and southwest during summer, so that at the latitude of Newport, Oregon (-180 km from the river mouth), its low salinity (-27) core is about 110 km from the coast (Pak et al, 1970). Off Oregon, the largest CCS meridional transport is to the south in July, and there is transport to the north during winter at irregular intervals (Stevenson et al, 1969; Hickey, 1979). The Columbia River discharges 77% of the total drainage into the Pacific between British Columbia and California (Hickey, 1979). Surface salinity summer distributions shown by Reid (1960) only depict the Columbia River plume, with no significant impact of other rivers and with relatively high salinities near the coast due to upwelling. Reid (1960) reported salinities as low as 33.2 for August 1955 in the area off Point Conception (34°35'N), -380 km from the coast, with values around 33.6 closer to the coast. Some water parcels with low salinity could be transported to the south and then to the coast by the summer onshore flux off Ensenada, and then to both San Quintín Bay and the Southern California Bight by coastal currents as shown in figure 1. Salinities as low as <33.0 may occur in relatively isolated water parcels that are possibly formed from meanders and eddies, but they would need to avoid mixing by turbulent diffusion to a certain extent in order to maintain their characteristics. These low salinity water parcels have to be relatively scarce at low latitudes, such as off San Quintín Bay, and that is why they are often not detected during cruises, when sampling localities are far apart. Of the samples collected during several days in July 1984 from the same locality, 8 km off the mouth of San Quintín Bay, 35% had salinities <33.0 (Torres-Moye and Álvarez-Borrego, 1987). The relatively abundant presence of these low salinity water parcels in 1984 off the bay mouth indicates that with the beginning of El Niño relaxation there could have been a more intense flux of the CCS towards the equator.
An alternative possibility is that these low salinity water parcels came directly from the subarctic, without the effect of the Columbia River plume, and were due to lower salinities in the subarctic because of high precipitation during those years; this is something that needs exploration. Finally, if these low salinity water parcels originate from the Columbia River estuarine plume, there are some implications concerning genetic flux between the two coastal water bodies by means of spores, eggs, etc. Furthermore, during the 1960s and up to 1971, the Columbia River estuary was a major source of radioactivity, carrying some 25,000 curies per month to the sea, due to the operation of nine plutonium reactors at Hanford. The radioactive plume could be clearly traced in the ocean 350 km to the southwest of the river mouth with 51Cr (Osterberg et al, 1965). The half-life of 51Cr is only 0.08 years, but some radioisotopes of large half-life (such as 60Co) may have reached the sediments of San Quintín Bay. This is of course a remote possibility since there is a high dilution factor due to mixing with subarctic water, and because metals can be removed by scavenging. Nevertheless, natural concentrations of radioisotopes are in the range of 10-6 to 10~29 g g-1 in ocean sediments (Libes, 1992), and the possibility of alteration of very low natural concentrations should be taken into account in future studies at San Quintín Bay.
Acknowledgements
The constructive comments and suggestions of Sergio Sañudo-Wilhelmy (Marine Sciences Research Center at Stony Brook University, New York) and two anonymous reviewers greatly helped to improve the manuscript. The drawings were produced by J.M. Domínguez and F. Ponce.
References
Aguirre-Muñoz, A., Buddemeier, R.W., Camacho-Ibar, V.F., Carriquiry, J.D., Ibarra-Obando, S.E., Massey, B., Smith, S.V. and Wulff, F. (2001). Sustainability of coastal resources in San Quintin, Mexico. Ambio, 30: 142-149. [ Links ]
Álvarez-Borrego, S. (1970). Chemico-oceanographical parameters of the central North Pacific Ocean. M.Sc. thesis, Oregon State University, Corvallis, 84 pp. [ Links ]
Álvarez-Borrego, S. y López-Álvarez, C. (1975). Distribución de biomasa de fitoplancton por grupos taxonómicos en Bahía San Quintín, B.C., a través de un ciclo anual. Reporte para el Instituto Nacional de Pesca de la SIC, y para la Dirección General de Acuacultura de la SRH. Unidad de Ciencias Marinas, Universidad Autónoma de Baja California, Ensenada. [ Links ]
