










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkCiencias marinas
versão impressa ISSN 0185-3880
Cienc. mar vol.29 no.3 Ensenada Set. 2003
Artículos
Interacción entre el lobo marino de California y la pesquería artesanal en la Bahía de La Paz, Golfo de California, México
Interaction between the California sea lion and the artisanal fishery in La Paz Bay, Gulf of California, Mexico
David Aurioles-Gamboa1*, Francisco García-Rodríguez1, Mauricio Ramírez-Rodríguez1 y Claudia Hernández-Camacho1
1 Departamento de Pesquerías y Biología Marina, Centro Interdisciplinario de Ciencias Marinas, Apartado postal 592, La Paz, Baja California Sur, México. *E-mail: dgamboa@ipn.mx
Resumen
Se analizó información acerca de las presas del lobo marino de California (Zalophus californianus californianus) y la composición de la captura de la pesquería artesanal alrededor de la lobera de Los Islotes, Bahía de La Paz, México, para estimar la superposición y competencia potencial entre estos consumidores. Los lobos marinos consumen cerca de 435 t anualmente, con un pico de consumo en el invierno. La pesquería artesanal extrae cerca de 730 t de peces óseos anualmente, registrándose un incremento en verano. De acuerdo con muestreos que cubren cinco años entre 1980 y 1994, los lobos marinos depredaron sobre un total de 76 especies de peces; sin embargo, en términos de importancia relativa (IR), el 35% estuvo concentrado en cinco presas, de las cuales sólo una tiene valor comercial en el área. Los principales objetivos de la pesquería se centraron en cuatro familias que significaron el 76.2% de la captura total e incluyeron 28 especies. De estas especies, únicamente dos aparecieron en la materia fecal de lobo marino, siendo la cabrilla arenera (Paralabrax maculatofasciatus) la única con valor comercial significativo, pero ésta estuvo pobremente representada en la dieta del lobo marino (< 5% IR). A pesar de la poca superposición en sus objetivos de pesca, la población de Los Islotes presenta el mayor índice de enmalle (7-9%) de todas las colonias de lobo marino estudiadas en México. Todas las categorías sexo/edad de lobo marino tuvieron índices de enmalle similares, siendo la fracción de lobos marinos enmallados aproximadamente proporcional en estructura a la población total, durante tres años distintos. Estos datos sugieren que la mayoría de los animales se enmallan accidentalmente y al azar. El enmalle se debe probablemente al efecto combinado del amplio uso de redes agalleras en la bahía y la superposición de las áreas de alimentación de los lobos marinos y las áreas operativas de los pescadores.
Palabras clave: lobo marino de California, pesquería artesanal, hábitos alimentarios, enmalle, México.
Abstract
Information about California sea lion prey and the artisanal fishery catch in the area of the Los Islotes sea lion rookery, in northeastern La Paz Bay, México, was examined to estimate potential overlap and competition between these consumers. Sea lions there consume about 435 t annually, with peak consumption in winter. The artisanal fishery captures about 730 t of bony fish annually, with an increase in summer. Based on a five-year study, sea lions preyed on 76 fish species, but in terms of relative importance (RI), 35% of these was represented by only five species, of which only one has regional commercial value. Principal fishery targets comprised 28 species in four families, representing 76.2% of the total capture. Of these, the remains of only two species appeared in sea lion scats, and of them, only the spotted sand bass, Paralabrax maculatofasciatus, has significant commercial value, but it is poorly represented as sea lion prey (< 5% RI). Despite the limited overlap with fishery target species, the Los Islotes population exhibited the highest entanglement index (7-9%) of all sea lion colonies studied in Mexico. All sex/age categories of sea lions showed similar entanglement indices, with the entangled fraction approximately proportional to the total population for three surveys in different years. The data suggest that most animals become entangled by accident. Entanglement is probably due to the combined effect of extensive gillnet deployment in the bay and overlap in the areas where sea lions feed and fishermen operate.
Key words: California sea lion, artisanal fishery, feeding habits, entanglement, Mexico.
Introducción
La interacción creciente entre las pesquerías y las poblaciones de mamíferos marinos es de gran interés, no sólo porque es uno de los principales problemas para su conservación (Northridge, 1985; Northridge y Hofman, 1999), sino también porque es difícil de evaluar y manejar racionalmente. Los pinnípedos, particularmente los otáridos, interactúan con las pesquerías ribereñas debido a que se reproducen en islas costeras y están asociados con áreas de alta productividad, como las zonas de surgencias (Costa, 1993), donde también operan muchas pesquerías.
Entre las especies que interactúan con las pesquerías costeras se encuentra el lobo marino de California, Zalophus californianus californianus Lesson, 1828. Éste interactúa con la pesquería del salmón (Bigg et al., 1990; Fraker y Mate, 1999); la merluza del Pacífico, Merluccius productus Ayres, 1855 (Fiscus, 1979; Ainley et al., 1982); rocotes, Sebastes spp.; el calamar, Loligo opalescens Berry, 1911 (Fiscus, 1980; Jones, 1981); el arenque del Pacífico, Engraulis mordax Girard, 1854 (Fiscus, 1980); y la sardina Monterrey, Sardinops sagax Girard, 1854 (Aurioles-Gamboa y García-Rodríguez, 1999).
