










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCiencias marinas
versión impresa ISSN 0185-3880
Cienc. mar vol.29 no.1 Ensenada mar. 2003
Artículos
Composición por tallas y edades de las poblaciones de abulón Haliotis fulgens y H. corrugata de la zona de Bahía Tortugas, Baja California Sur, México
Size and age composition of the populations of abalone Haliotis fulgens and H. corrugata of Bahía Tortugas, Baja California Sur, Mexico
María Georgina Gluyas-Millán1* y Jesús Talavera-Maya1
1 Centro Regional de Investigación Pesquera La Paz Instituto Nacional de la Pesca-SAGARPA Carretera a Pichilingue km. 1 s/n La Paz, CP 23020, Baja California Sur, México. E-mail: gmillan@balandra.uabcs.mx
Recibido en agosto de 2002;
aceptado en diciembre de 2002.
Resumen
Se estimó la biomasa del abulón amarillo Haliotis corrugata y abulón azul H. fulgens, y se determinó su composición por tallas y edades para la zona de Bahía Tortugas, en la costa occidental de la Península de Baja California en octubre de 1997. Se usó un muestreo al azar estratificado. Por medio de buceo semiautónomo tipo Hooka se colectaron abulones entre 1 y 18 m de profundidad con una unidad muestral de 10 m2. Los abulones se midieron y se tomó una submuestra para medir longitud y peso, determinar el sexo y el estadio de madurez, y colectar las conchas para determinar la edad en los anillos de la espira. Los promedios de la talla y la edad de la población accesible de H. corrugata fueron 135.8 mm y 6.6 años, y los intervalos de tallas y edades encontrados fueron de 55 a 180 mm en individuos de 2+ a 16+ años. Para H. fulgens los promedios fueron 154.1 mm y 5 años, con un intervalo de tallas de 57 a 206 mm en individuos de 1+ a 20+ años de edad. Se discuten las posibles causas de la diferencia en reclutamiento entre el abulón azul y el abulón amarillo, la disminución de grupos de edad respecto a años anteriores, y el adelanto del desove de H. corrugata a la luz del estatus del recurso y de las condiciones ambientales que prevalecieron durante de El Niño 1997-98.
Palabras clave: abulón, Haliotis fulgens, H. corrugata, tallas, edad, Bahía Tortugas, Baja California Sur.
Abstract
The size and age composition was determined for pink abalone Haliotis corrugata and green abalone H. fulgens populations of Bahía Tortugas in the western coast of the Baja California Peninsula, Mexico, in October 1997. The biomass was estimated by means of a stratified random design using semiautonomous Hooka diving. The abalones were collected from 1 to 18 m of depth with a 10-m2 sampling unit. All shell lengths were measured and a subsample was taken to measure shell length, to weigh, to indentify sex and maturity stage and to collect the shells to determine the age in the rings of the spire. The size interval of the population accessible of H. corrugata was 55 to 180 mm for individuals 2+ to 16+ years old, with a mean length of 135.8 mm and mean age of 6.6 years. For H. fulgens, organism sizes ranged from 57 to 206 mm for individuals 1+ to 20+ years old, with a mean length of 154 mm and mean age of 5 years. Possible causes are discussed regading the difference in recruitment between H. fulgens and H. corrugata, the decrease of the oldest year class in both species, and the earlier spawning of H. corrugata to the light of the status of the resource and of the environmental conditions that prevailed during El Niño 1997-98.
Key words: abalone, Haliotis fulgens, H. corrugata, age, size, Bahía Tortugas, Baja California Sur.