Álvarez-Borrego, S. y Chee-Barragán, A. (1976). Distribución superficial de fosfatos y silicatos en Bahía San Quintín, B.C. Cieñe. Mar., 3(1): 51-61. [ Links ]
Álvarez-Borrego, S. and Nájera-de-Muñoz, S. (1979). Phytoplankton time series from two coastal lagoons of Baja California. Cienc. Mar., 6: 75-88. [ Links ]
Álvarez-Borrego, J. and Álvarez-Borrego, S. (1982). Temporal and spatial variability of temperature in two coastal lagoons. CalCOFI Rep., XXIII: 188-197. [ Links ]
Álvarez-Borrego, S., Ballesteros-Grijalva, G. y Chee-Barragán, A. (1975). Estudio de algunas variables fisicoquímicas superficiales en Bahía San Quintín, en verano, otoño e invierno. Cienc. Mar., 2(2): 1-9. [ Links ]
Álvarez-Borrego, S., Acosta-Ruiz, M.J. y Lara-Lara, J.R. (1977a). Hidrología comparativa de las bocas de dos antiestuarios de Baja California. Cienc. Mar., 4(1): 1-11. [ Links ]
Álvarez-Borrego, S., Lara-Lara, J.R. y Acosta-Ruiz, M.J. (1977b). Parámetros relacionados con la productividad orgánica primaria en dos antiestuarios de Baja California. Cienc. Mar., 4(1): 12-22. [ Links ]
Anderson, R.F. (2003). What regulates the efficiency of the biological pump in the Southern Ocean? U.S. JGOFS News, 12: 1-4. [ Links ]
Barber, R.T., Chávez, F.P. and Kogelschatz, J.E. (1985). Biological effects of El Niño. In: M. Vegas (ed.), Seminario Regional de Ciencia, Tecnología y Agresión Ambiental: El fenómeno "El Niño". Contec Press, Lima, Perú, pp. 399-138. [ Links ]
Barnard, L. (1962). Benthic marine exploration of Bahía de San Quintín, Baja California, 1960-61. General. Pac. Nat., 3: 249274. [ Links ]
Barton, E.D. (1985). Low-frequency variability of currents and temperatures on the Pacific continental shelf off northern Baja California, 1978 to 1979. Cont. Shelf Res., 4: 425-143. [ Links ]
CalCOFI Data Report (1984). Physical, Chemical and Biological data, Cruises 8404, 8405, 8406. SIO Reference 84-25. Scripps Institution of Oceanography, UCSD, La Jolla, 224 pp. [ Links ]
Camacho-Ibar, V.F. and Álvarez-Borrego, S. (1988). Nutrient concentrations in pore waters of intertidal sediments in a coastal lagoon: Patchiness and temporal variations. Sci. Total Environ., 75: 325-339. [ Links ]
Camacho-Ibar, V.F., Carriquiry, J.D. and Smith, S.V. (2003). Non-conservative P and N fluxes and net ecosystem production in San Quintin Bay, Mexico. Estuaries 26(5): 1220-1237. [ Links ]
Chávez, F.P., Ryan, J., Lluch-Cota, S.E. and Ñiquen, M. (2003). From anchovies to sardines and back: Multidecadal change in the Pacific Ocean. Science, 299: 217-221. [ Links ]
Chávez-de-Nishikawa, A. y Álvarez-Borrego, S. (1974). Hidrología de la Bahía de San Quintín, Baja California, en invierno y primavera. Cienc. Mar., 1(2): 31-62. [ Links ]
Dawson, E.Y. (1962). Benthic marine exploration of Bahía de San Quintín, Baja California, 1960-61. Marine and marsh vegetation. Pac. Nat., 2: 275-280. [ Links ]
Falkowski, P.G. (1980). Light-shade adaptation in marine phytoplankton. In: P.G. Falkowski (ed.), Primary Productivity in the Sea. Plenum Press, New York, pp. 99-119. [ Links ]
Farfán, B.C. and Alvarez-Borrego, S. (1983). Variability and fluxes of nitrogen and organic carbon at the mouth of a coastal lagoon. Estuar. Coast. Shelf Sci., 17: 599-612. [ Links ]
Fiedler, P.C. (1984). Satellite observations of the 1982-1983 El Niño along the U.S. Pacific coast. Science, 224: 1251-1254. [ Links ]
Gorsline, D.S. and Stewart, R.A. (1962). Benthic marine exploration of Bahía de San Quintín, Baja California, 1960-61. Marine and quaternary geology. Pac. Nat., 2: 281-319. [ Links ]
Hickey, B. (1979). The California Current System: Hypothesis and facts. Prog. Oceanogr., 8: 191-279. [ Links ]
Ibarra-Obando, S.E., Camacho-Ibar, V.F., Carriquiry, J.D. and Smith, S.V. (2001). Upwelling and lagoonal ecosystems of the dry Pacific coast of Baja California. In: U. Seeliger and B. Kjerfve (eds.), Coastal Marine Ecosystems of Latin America. Springer, Berlin, pp. 315-329. [ Links ]
Kjerfve, B. (1994). Coastal Lagoon Processes. In: B. Kjerfve (ed.), Coastal Lagoon Processes, Elsevier, Amsterdam, pp. 1-15. [ Links ]
Knoppers, B. (1994). Aquatic primary production in coastal lagoons. In: B. Kjerfve (ed.), Coastal Lagoon Processes. Elsevier, Amsterdam, pp. 243-286. [ Links ]