En el Golfo de California, las principales industrias pesqueras están basadas en la sardina Monterrey y el camarón. La pesquería artesanal, sin embargo, es muy común y constituye una importante fuente de productos y empleos. Ésta se caracterizan por ser una extensa flota regional de pequeños botes (pangas) de aproximadamente 7 m de largo. Los pescadores artesanales entran a menudo en conflicto con una variedad de mamíferos marinos, pero es el lobo marino de California, el único pinnípedo que se reproduce dentro del Golfo de California (Le Boeuf et al., 1983), con el que hay más interacción. En este conflicto, los lobos marinos son afectados principalmente al enmallarse en las redes de pesca causándoles la muerte directamente por asfixia en inmersión o indirectamente debido a las heridas provocadas durante el escape. En la Bahía de La Paz el enmalle puede afectar hasta 5-7% de la población (Harcourt et al., 1994), mientras que en algunas colonias del norte del Golfo de California este se ha registrado entre 0-2.2% (Zavala y Mellink, 1997).
Para las pesquerías artesanales, la pérdida económica es representada por los peces consumidos o dañados, las redes rotas o desechadas y el lucro cesante reparando redes y por consumo de gasolina.
En la Bahía de La Paz se usan diversas artes de pesca. De encuestas realizadas a 59 pescadores, los artes más comunes fueron línea y anzuelo (96.6%), redes agalleras de diferente luz de malla (25.1%) y buceo libre (3.38%) (SEMARNAT, 2000). Las redes agalleras son colocadas generalmente durante la noche, entre 2 y 30 m de profundidad. La mayoría de las redes son de monofilamento y están diseñadas para especies de peces óseos. Hay cerca de 18 diferentes tipos de redes en uso, dependiendo de luz de malla, área, coeficiente de estiramiento y diámetro de la línea principal. El tamaño de malla varía entre 20 y 200 mm, pero la más frecuente es entre 60 y 200 mm (Ramírez-Rodríguez, 1991).
La Bahía de La Paz representa un buen caso de estudio de las interacciones lobo marino-pesquerías, debido a que la única pesquería en la bahía es de tipo artesanal, y a que el único pinnípedo en el área es el lobo marino de California. Aunque entran en la Bahía de La Paz otras tres especies de pinnípedos (elefante marino Mirounga angustirostris, foca común Phoca vitulina y lobo fino de Guadalupe Arctocephalus townsendi; Aurioles-Gamboa et al., 1993), su presencia es infrecuente y representada por individuos solitarios que tienen una mínima contribución a la interacción pinnípedos-pesquerías. Por un lado, existe información sobre la captura total de peces (biomasa), el tipo de arte de pesca, y la variación específica de la captura a lo largo del año (Ramírez-Rodríguez, 1991; Ramírez-Rodríguez, datos no publicados); por otro lado, Los Islotes es una colonia de lobo marino monitoreada desde 1978 donde se han realizado varios estudios sobre hábitos alimentarios, enmalle y dinámica poblacional. Esta información fue analizada conjuntamente para evaluar la interacción entre las pesquerías artesanales y los lobos marinos en la Bahía de La Paz.
En el presente trabajo se abordaron dos aspectos de la interacción entre lobos marinos y pesquería artesanal: (1) se determinó la superposición entre los hábitos del lobo marino y la composición de la pesquería y se compararon los niveles de consumo de alimento del lobo marino y la captura de las pesquerías; y (2) se midió el grado de enmalle de lobos marinos durante dos estaciones reproductivas (1998 y 2000) y se compararon con un estimado para 1992 con el objeto de determinar la importancia del problema y su tendencia en la Bahía de La Paz.
Materiales y métodos
Tamaño y estructura de la población de lobo marino de California en Los Islotes
Se realizaron mensualmente censos directos de la población por categorías de sexo y edad en la lobera de Los Islotes, en la boca de la bahía (fig. 1). Cada categoría está integrada por animales con las siguientes características morfológicas relacionadas con la edad y el sexo: machos adultos, machos sub-adultos, hembras adultas, juveniles y crías. La diferenciación y descripción morfológica se basó en diversos trabajos anteriores (Peterson y Bartholomew, 1967; Lluch, 1969; Orr et al., 1970; Le Boeuf et al., 1983).

Para determinar la fluctuación media de la población a lo largo de un año típico, se integró una base de datos con 110 censos poblacionales mensuales desde 1978 hasta 1986. La base de datos proviene de información ya publicada (Aurioles-Gamboa et al., 1983; Aurioles-Gamboa, 1988; Aurioles-Gamboa y Sinsel, 1988; Aurioles-Gamboa y Le Boeuf, 1991), y de datos aún no publicados.
Dado que no se aplicaron correcciones a los censos, existe una subestimación debido a que parte de la población se encuentra en el mar alimentándose. Asumimos que este error en los censos se repite en todos los muestreos, por lo tanto la composición de la población y su variación pueden ser aun interpretadas (Aurioles-Gamboa, 1988; Aurioles-Gamboa y Zavala, 1994).