Introducción
El abulón es un molusco gasterópodo de la familia Haliotidae que habita zonas rocosas asociado a mantos de Macrocystis pyrifera y otras algas. Este recurso ha sostenido una pesquería por más de 100 años en la costa occidental la Península de Baja California. Fue aprovechado por extranjeros desde finales del siglo XIX hasta los años cuarenta, y por pescadores mexicanos en los últimos 50 años (Ponce-Díaz et al., 1998). Es un recurso de alto valor y demanda en al mercado internacional que ocupa el cuarto lugar en generación de divisas, después del camarón, el atún y la langosta (SEMARNAP, 1996). Las dos especies que soportan la pesquería son el abulón azul, Haliotis fulgens, y el abulón amarillo, H. corrugata (Muciño Díaz et al., 2000). No obstante los esfuerzos de investigación y las medidas de manejo como talla mínima de captura, veda y cuotas de captura (Diario Oficial de la Federación, 1981; 1987; 1993a; 1993b; 1999), la producción ha disminuido de 6000 t en 1950 (León y Muciño, 1996) a 4671 en 1997 (SEMARNAP, 1997). La Carta Nacional Pesquera (CNP) señala el estatus del abulón como un recurso deteriorado, sus poblaciones se encuentran a un nivel abajo del óptimo y bajo una estrategia de recuperación (Diario Oficial de la Federación, 2000) que consiste en incrementar el tamaño de la población, no permitir que disminuya y llevarla al nivel donde se obtenga el mayor rendimiento (Muciño Díaz et al., 2000). Como parte de esa estrategia, el Instituto Nacional de La Pesca (INP) recomendó orientar la investigación al análisis de la dinámica de la población con un modelo estructurado por edades, además del modelo de producción excedente que actualmente se usa. La información básica necesaria para ello es conocer la composición de la biomasa, esto es, el número de organismos por tamaños y edades que la componen. Esta información la proporciona la estructura por tallas y edades que es necesaria para reconstruir la historia de la población. Los cambios en el tiempo de esa estructura reflejarían: (1) los efectos del reemplazamiento, el crecimiento y la mortalidad; (2) la variación del vigor de las clases anuales; y (3) los cambios del promedio de edad reflejarían el incremento o disminución del reclutamiento por efecto de clases anuales vigorosas o débiles, o por la pesca entre otros. La estructura de tallas y edades es específica de cada población, refleja la relación de la especie con el ambiente y los efectos de la extracción por pesca. Una especie existe bajo condiciones continuamente cambiantes, la estructura refleja tales condiciones con ciertos límites y, por consiguiente, representa las adaptaciones de las poblaciones a esas condiciones (Nikolskii, 1969).
El presente trabajo tiene por objeto, determinar la estructura de tallas y edades de las poblaciones de abulón H. fulgens y H. corrugata de la zona de Bahía Tortugas por medio de la determinación de la edad de manera directa. Los resultados se discuten a la luz del estado del recurso y las condiciones ambientales durante el evento El Niño 1997-98. La composición por edades de las poblaciones de abulón se considera una información representativa y única en su tipo a la fecha, ya que para su obtención se han seguido los procedimientos recomendados para ser aplicados a la evaluación de poblaciones (Ricker, 1975; Lin Lai et al.,1996).
Materiales y métodos
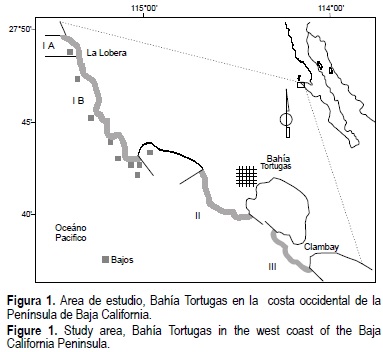
Se llevó a cabo una evaluación cuantitativa y cualitativa de abulón entre La Lobera y el punto medio del playón de Clam Bay (fig. 1), en la costa noroccidental de Baja California Sur (BCS), del 18 al 29 octubre de 1997. Se utilizó la metodología estándar implementada por el INP para evaluar el recurso (Muciño-Díaz et al., 2000) que consiste en un muestreo al azar estratificado (Cochran, 1978; Anonymous, 1998). Se determinaron cinco estratos (IA, IB, II, III y Bajos), de acuerdo a la distribución horizontal del recurso que está en función de la presencia de sustrato rocoso. De esta manera los estratos quedaron definidos naturalmente, aislados por zonas de arena que mantienen una menor diversidad biológica que aquellas donde se distribuye el abulón, y que se caracterizan por la presencia de grandes laminarias (Macrocystis piryfera y Eisenia), algas coralinas calcáreas articuladas y costrosas, y densos pastos de Phyllospadix torreyii en la zona de rompiente (Guzmán del Proo et al., 1991). Se midieron las dimensiones de los estratos considerando el hábitat efectivo para el abulón, descartando el área de arena. Se definieron bloques de 500 m de largo, paralelos a la costa, por la correspondiente distancia del ancho hasta donde se presenta el sustrato rocoso o a la ísobata de 18 m. En cada bloque se realizaron de 8 a 10 inmersiones por medio de buceo semiautónomo tipo Hooka, en cada inmersión se colectaron los abulones presentes por unidad muestral de 10 m2, se midieron y se regresaron al sitio original con el mínimo disturbio (tabla 1). Con esta información se determinó la composición por tallas. Se tomó una submuestra seleccionando al azar hasta cinco individuos por intervalo de talla, se midió su longitud, el peso del callo, se determinó el sexo y la madurez de la gónada, y se colectó la concha para determinar edad. La madurez gonádica se determinó con una escala morfocromática (García-Juárez y Ortiz-Quintanilla, 1992; Reinecke-Rodríguez, 1997) con los siguientes estadios: (I) Incipiente o Inmaduro, (II) Madurando, (III) Maduro, (IV) Desove o desovando, y (V) Desovado o en reposo. El sexo se determinó con el color de las gónadas, en los machos color crema y blanco y, en las hembras, verde olivo y café oscuro (Belmar y Guzmán del Proo, 1992).