Lara-Lara, J.R. y Álvarez-Borrego, S. (1975). Ciclo anual de clorofilas y producción orgánica primaria en Bahía San Quintín, B.C. Cienc. Mar., 2(1): 77-97. [ Links ]
Lara-Lara, J.R., Álvarez-Borrego, S. and Small, L.F. (1980). Variability and tidal exchange of ecological properties in a coastal lagoon. J. Estuar. Coast. Mar. Sci., 11: 613-637. [ Links ]
Lares, M.L., Flores-Muñoz, G. and Lara-Lara, J.R. (2002). Temporal variability of bioavailable Cd, Hg, Zn, Mn, and Al in an upwelling regime. Environ. Pollut., 120: 595-608. [ Links ]
Libes, S.M. (1992). An Introduction to Marine Biogeochemistry. John Wiley, New York, 733 pp. [ Links ]
Martin, J.H. (1992). Iron as a limiting factor in oceanic productivity. In: P.G. Falkowski and A.D. Woodhead (eds.), Primary Productivity and Biogeochemical Cycles in the Sea. Plenum Press, New York, pp. 123-138. [ Links ]
Martori-Oxamendi, J.I. (1989). Variabilidad de la circulación y sus causas en Bahía San Quintín, B.C. Tesis de maestría en ciencias, CICESE, Enseanda, 110 pp. [ Links ]
Millán-Núñez, R., Álvarez-Borrego, S. and Nelson, D.M. (1982). Effects of physical phenomena on the distribution of nutrients and phytoplankton productivity in a coastal lagoon. Estuar. Coast. Shelf Sci., 15: 317-335. [ Links ]
Millán-Núñez, R., Santamaría-del-Angel, E., Cajal-Medrano, R. and Barocio-León, O.A. (1999). The Colorado River Delta: A high primary productivity ecosystem. Cienc. Mar., 25: 509-524. [ Links ]
Ocampo-Torres, F.J. (1980). Análisis de mareas y predicción de velocidad mediante un modelo unidimensional en Bahía San Quintín, B.C. Tesis profesional, Universidad Autónoma de Baja California, Ensenada, 94 pp. [ Links ]
Osorno-Velázquez, A.I. (2000). Impacto del cultivo de moluscos filtroalimentadores en el fitoplancton y calidad del agua de Bahía San Quintín. Tesis de maestría en ciencias, CICESE, Ensenada, 48 pp. [ Links ]
Osterberg, C, Cutshall, N. and Cronin, J. (1965). Chromium-51 as a radioactive tracer of Columbia River water at sea. Science, 150: 1585-1587. [ Links ]
Pak, H., Beardsley, Jr., G.F. and Park, P.K. (1970). The Columbia River as a source of marine light-scattering particles. J. Geophys. Res., 75: 4570-1578. [ Links ]
Park, P.K. (1967). Chemical features of the subarctic boundary near 170°W. J. Fish. Res. Board Canada, 24: 899-908. [ Links ]
Peláez, J. and McGowan, J.A. (1986). Phytoplankton pigment patterns in the California Current as determined by satellite. Limnol. Oceanogr.,31(5): 927-950. [ Links ]
Petersen, J.H., Jahn, A.E., Lavenberg, R.J., McGowen, G.E. and Grove, R.S. (1986). Physical-chemical characteristics and zooplankton biomass on the continental shelf off southern California. CalCOFI Rep., XXVII: 36-52. [ Links ]
Reid, J.L. (1960). Oceanography of the northeastern Pacific Ocean during the last ten years. CalCOFI Rep., VII: 77-90. [ Links ]
Silva-Cota, S. and Álvarez-Borrego, S. (1988). The "El Niño" effect on the phytoplankton of a north-western Baja California coastal lagoon. Estuar. Coast. Shelf Sci., 27: 109-115. [ Links ]
Simpson, J.J. (1985). Air-sea exchange of carbon dioxide and oxygen induced by phytoplankton. Methods and interpretation. In: A. Zirino (ed.), Mapping Strategies in Chemical Oceanography. American Chemical Soc, Washington, D.C., pp. 409-450. [ Links ]
Smith, S.V. and Hollibaugh, T. (1997). Annual cycle and interannual variability of ecosystem metabolism in a temperate climate embayment. Ecol.Monogr., 67: 509-533. [ Links ]
Stephens, C, Levitus, S., Antonov, J. and Boyer, T.P. (2001). On the Pacific Ocean regime shift. Geophys. Res. Lett., 28: 3721-3724. [ Links ]
Stevenson, M., Pattullo, J.G. and Wyatt, B. (1969). Subsurface currents off the Oregon coast as measured by parachute drogues. Deep-Sea Res., 16: 449-161. [ Links ]
Teague, K.G., Madden, C.J. and Day, J.W. Jr. (1988). Sediment-water oxygen and nutrient fluxes in a river-dominated estuary. Estuaries, 11: 1-9. [ Links ]
Torres-Moye, G. and Álvarez-Borrego, S. (1985). The 1983 "El Niño" effects on nutrients and summer phytoplankton of western Baja California coastal waters. Cienc. Mar., 11: 107-113. [ Links ]
Torres-Moye, G. and Álvarez-Borrego, S. (1987). Effects of the 1984 El Niño on the summer phytoplankton of a Baja California upwelling zone. J. Geophy. Res. (Oceans), 92(C13): 14383-14386. [ Links ]
Usui, T., Koike, I. and Ogura, N. (2001). N20 production, nitrification and denitrification in an estuarine sediment. Estuar. Coast. Shelf Sci., 52: 769-781. [ Links ]