Hábitos alimentarios
Se integró una base de datos de los hábitos alimentarios de lobo marino a partir de cuatro años de muestreos de su materia fecal (Aurioles-Gamboa et al., 1984; García-Rodríguez, 1995; Aurioles-Gamboa, datos no publicados) para determinar el espectro de alimentación promedio mensual de la población del lobo marino de Los Islotes. Se recolectaron mensualmente, con algunas excepciones, todas las deposiciones de materia fecal fresca de lobo marino disponibles en la lobera (cada muestra fue almacenado en una bolsa de plástico etiquetada y sin líquidos conservantes). Esta tarea fue realizada en meses discontinuos de acuerdo con el siguiente calendario: 1980 (septiembre y diciembre; 93 muestras); 1981 (enero, abril, junio, agosto, noviembre y diciembre; 172 muestras); 1982 (enero y abril; 27 muestras); 1990 (febrero, abril, junio, agosto y septiembre; 163 muestras); y 1993 (febrero, abril, junio y noviembre; 445 muestras).
En el laboratorio, la materia fecal fue separada utilizando una serie de tamices de 2.0, 1.19, 0.71 y 0.59 mm de luz de malla y bajo el chorro de agua. Todos los otolitos de peces y picos de cefalópodos fueron separados y mantenidos en seco para su posterior identificación.
La identificación de las presas se realizó con base a nuestra colección de otolitos de más de 110 especies, así como a referencias de la literatura (Fitch, 1964, 1966, 1968; Fitch y Brownell, 1968). Para los ejemplares no representados en nuestra colección o en las referencias fotográficas, se consultó la colección de otolitos del Museo de Historia Natural del Condado de Los Ángeles.
La importancia de cada presa en los hábitos alimentarios de los lobos marinos se basó en la abundancia relativa (AR) de sus otolitos en las heces: AR = Ni x 100/N, donde Ni es el número de otolitos de la especie i y N es el número total de otolitos identificados. Existe un sesgo en esta aproximación debido a dos problemas principales: diferencias en biomasa entre las especies de peces, que pueden ser corregidas cuando existen regresiones del tamaño del otolito versus la longitud y peso del cuerpo del pez y, por lo tanto, cada otolito puede ser transformado en unidades de biomasa (Wyllie-Echeverría, 1987; Aurioles-Gamboa, 1991). También puede existir un sesgo debido a las diferencias potenciales en la digestión del otolito en el tracto del lobo marino, debido al tamaño y la composición mineral del otolito, por lo que puede subestimarse el número de otolitos presentes en cada muestra fecal, afectando así la importancia de la presa. Ninguno de estos sesgos fue considerado en el presente estudio, al igual que en muchos otros, debido a la falta de las regresiones mencionadas y de valores de digestión diferencial para la mayoría de los peces encontrados como presas.
Estimación del consumo de los lobos marinos
Peso promedio de los lobos marinos
Debido a que los censos por categoría de edad y sexo están basados en las características dimórficas y en la talla de los animales, éstos pueden ser utilizados para estimar el peso de cada categoría y aplicar tasas de consumo de alimento diferenciales para obtener estimaciones más aproximadas del consumo de alimento de la población. Se utilizó información de la literatura para determinar el peso promedio de cada categoría (Scheffer, 1958; Peterson y Bartholomew, 1967; Lluch, 1969; Odell, 1975; King, 1983; Aurioles-Gamboa, 1988; Perez et al., 1990). El peso promedio para los machos adultos fue de 265 kg, para los machos subadultos de 180 kg, para las hembras adultas de 85 kg y para los juveniles de 38kg. Las crías fueron excluidas debido a que dependen principalmente de la leche de sus madres durante el primer año de vida.
Estimaciones del consumo de alimento
Perez et al. (1990) obtuvieron amplia información sobre el consumo de alimento de mamíferos marinos en cautiverio. De 13 hembras de lobo marino de California entre 54 y 118 kg de masa corporal, la cantidad de alimento ingerido por día fue equivalente al 5.15% (± 0.58%) de su masa corporal, lo cual es probablemente bajo comparado con animales en vida libre que gastan energía en obtener su comida, particularmente las hembras lactantes. Costa (1991), utilizando agua isotópica doblemente etiquetada en experimentos con hembras lactantes de 85 kg de peso, estimó un consumo de alimento promedio de 60 g por buceo y 202 buceos por día. La masa de alimento ingerido diariamente fue igual a 12.12 kg, equivalente a 14.25% de su peso corporal. Este consumo diario fue 2.7 veces mayor que el estimado para las hembras en cautiverio (Perez et al., 1990).
No existen estimaciones del consumo de alimento basadas en estudios metabólicos para las otras categorías (hembras no lactantes, machos subadultos y machos adultos de lobo marino de California), excepto para juveniles. Los animales incluidos en esta categoría pesaron cerca de 36 kg y presentaron un consumo de alimento diario de 10.1% respecto a su masa corporal (Costa, 1987, 1991). Por lo tanto, para un juvenil de 40 kg de peso, el consumo de alimento diario estimado es de aproximadamente 4.04 kg. Dado que los juveniles usualmente tienen tasas de consumo de alimento 1.4 veces mayores a las de los adultos (Innes et al., 1987), estimamos la ingestión de alimento diario para las otras categorías en 7.2% de su masa corporal, por lo que para hembras adultas no lactantes resultó 6.12 kg/día, para machos subadultos 13 kg/día y para machos adultos 19 kg/día.