La biomasa de la población se estimó utilizando el muestreo estratificado aleatorio (Cochran, 1978; Annónimo, 1998) de acuerdo a los siguientes estimadores:

Los límites de confianza (a = 0.05) de la biomasa se calcularon de acuerdo a:
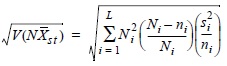
donde Ni es el tamaño del estrato en número de unidades muestrales de 10 m2, N es la suma del tamaño de todos los estratos en unidades muestrales, L es el número de estratos (IA, IB, II, III y Bajos), ni es el número de unidades muestrales o número de inmersiones efectuadas en cada estrato, Xi es la biomasa promedio por estrato para las i-ésimas observaciones de ni, y si2 es la varianza por estrato para las i-ésimas observaciones de ni.
La edad se determinó en la espira de la concha del abulón, una vez desbastada con un pulidor eléctrico y posteriormente con lija de grano fino, hasta que los anillos quedaron descubiertos y apareció un orificio en una sección horizontal (Muñoz-López, 1976; Shepherd et al., 1995). En un microscopio estereoscópico se contaron los anillos expuestos desde el margen al orificio. El conteo se hizo tres veces por dos lectores, hasta llegar a un acuerdo en el número de anillos observados. Para la identificación de los anillos en H. fulgens se siguieron las recomendaciones de Shepherd y Turrubiates-Morales (1997), quienes validaron la periodicidad anual de formación de los anillos obscuros hasta los 5 años, y para H. corrugata se siguieron las recomendaciones de Shepherd y Ávalos-Borja (1997), quienes validaron los anillos de pigmentación oscura hasta los 12 años. En este trabajo se asumió que después del sexto anillo pigmentado en H. fulgens y del décimo tercer anillo en H. corrugata, éstos continúan teniendo una periodicidad anual en su formación. Esta suposición es común después de validar la periodicidad de formación de una o dos marca(s) de crecimiento en especies de peces o moluscos, principalmente en las longevas. Sin embargo, nosotros apoyamos esta suposición en que: (1) tanto para el abulón azul como para el amarillo, se siguieron los mismos criterios de lectura de Shepherd y Turrubiates-Morales (1997), y Shepherd y Ávalos-Borja (1997); (2) se mantuvo consistencia en tales criterios; (3) en la zona de estudio se mantiene una marcada estacionalidad del ambiente físico (Guzmán del Proo et al., 1991; Turrubiates-Morales y Castro-Ortiz, 1992), especialmente de la temperatura del mar, presumiblemente uno de los factores que promueven la formación de la marca anual y que influye en la reproducción, la cual continua ocurriendo en H. fulgens > 5 años y en H. corrugata > 12 años. La composición por edades de cada especie se determinó a partir de una clave edad-longitud siguiendo las recomendaciones de Ricker (1975) y Lin Lai et al. (1996) para su aplicación en la evaluación de poblaciones.