Ingestión de alimento promedio mensual por categoría
Para estimar el consumo de alimento por mes (CPM) se utilizaron las tasas de consumo de alimento junto con el número de lobos marinos en cada categoría de sexo y edad de la siguiente forma: CPM = ID x 30 x TPi, donde CPM es el consumo de alimento promedio mensual, ID es la tasa de ingestión diaria, 30 es el número promedio de días por mes y TPi es el tamaño poblacional promedio para la categoría i en cada mes. Se aplicaron las siguientes suposiciones a la población de lobo marino de California: (1) las crías pueden ser excluidas debido a que dependen principalmente de sus madres durante el primer año de vida (Peterson y Bartholomew, 1967); (2) el número de hembras lactantes en cada mes es igual al número de crías vivas registradas para el mismo mes en la lobera; y (3) los machos adultos no se alimentan durante la temporada reproductiva, de mediados de mayo a mediados de agosto (Peterson y Bartholomew, 1967).
El consumo de alimento mensual total por la población de lobo marino en la Bahía de La Paz fue distribuido entre las diferentes presas en proporción a la importancia relativa de cada presa durante ese mes.
Captura y composición de la pesquería artesanal
Se contó con los registros de todos los peces capturados anualmente en la Bahía de La Paz (fig. 1) durante el periodo 1987-1990, cubriendo todos los campos pesqueros y áreas de pesca de la bahía. Cada reporte de captura incluía especie, fecha, biomasa (kilogramo) y localidad de la captura (Ramírez-Rodriguez, 1997). La captura está compuesta de varias especies de peces (Osteichtyes y Elasmobranchii) y también de varias especies de crustáceos y moluscos. Sólo se utilizaron las capturas de peces óseos para la comparación, debido a que reportes previos indican que más del 90% del alimento de los lobos marinos en la Bahía de La Paz son peces óseos (Aurioles-Gamboa et al., 1984; Aurioles-Gamboa, 1988; García-Rodríguez, 1995; Orr, 1998).
Incidencia de lobos marinos enmallados
Se examinó la población de lobo marino de Los Islotes para determinar la presencia de restos de redes o cicatrices producidas por el enmallamiento. Cada animal afectado fue identificado con fotografías y dibujos. El registro de animales enmallados se realizó en las temporadas reproductivas de 1998 y 2000 en Los Islotes y se estimó la frecuencia de lobos marinos en cada categoría de sexo y/o edad específica, y luego esta estructura se comparó con la población total de lobo marino observada en Los Islotes. Existe una estimación previa de lobos marinos con marcas de enmalle hecha en 1992 (Harcourt et al. 1994); sin embargo, estos datos estaban basados en censos ajustados. Para comparar nuestros datos y otros de la literatura con la estimación de 1992, usamos la lista de animales enmallados reportados por Harcourt et al. (1994), pero la comparamos con los censos de población de 1992 de nuestra base de datos para dicho año. Posteriormente, calculamos la proporción de la población de lobo marino que mostraba signos de enmallamiento viejo o reciente (PE) para 1992, 1998 y 2000, como PE = IE x 100/ PT, donde IE, es el número de individuos con marcas de red y PT es el tamaño poblacional total de lobos marinos censado en Los Islotes en cada visita (año).
Resultados
Fluctuación poblacional a lo largo del año
La variación en la población total de lobo marino de California de Los Islotes entre 1978 y 1986 (fig. 2a) revela dos picos de abundancia: uno en verano y otro en invierno. Estos picos han sido descritos previamente (Aurioles-Gamboa et al., 1983; Aurioles-Gamboa, 1988) y coinciden parcialmente con fluctuaciones observadas en otras áreas (Odell, 1975). El pico poblacional de verano es producido por la presencia de hembras reproductoras y sus crías. Las hembras adultas están presentes a lo largo de todo el año, pero se incrementan en número de mayo a septiembre. El pico de invierno es generado principalmente por los machos subadultos (fig. 2b), que son abundantes de noviembre a marzo y luego disminuyen a lo largo de la primavera y el verano.
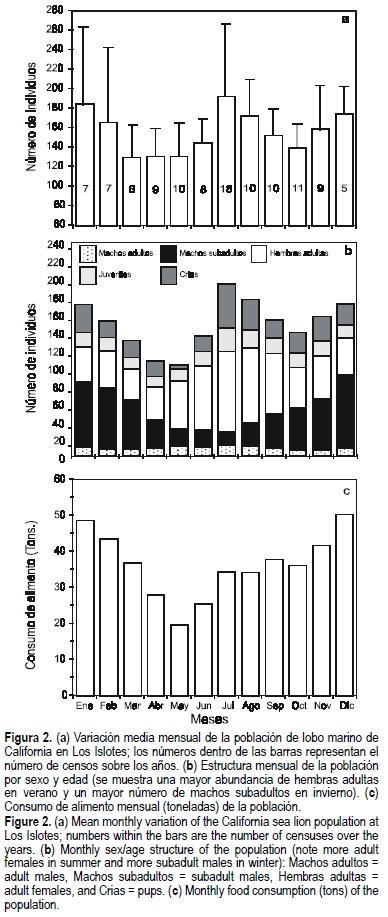
Consumo de alimento por los lobos marinos a lo largo del año
La biomasa mensual consumida por la población de lobo marino en Los Islotes se estimó en alrededor de 20 a 50 t y el consumo de alimento anual total fue estimado en cerca de 435 t. Con base en la estructura de la población en cada mes, en la figura 2c se muestra la fluctuación del consumo de alimento. Las hembras adultas tienen su máximo consumo de alimento cuando la mayoría de las hembras lactantes está presente, y luego éste empieza a disminuir hasta mayo. Mayo presenta el menor consumo debido a una disminución general en el número de individuos de todas las categorías, previa a la temporada reproductiva. El mayor consumo de alimento por los lobos marinos ocurre en invierno y es determinado en su mayor parte por los machos subadultos (diciembre y enero), quienes consumen entre 60% y 80% de las aproximadamente 50 t consumidas en cada uno de esos meses por toda la población (fig. 2c).
Composición de la dieta de los lobos marinos
Se reconocieron un total de 75 especies como presas del lobo marino en los Islotes. La figura 3 muestra las cinco presas preferenciales (de 15% a 4% en importancia relativa). Las 71 presas restantes (cada una con menos del 2%) formaron un grupo que representó cerca del 65% de importancia relativa.

La presencia de estas presas fue consistente a través de los años de muestreo, subrayando la relevancia de estas presas en la dieta del lobo marino; sin embargo, estos peces fluctuaron en abundancia a lo largo del año como se muestra en la figura 4. Una de las principales especies es el pez serpiente Aulopus bajacali Parin y Kotlyar, 1984, que habita en profundidades de 80 a 230 m, y que algunas veces es encontrada en montañas submarinas (guyots) y en bancos de mar abierto. La fluctuación mensual en la abundancia de A. bajacali a lo largo del año (fig. 4a) es semejante a la fluctuación del número de machos subadultos en Los Islotes (fig. 2b). Esta especie no tiene valor comercial en el área y se conoce poco sobre su ecología y biología. El serrano Pronotogrammus eos Gilbert, 1890, junto con P multifasciatus Gill, 1863, fueron presas recurrentes en la materia fecal del lobo marino. El primero fue consumido en un patrón irregular a lo largo del año (fig. 4b), mientras que el segundo fue más abundante en los copros de febrero a agosto (fig. 4c). Pronotogrammus mutifasciatus habita en profundidades de alrededor de 40 a 200 m (Robins et al., 1991), mientras que P. eos es encontrado entre 155 y 325 m (Heemstra, 1995).

Especies comerciales y su consumo por lobos marinos
La biomasa anual total extraída por la pesquería artesanal fue de alrededor de 1380 t, de las cuales cerca de 730 t correspondieron a peces que son presas potenciales de los lobos marinos. La tendencia de la captura mensual de peces se muestra en la figura 5, observándose un incremento en la biomasa de enero a julio, seguido de un descenso hacia diciembre. El incremento en la captura por la pesquería artesanal durante el verano siguió un patrón opuesto al menor consumo de alimento por los lobo marinos durante el mismo periodo (fig. 2c).

Las principales especies de peces capturados por la pesquería artesanal durante el periodo de estudio se muestran en la figura 6 (no incluye moluscos y tiburones). Entre las principales familias se encontraron los Lutjanidae, Serranidae, Malacanthidae y Carangidae. Estas cuatro familias representaron el 76.2% de la captura total en la bahía. Las especies incluidas en estas familias y las principales especies que son presas del lobo marino se muestran en la tabla 1. De las 28 especies capturadas comercialmente, únicamente dos (7.14%) aparecieron en las muestras fecales de lobo marino, siendo la cabrilla arenera, Paralabrax maculatofasciatus Steindachner, 1868, la única con valor comercial significativo. Sin embargo, esta especie representó menos del 5% de importancia relativa dentro de la dieta del lobo marino (fig. 3).


Enmallamiento de lobos marinos
Durante 1998 se realizaron tres visitas mensuales a la lobera (junio, julio y agosto) de seis días cada una. Durante esos viajes, se registraron un total de 17 animales (12 hembras adultas, 1 juvenil, 3 machos subadultos y 1 macho adulto) con restos de red en cuellos o cabezas, con heridas recientes o con cicatrices viejas. En el periodo de mayo a agosto de 2000, se realizaron cuatro viajes mensuales de cuatro días cada uno, identificándose un total de 19 individuos afectados por redes de pesca (16 hembras adultas, 1 juvenil, 1 macho subadulto y 1 macho adulto).
La distribución por sexo y edad de la población enmallada, comparada con la estructura de la población total observada en Los Islotes (fig. 7), no fue significativamente diferente en ninguno de los tres años (incluido 1992). El número de animales enmallados de cada categoría tuvo una alta correlación con las categorías similares de la población total (r = 0.97; g.l. 10; P > 0.028; R2 = 95%), un análisis de ji-cuadrada confirmó que las dos series de categorías de sexo y edad mostraron resultados similares (1992: Χ2 = 1.61, g.l. 3; 1998: Χ2 = 1.79, g.l. 3; 2000: Χ2 = 0.63, g.l. 3; P < 0.05). Estos datos sugieren que los individuos de cualquier categoría tuvieron la misma probabilidad de quedar enmallados.

El porcentaje de lobos marinos que mostraron marcas de enmalle, relativo al total de la población, fue muy cercano en los tres años de muestreo, observándose una ligera disminución en los siguientes años: 1992 = 10.4%, 1998 = 9.9%, y 2000 = 8.75%.