Resultados
La composición por tallas y edades se determinó para una área de 6,146,745 m2 en la franja litoral entre 1 y 18 m de profundidad. Se definieron 54 bloques en los cuales se realizaron 504 inmersiones, se obtuvo un total de 1549 abulones distribuidos en cuatro estratos (tabla 1), y se definió el porcentaje de cada estrato respecto al área total. El 31.6% correspondió a H. corrugata y 68.4% a H. fulgens. En la franja de 1 a 7 m se realizaron el 53% de las inmersiones, en la de 7.1 a 14 m el 35.2%, y en la de 14.1 a 18 m el 11.7%. El tipo de sustrato o hábitat efectivo del abulón fue roca alta, roca plana de grandes extensiones (de 15 a 40 m), grava, y roca con cuevas.
La biomasa total estimada en peso de callo fresco fue mayor para H. fulgens 337,653 con un intervalo de confianza de ± 44,712 kg, y menor para H. corrugata 79,123.5 ± 11,792, que representan 81% y 19% de la biomasa total, respectivamente. Los estratos más productivos de abulón en términos de biomasa fue el estrato III y II (fig. 2).
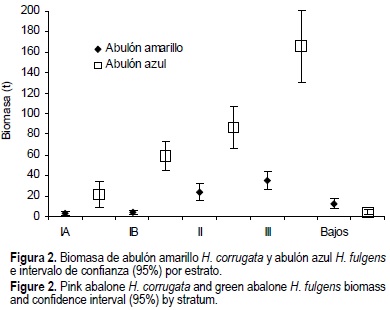
La composición por tallas de H. corrugata presentó un intervalo de 55 a 180 mm y una talla media de 135.8 mm (fig. 3). Para las tallas de H. fulgens el intervalo fue de 57 a 206 mm y la talla media de 154.1 (fig. 4). Si definimos el reclutamiento como la fracción de la población accesible, a partir de la talla mínima de captura (TMC) hasta la talla mínima registrada en la presente evaluación, para H. fulgens éste correspondería a individuos de 145 a 57 mm, y en H. corrugata a individuos de 135 a 55 mm. A partir de esta definición, el reclutamiento promedio de H. fulgens fue 27% y el de H. corrugata 48% (figs. 3 y 4; tabla 2), no obstante la diferencia del tamaño de la biomasa (fig. 2). Excepcionalmente, la zona II presenta un reclutamiento de 70% para H. fulgens (tabla 2).
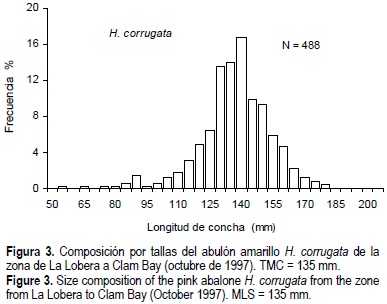
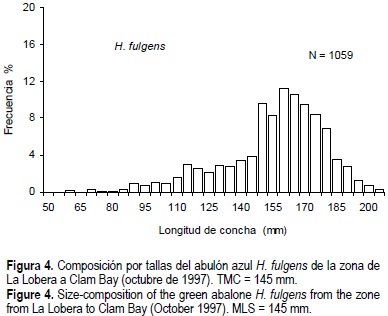

Se determinó la edad de manera directa en 105 conchas de H. corrugata y 192 de H. fulgens, se contaron las marcas y se obtuvo la edad cronológica. El intervalo de edades para H. corrugata fue de 2+ a 16+ años y para H. fulgens fue de 1+ a 20+ años. El 10% de las conchas registraron perforaciones superficiales de parásitos gasterópodos, en organismos con edades a partir de los 4+ años, siendo en su mayoría individuos de > 7 años. Estos organismos se encontraron aparentemente en buenas condiciones.
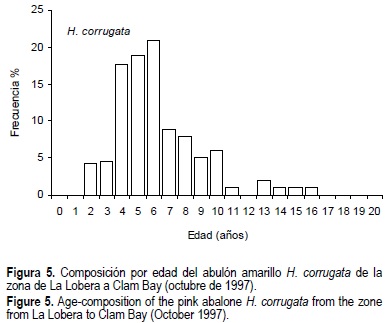
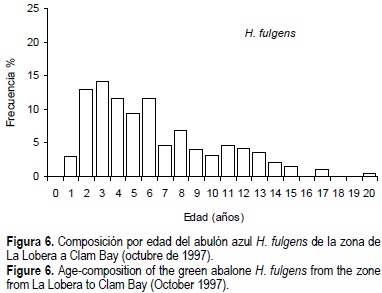
Más del 50% de la población de H. corrugata estuvo representada por individuos de 2+ a 6+ años de edad (fig. 5), el resto fueron individuos de 7+ a 16+ años. La edad media fue 6.6 años. Las edades más abundantes fueron 6+, 5+ y 4+ años, que corresponden a las clases anuales 1991, 1992 y 1993, respectivamente. En H. fulgens, el 51% de la población fueron individuos de 1+ a 5+ años, la edad media fue de 5 años y sobresalieron las edades de 3+, 2+, 4+ y 6+ años que corresponden a las clases anuales 1994, 1995, 1993 y 1991 respectivamente (fig. 6).