Tipos de redes involucradas en el enmalle
Algunos lobos marinos de Los Islotes fueron rescatados para eliminar los restos de redes de sus cuerpos y determinar el tipo de material. En la tabla 2 se muestran los datos relacionados con las capturas. Todos los animales fueron capturados con red tipo mariposa y fueron liberados inmediatamente después de quitarles el material de pesca, de ese modo se redujo el manejo a un tiempo promedio de 15 min.
Discusión
La fluctuación en la población de lobo marino de Los Islotes fue consistente año con año, y estuvo influenciada por dos factores principales: (1) la estación reproductiva (verano), cuando hembras adultas, juveniles y los crías son más abundantes; y (2) el pico de invierno, cuando los machos sub-adultos inmigrantes son abundantes y tienen un fuerte impacto sobre el consumo de alimento total de la población. Durante la temporada reproductiva, el consumo de alimento fue menor que en el invierno. Esto se debe a que las crías contribuyen al incremento de la población, pero se encuentran en su periodo de lactación.
En verano, la mayoría del consumo se debe a las hembras adultas y a los juveniles, concentrándose en dos especies de peces del género Pronotogrammus. Diciembre y enero fueron los meses con el mayor consumo de alimento, debido en este caso a la presencia de los machos subadultos. Mayo tuvo el menor consumo de alimento debido a que en ese mes, tanto los machos subadultos como las hembras adultas estuvieron presentes en menor número.
La especie que apareció con mayor frecuencia en la materia fecal del lobo marino fue el pez serpiente A. bajacali, la cual también fue su presa más importante en un estudio realizado en 1994-1995 (Orr, 1998). La especie fue consumida principalmente en el invierno y no se encontraron restos en los copros recolectados en junio y julio. Un patrón similar fue reportado por Orr (1998), aunque este autor encontró menos restos de Aulopus durante la estación reproductiva (número 12 en el rango de importancia). La aparición de restos de A. bajacali a principios de agosto y su posterior incremento coinciden con la llegada de los machos subadultos a Los Islotes y es equiparable al incremento en el número de machos subadultos. Aunque circunstancial, el patrón sugiere que A. bajacali deber ser principalmente consumido por los machos subadultos.
En general, la información registrada sugiere poca superposición sobre los recursos pesqueros entre la pesquería artesanal y la población de lobo marino. Esta conclusión está basada en los siguientes argumentos. (1) Los lobos marinos en la Bahía de La Paz depredaron principalmente sobre especies que no fueron objetivo de los pescadores artesanales; A. bajacali y Pronotogrammus spp. son habitantes de aguas moderadamente profundas, quedando fuera del rango de pesca de los pescadores, lo que explicaría, al menos parcialmente, por qué estas especies no son capturadas por la pesquería arte-sanal. (2) El consumo del lobo marino a lo largo del año tuvo una tendencia opuesta a la de la captura de la pesquería artesanal, indicando que cuando los machos subadultos se incrementan en número y el consumo de alimento total por el lobo marino es mayor, la pesquería disminuye su esfuerzo de captura; los pescadores artesanales reducen la pesca de noviembre a abril debido a que los vientos intermitentes del noreste causan mal tiempo (Ramírez-Rodríguez, 1997). (3) Las dos especies comerciales que estuvieron presentes en la materia fecal del lobo marino tuvieron una baja frecuencia en la dieta y un valor comercial significativo.
En contraste con esta última evidencia, los lobos marinos en La Bahía de La Paz presentan el mayor índice de enmalle (8.75-10.4%) en el Golfo de California (Harcourt et al., 1994; Zavala y Mellinck, 1997) y con valores varias veces mayores al 0.08% reportado en California (Stewart y Yochem, 1987) y al 0.002% registrado en las Islas San Benitos (Aurioles-Gamboa, datos no publicados). El amplio uso de redes agalle-ras en la Bahía de La Paz es probablemente la causa principal de la alta tasa de enmalle de lobos marinos, particularmente durante los viajes de alimentación nocturnos, cuando el nylon no es detectado. Los lobos marinos parecen enmallarse por accidente, ya que la estructura de edad y sexo similar entre la población enmallada y la población total de Los Islotes sugiere que todas las clases se enmallan con la misma probabilidad.
En el verano, las dos categorías que sufren mayores tasas de enmalle son las hembras adultas y los juveniles. Esto coincide con el momento en que la pesquería artesanal incrementa su esfuerzo y hay más redes agalleras en la bahía, mientras que la población de lobo marino se está reproduciendo y las hembras adultas y los juveniles son los principales depredadores de la colonia. Esta concentración temporal de redes agalleras puede tener un efecto negativo en la nutrición y la supervivencia de las hembras adultas, afectando la supervivencia de sus crías.
Pese a estas capturas incidentales no se ha detectado ningún efecto negativo aparente en la dinámica de la población de Los Islotes. La población de lobo marino de Los Islotes ha mostrado un incremento constante en los últimos diez años (Castro-González et al., 2001). Sin embargo, si la tasa de enmalle incrementa y continúa afectando a las hembras adultas y a los juveniles, es de esperarse efectos perjudiciales a largo plazo.
Referencias
Ainley, D.G., Huber, R.H. and Bailey, M.K. (1982). Population fluctuations of California sea lions and the Pacific whiting fishery off Central California. Fish. Bull., 80: 253-258. [ Links ]
Aurioles-Gamboa, D. (1988). Behavioral ecology of California sea lions in the Gulf of California. Ph.D. thesis, University of California, Santa Cruz, 175 pp. [ Links ]
Aurioles-Gamboa, D. (1991). Otolith size versus weight and body length relationships for some fishes off the Pacific coast of Baja California Sur, Mexico. Fish. Bull., 89: 701-706. [ Links ]
Aurioles-Gamboa, D. and Sinsel, F. (1988). Mortality of California sea lion pups at Los Islotes, Baja California Sur, Mexico. J. Mammal., 69(1): 180-183. [ Links ]
Aurioles-Gamboa, D. and Le Boeuf, B.J. (1991). Effects of the El Niño 1983 on the California sea lion population in Mexico. In: F. Trillmich and K. Ono (eds.), Pinnipeds and El Niño Responses to Environmental Stress. Springer-Verlag. 293 pp. [ Links ]
Aurioles-Gamboa, D. y Zavala, G.A. (1994). Algunos factores ecológicos que determinan la distribución y abundancia del lobo marino Zalophus californianus, en el Golfo de California. Cien. Mar., 20(4): 535-553. [ Links ]
Aurioles-Gamboa, D. y García-Rodríguez, F. (1999). Fluctuaciones de la población de lobo marino y la pesquería de sardina en el Golfo de California. Resúmenes XXIV Reunión Internacional para el Estudio de los Mamíferos Marinos, Mazatlán, Sinaloa, Ab. pp. 18-22. [ Links ]
Aurioles-Gamboa, D., Sinsel, F., Fox, C., Alvarado, E. and Maravilla, O. (1983). Winter migration of subadult male California sea lions (Zalophus californianus) in the southern part of Baja California. J. Mammal., 64 (3): 513-518. [ Links ]
Aurioles-Gamboa, D., Fox, C., Sinsel, F. and Tanos, G. (1984). Prey of the California sea lion (Zalophus californianus) in the Bay of La Paz, Baja California Sur, Mexico. J. Mammal., 65(3): 519-521. [ Links ]
Aurioles-Gamboa, D., Le Boeuf, B.J. y Findley, L.T. (1993). Registros de pinnípedos poco comunes para el Golfo de California. Rev. Inv. Cient., Univ. Aut. Baja California Sur (No. Especial de la SOMMEMA), 1: 13-19. [ Links ]
Bigg, A.M., Ellis, M.G., Cottrell, P. and Milette, L. (1990). Predation by harbour seals and sea lions on adult salmon in Comox Harbour and Cowichan Bay, British Columbia. Can. Tech. Rep. Fish. Aquat. Sci., 1769, 31 pp. [ Links ]
Castro-González, M.I., Aurioles-Gamboa, D., Montaño, S.B., Pérez-Gil, R.F. y López, N.O. (2001). Lípidos totales, colesterol y triglicéridos plasmáticos en crías del lobo marino de California (Zalophus californianus) del Golfo de California. Cien. Mar., 27(3): 375-396. [ Links ]
Costa, D.P. (1987). Isotopic methods for quantifying material and energy intake of free-ranging marine mammals. In: A.C. Huntley, D.P. Costa, G.A. Worthy and M.A. Castellini (eds.), Approaches to Marine Mammal Energetics. Allen Press. 210 pp. [ Links ]
Costa, D.P. (1991). Reproductive and foraging energetics of pinnipeds: Implications for life history patterns. In: D. Renouf (ed.), Behavior of Pinnipeds. Chapman and Hall, London, pp. 300-344. [ Links ]
Costa, P.D. (1993). The relationship between reproductive and foraging energetics and the evolution of the Pinnipedia. In: I.L. Boyd (ed.), Marine Mammals: Advances in Behavioural and Population Biology. Zoological Society of London, Monogr. 66, Clarendon Press, Oxford, pp. 293-314. [ Links ]
Fiscus, C.H. (1979). Interactions of marine mammals and Pacific hake. Mar. Fish. Rev., October: 1-9.