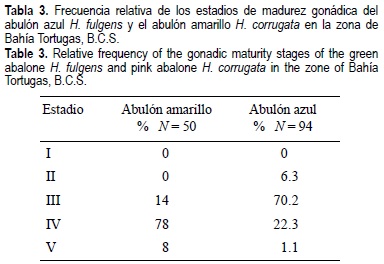
El 78% de H. corrugata > 110 mm estaba desovando (estadio IV), mientras que el 70% de H. fulgens > 115 mm se encontró maduro (estadio III) próximo a desovar, y el 30% estaba desovando (estadio IV) (tabla 3). La proporción de machos por hembra fue de 1:1.7 en H. corrugata y de 1:1.4 en H. fulgens. El desove coincidió con un descenso de la temperatura superficial del mar de 25 a 23°C durante el periodo de estudio.
Durante las inmersiones se observó un hábitat totalmente descubierto del alga M. pyrifera, quedando la población completamente expuesta a la vista del buzo. El sustrato rocoso donde se adhiere el abulón, y donde ocurre el asentamiento de sus larvas, se observó con escasa flora que normalmente cubre las rocas en años de condiciones climáticas normales (en que no se manifiesta en fenómeno de El Niño). Los abulones mostraron condiciones aparentemente sanas, con escaso alimento en el estómago, el cual consistió en algas coralinas calcáreas articuladas y costrosas, y partículas de sedimento.
Discusión
La naturaleza sedentaria del abulón permite realizar evaluaciones directas que pueden utilizarse como un método que provee estimaciones independientes de la pesquería. Esta metodología ha sido utilizada por el INP desde principios de los años noventa. En esta ocasión la evaluación se realizó en condiciones en que las poblaciones de abulón estuvieron completamente expuestas y accesibles, debido a la ausencia de M. pyrifera, lo que permitió obtener un amplio intervalo de tallas y edades de H. corrugata y H. fulgens. Consideramos que esta información es adecuada, ya que representa una extensa área, con una amplia variedad de hábitats, y a pesar de que se generó de un periodo corto.
La utilización de marcas de crecimiento, previa validación de su periodicidad anual de formación, hace que el método utilizado en el presente trabajo sea potencialmente útil para las necesidades de investigación actuales. La edad mínima encontrada para H. corrugata fue 2 + años en individuos de 55 mm y de 1+ años para H. fulgens, en individuos de 57 mm. Aunque el propósito de este trabajo no es determinar el crecimiento individual del abulón, los antecedentes señalan una gran variabilidad en el crecimiento individual entre zonas y, dentro de las zonas, variación estacional en el crecimiento y, por consiguiente, una amplia distribución de longitudes alrededor del promedio de longitud por edad, especialmente en los primeros años (Shepherd et al., 1991; Turrubiates-Morales y Castro-Ortiz 1992; Day et al., 1995; Wells y Mulvay, 1995; Worthington et al., 1995; Troynikov et al., 1998). Los resultados siguientes ilustran lo anterior: para H. corrugata de 2+ años, Marín (1981) estima una talla media de 83 mm; Shepherd y Ávalos Borja (1997), con la misma técnica del presente trabajo, reportan una talla de 50 mm para individuos de 2+ años semejante a la de nuestros resultados. Haliotis fulgens en Bahía Tortugas logra alcanzar una talla media de 45 mm a la edad 0+ años (Turrubiates-Morales, com. pers., INP-SAGARPA). Shepherd et al. (1991) reportan 70 mm en individuos de 2+ años. Shepherd et al. (1998) encuentran en Isla Natividad, BCS, individuos de 40 a 110 mm, con edades de 2+ y 3+ años. Guzmán del Proo et al. (2001) estima un crecimiento medio de 55.4 ± 5.1 mm/año en abulones silvestres de 25-70 mm, de tal manera que individuos > 1+ y < 2+ años medirían aproximadamente 100 mm. Nosotros encontramos en H. fulgens individuos de 1+ años de 57 mm, lo cual es consistente con algunos antecedentes considerando la amplia variación de tallas por edad.