Fiscus, C.H. (1980). Marine mammal-salmonid interactions: A review. In: J.W. McNeil and C.D. Himsworth (eds.), Salmonid Ecosystems of the North Pacific. Oregon State Univ. Press, Corvallis, pp. 121-132. [ Links ]
Fitch, J.E. (1964). The fish fauna of the Playa del Rey locality, a southern California marine Pleistocene deposit. Los Angeles County Mus., Contrib. in Science, 82: 1-35. [ Links ]
Fitch, J.E. (1966). Additional fish remains, mostly otoliths, from a Pleistocene deposit at Playa del Rey, California. Los Angeles County Mus., Contrib. in Science, 119: 1-16. [ Links ]
Fitch, J.E. (1968). Otoliths and other fish remains from the Timms Point Silt (Early Pleistocene) at San Pedro, California. Los Angeles County Mus., Contrib. in Science, 146: 1-29. [ Links ]
Fitch, J.E. and Brownell, R.L. (1968). Fish otoliths in Cetacean stomachs and their importance in interpreting feeding habits. J. Fish. Res. Board Can., 25(12): 2561-2574. [ Links ]
Fraker, M.A. and Mate, R.B. (1999). Seals, sea lions, and salmon in the Pacific Northwest. In: J.R. Twiss and R.R. Reeves. (eds.), Conservation and Management of Marine Mammals. Smithsonian Inst. Press, Washington, DC, pp. 99-119. [ Links ]
García-Rodríguez, F.J. (1995). Ecología alimentaria del lobo marino de California, Zalophus californianus californianus, en Los Islotes, BCS, México. Tesis profesional, Universidad Autónoma de Baja California Sur, La Paz, 106 pp. [ Links ]
Harcourt, R., Aurioles-Gamboa, D. and Sánchez, J. (1994). Entanglement of California sea lions at Los Islotes, Baja California Sur, Mexico. Mar. Mammal Sci., 10(1): 122-125. [ Links ]
Heemstra, P.C. (1995). Serranidae. In: W. Fischer, F. Krupp, W. Schneider, C. Sommer, K.E. Carpenter and V.H. Niem (eds.), Guia FAO para Identificación de Especies para los Fines de la Pesca. Pacifico Centro-Oriental. Vol. III. Vertebrados: Parte 2. FAO, Roma, pp. 1565-1613. [ Links ]
Innes, S., Lavigne, M.D., Earle, M.W. and Kovacs, M.K. (1987). Feeding rates of seals and whales. J. Animal Ecol., 56: 115-130. [ Links ]
Jones, R.E. (1981). Food habits of smaller marine mammals from northern California. Proc. Calif. Acad. Sci., 42(16): 409-433. [ Links ]
King, J.E. (1983). Seals of the World. Comstock Publishing Assoc., Ithaca, New York, 240 pp. [ Links ]
Le Boeuf, B.J., Aurioles-Gamboa, D., Condit, R., Fox, C., Gisiner, R., Romero, R. and Sinsel, F. (1983). Size and distribution of the California sea lion population in Mexico. Proc. Calif. Acad. Sci., 43(7): 77-85. [ Links ]
Lluch, D.B. (1969). El lobo marino de California Zalophus californianus californianus (Lesson, 1828) Allen, 1880. Observaciones sobre su ecología y explotación. Inst. Mexicano de Recursos Naturales Renovables, México, DF, 69 pp. [ Links ]
Northridge, S.P. (1985). Estudio mundial de las interacciones entre los mamíferos marinos y la pesca. FAO, Documento Técnico de Pesca, No. 251, 190 pp. [ Links ]
Northridge, S.P. and Hofman, J.R. (1999). Marine mammal interactions with fisheries. In: J.R. Twiss and R.R. Reeves (eds.), Conservation and Management of Marine Mammals. Smithsonian Inst. Press, Washington, DC, pp. 99-119. [ Links ]
Odell, D.K. (1975). Breeding biology of the California sea lion, Zalophus californianus. Rapp. P.-V. Reun. Con. Int. Explor. Mer, 169: 374-378. [ Links ]
Orr, A.J. (1998). Foraging characteristics and activity patterns of the California sea lion (Zalophus californianus californianus) in the Bay of La Paz, Baja California Sur. M.Sc. thesis, California State University at Fresno, 116 pp. [ Links ]
Orr, R.T., Shonewald, J. and Kenyon, W.K. (1970). The California sea lion: Skull comparison of two populations. Proc. Calif. Acad. Sci., 37: 381-394. [ Links ]
Perez, M.A., McAlister, B.W. and Mooney, E.E. (1990). Estimated feeding rate relationship for marine mammals based on captive animal data. NOAA Tech. Memorandum NMFS F/NWC 184, 30 pp. [ Links ]
Peterson, R.S. and Bartholomew, A.G. (1967). The Natural History and Behavior of the California Sea Lion. Spec. Publ. No. 1, American Soc. of Mammalogists, 79 pp. [ Links ]
Ramírez-Rodríguez, M. (1991). Redes agalleras en la Bahía de La Paz, BCS. Rev. Inv. Cient., Univ. Aut. de Baja California Sur, 2(2): 65-74. [ Links ]
Ramírez-Rodríguez, M. (1997). Producción pesquera en la Bahía de La Paz, BCS. En: J. Urbán-Ramírez y M. Ramírez-Rodríguez (eds.), La Bahía de La Paz, Investigación y Conservación. Univ. Aut. de Baja California Sur, La Paz, México, pp. 273-281. [ Links ]
Scheffer, V.B. (1958). Seals, Sea Lions and Walruses. Stanford Univ. Press, California, 179 pp. [ Links ]
SEMARNAT: Secretaría de Medio Ambiente y Recursos Naturales (2000). Programa de Manejo Complejo Insular del Espíritu Santo, México. Comisión Nacional de Áreas Naturales Protegidas, México, DF, 194 pp. [ Links ]
Stewart, B. and Yochem, P. (1987). Entanglement of pinnipeds in synthetic debris and fishing nets and line fragments at San Nicolas and San Miguel Islands, California. Mar. Pollut. Bull., 18: 336-339. [ Links ]
Wyllie-Echeverria, T. (1987). Relationship of otolith length to total length in rockfishes from northern and central California. Fish. Bull., 85: 383-387. [ Links ]
Zavala, G.A. and Mellink, E. (1997). Entanglement of California sea lions, Zalophus californianus californianus, in fishing gear in the central-northern part of the Gulf of California. Fish. Bull., 95: 180-184. [ Links ]