Composición por tallas y edades
En especies longevas como el abulón Haliotis spp., las determinaciones directas de edad tienen algunas ventajas respecto a los métodos de distribuciones de frecuencia de tallas: se distinguen los grupos de edad mas viejos; se identifica el tiempo de vida del primer grupo de edad y se distinguen los organismos de lento y rápido crecimiento, entre y dentro de zonas, entre otros.
Los resultados indican que la composición de tallas está cargada hacia la derecha donde predominan varias clases anuales acumuladas, y pocos organismos pequeños y jóvenes (figs. 3, 4, 5 y 6), tal como se observa en otras especies de abulón (Wells y Keesing, 1990; Tegner et al., 1992; Day y Fleming, 1992; Wells y Mulvay, 1995). La baja frecuencia de abulones chicos sugiere su hábitat críptico (McShane, 1992; Prince, 1992). El abulón empieza a salir de su hábitat críptico cuando alcanza la madurez (Prince, 1992). Posiblemente el método utilizado no sea el más adecuado para evaluar esa fracción de la población, si consideramos la complejidad del hábitat natural del molusco (McShane, 1992). Otro escenario sería que la abundancia de juveniles fuera realmente baja, pero que tuvieran una alta tasa de crecimiento que les permita rápidamente alcanzar la talla adulta y sobrevivir varios años como adultos (Wells y Keesing, 1997), o una combinación de los anteriores. Tal situación aún no es completamente clara. Una revisión sobre reclutamiento de varias especies de abulón concluye que las diferencias en el reclutamiento pueden ser grandes (McShane, 1995). A partir de lo anterior y considerando el método de evaluación, se reconsideró la definición de reclutamiento, que sería la fracción de la población accesible, entre 145 mm (TMC) y 80 mm. Los individuos < 80 mm no están completamente accesibles por su condición críptica y representan < 0.8% de la población utilizando la metodología aquí descrita. Resultados similares se han observado en distintas áreas de distribución del abulón en la costa occidental de BCS desde finales de los años noventa (Muciño Díaz et al., 2000; Turrubiates-Morales com. pers), y es una situación común en otras poblaciones de abulón del mundo (Prince y Guzmán del Proo, 1993).
Después de esta consideración, la diferencia en la proporción de reclutas de H. fulgens (27%) respecto a H. corrugata (48%) (figs. 3 y 4) se intenta explicar de la siguiente manera: (1) si consideramos que la distribución de H. fulgens es desde la zona entre mareas hasta aproximadamente 24 m de profundidad, una zona de alta energía de oleaje y corrientes donde los individuos expuestos serán los más fuertes para resistir esas condiciones, los menos fuertes tenderían a ocultarse. Sin embargo, durante El Niño 1997-98 la temperatura superficial del mar aumentó 3°C o más por al menos seis meses, y desaparecieron los mantos de M. piryfera (Ladha et al., 1999), una de las principales fuentes de alimento y protección del abulón (Guzmán del Proo et al., 1991). Esta situación podría provocar una disminución en la condición del molusco que se reflejaría en adoptar un comportamiento críptico para protegerse; (2) sin embargo, la ausencia de M. pyrifera promovió que la población de abulón estuviera completamente expuesta y en busca de alimento, encontrándose en su tracto digestivo algas coralinas y costrosas, y gran cantidad de partículas de sedimento, que ran los únicos alimentos disponibles entonces, reduciéndose así la posibilidad de una subestimación de la proporción de los reclutas aquí definidos; (3) por otro lado, la remoción histórica de agregaciones de reproductores afectaría la magnitud del reclutamiento y, por lo tanto, la composición por tallas y edades; (4) además, el reclutamiento a la pesquería puede depender de un exitoso asentamiento de larvas en un sustrato apropiado (McShane, 1995), sin embargo, aún hay incertidumbre en la relación entre el suministro de larvas y el reclutamiento del abulón (McShane 1995), indicando la necesidad de mayor información sobre las estrategias de la historia de vida de H. fulgens y H. corrugata. Finalmente, consideramos que la reducida proporción de reclutas en H. fulgens sería una evidencia del estado en deterioro del recurso, específicamente, de una sobrepesca del reclutamiento. Esta situación cambiará conforme se cumpla con la estrategia de recuperación del recurso.
Para H. corrugata la población estuvo completamente expuesta en busca de alimento (fig. 3) igual que H. fulgens (fig. 4), por lo que también es poco probable una subestimación de la proporción de reclutas. Sin embargo, H. corrugata se distribuye a mayor profundidad, aproximadamente a 44 m, donde la accesibilidad por buceo semiautónomo tipo Hooka tiene riesgos. En esas profundidades podrían encontrarse los individuos más viejos que estarían aportando reclutas a la población, lo cual podría ser el origen de la mayor proporción de reclutas, y confirma la evidencia de una ligera recuperación de H. corrugata señalada por Muciño-Díaz et al. (2000). Las profundidades > 24 m se consideran un refugio para H. corrugata, por su baja accesibilidad. El refugio puede jugar un rol importante en la estrategia de conservación y recuperación del recurso, manteniendo una producción de huevos y diversidad genética (Shepherd y Brown, 1993).
Lo anterior explica también por qué encontramos menos de 5% de individuos > 11 años y una edad máxima de 16 años. Otra explicación sería una disminución real de los grupos de edad más viejos y, tratándose de zonas de pesca históricas tradicionales, esta disminución es una evidencia del deterioro de la población. Nosotros encontramos en 1997 una edad máxima de 16 años para H. corrugata, mientras que Muñoz López (1976) reportó individuos de hasta 21 años antes de 1976, y Shepherd y Ávalos-Borja (1997) individuos de 18 años en 1970 para Isla Natividad, BCS, México, ambos usando la técnica de los anillos de la espira. Marín (1981), usando distribuciones de frecuencia de tallas, reporta un grupo de edad de 20 años.
Una situación similar ocurre con H. fulgens. En este trabajo se encontró una edad máxima de 20 años, que es una cantidad menor de grupos de edad respecto a 1982 y 1985 cuando Turrubiates-Morales y Castro-Ortiz (1992) reportan, para la misma zona y usando la técnica del presente trabajo, individuos de hasta 25 años. Esta situación podría ser una evidencia de sobrepesca del reclutamiento, que sería la disminución de adultos de una manera tal que no se producen suficientes huevecillos para remplazar el reclutamiento. La pesca retira los organismos más viejos, que es una de las principales características de las poblaciones explotadas a niveles de intensidad importantes. Al inicio de la pesquería, cuando la producción alcanzó cerca de 5000 t, H. corrugata predominaba en las capturas, después, a partir de los años ochenta H. fulgens empezó a predominar pero sin alcanzar la producción de H. corrugata (León y Muciño, 1996).
Desove
Si consideramos que de 1990 a 1995 el máximo del desove de H. fulgens se registró en diciembre-enero, y el de H. corrugata de octubre a diciembre (Muciño-Díaz et al., 1996), la presencia de más de 70% de H. corrugata en estadio de desove y 70% de H. fulgens a punto de desovar en octubre de 1997, indicó un adelanto del desove en H. corrugata . Una situación similar se observó durante los eventos El Niño de 1982-83 y 1991-92 en la misma zona (Espinoza-Castro y Turrubiates-Morales, com. pers., INP-SAGARPA). Desde finales del verano de 1997 hasta el verano de 1998, las aguas costeras de California y la Península de Baja California estuvieron influenciadas fuertemente por condiciones El Niño (Lynn et al., 1998), con un amplio periodo de calentamiento del mar (Ladah et al., 1999), situación que, aunada a un descenso brusco de la temperatura superficial del mar de 25 a 23°C en octubre 1997, pudo promover el adelanto del desove. No obstante, el evento reproductivo está protegido con un periodo de veda del 1 de agosto al 31 de diciembre (Diario Oficial de la Federación, 1981).
Las siguientes situaciones observadas y documentadas para la zona de estudio podrían afectar el vigor de la cohorte de abulón de 1997: (1) un amplio periodo de calentamiento del mar debido a El Niño 1997-98 (Lynn et al., 1998) adelantó el desove de H. corrugata, evento que está en función del incremento de la temperatura del mar (Cox 1962), encontrando así condiciones ambientales distintas a las de años normales que pudieran no favorecer la sobrevivencia; (2) un cambio de los elementos que componen la dieta del abulón sugiere un cambio de su contenido energético y valor nutritivo (lo que afectaría la calidad de los gametos), ya que durante el Niño 1997-98 se observó en el tracto digestivo predominancia de las algas coralinas articuladas Bossiella y Corallina spp. y partículas de sedimento. Estas algas no se reportan en la dieta de H. fulgens en años en los que no se presenta El Niño (Servier et al. , 1998), no obstante que forman parte del hábitat de Haliotis spp. (Guzmán del Proo et al., 1991). No obstante, algunas observaciones en años No Niño señalan escasa presencia de esas algas en el tracto digestivo de H. fulgens de Bahía Tortugas (Turrubiates Morales, com. pers); (3) la ausencia de M. pyrifera afectaría el proceso de asentamiento de la larva y por consiguiente la sobrevivencia, ya que sus bosques sirven de protección y constituyen una barrera que disminuye la velocidad de la corriente y evita el transporte de larvas fuera de las zonas de producción o arrecifes (Guzmán del Proo et al., 2000); (4) un cambio en la composición cuantitativa y cualitativa de la flora del sustrato, por las condiciones ambientales que prevalecieron durante El Niño 1997-98, afectaría la naturaleza del sustrato de asentamiento. Esta situación significaría ausencia de alimento adecuado durante el asentamiento de la larva, afectando el crecimiento individual, la sobrevivencia y, por consiguiente, el reclutamiento. No obstante que aún no se define la etapa de vida o periodo crítico en el cual se determina el reclutamiento en el abulón, el inicio de la primera alimentación exógena tiene relación con la etapa de asentamiento de la larva; para el caso del abulón éste podría ser el periodo crítico señalado por Hjort (1926) y citado por Ricker (1975).
El conocimiento de la composición por tallas y edades de las poblaciones de abulón en la costa occidental de la Península Baja California proporciona evidencias del estado del recurso, constituye un avance para su análisis cuantitativo, y aporta elementos biológicos que, junto con otra información de parámetros poblacionales y fecundidad por edad, permitirán definir nuevos puntos de referencia y probar otras alternativas para el análisis de la dinámica de la población.
Agradecimientos
A los socios, directivos y a Mario Andrade Patrón de la SCPP Bahía Tortugas, SCL, por su valioso apoyo en el trabajo logístico y de buceo; a Miguel Ángel Reinecke, Agustín del Valle y Ramón Ayala por su participación en el trabajo de campo; a Felipe Neri Melo y Armando Hernández por su apoyo en el laboratorio. Un agradecimiento especial a José Remedios Turrubiates, Gerónimo Espinoza, Margarita Muciño y Pedro Sierra del CRIP-La Paz-INP-SAGARPA por su disposición para comunicar sus observaciones y experiencias sobre la biología del abulón; y a dos revisores anónimos por sus observaciones, y crítica interesante y constructiva. El primer autor recibió apoyo de CONABIO para desarrollar el Proyecto de Investigación L-200 convenio FB469/L200/97. Este trabajo está dedicado a la memoria de Gilberto León Carballo, pionero de las evaluaciones de abulón en México.
Referencias
Anonymous (1998). Improving fish stock assessments. Committee on Fish Stock Assessment Methods, Ocean Studies Board, Commission on Geosciences, Environment and Resources, National Research Council. National Academy Press. Washington, D.C. 1-177. [ Links ]
Belmar-Pérez, J. y Guzmán del Proo, S. (1992). Madurez sexual y ciclo gonádico en Haliotis fulgens y Astraea undosa en Bahía Tortugas. Pag.121-130. En: Guzmán del Proo (ed.), Memorias del Taller México-Australia sobre reclutamiento de recursos bentónicos de Baja California. IPN-Secretaria de Pesca. 212 pp. [ Links ]
Cochran, W.G. (1984). Técnica de Muestreo. Edit. Continental S. A. de C. V. cuarta reimpresión 513 pp. [ Links ]
Cox, K.W. (1962). California abalones, family Haliotidae. Fish. Bull. California, 118, pp.1-129. [ Links ]
Day, R.W. and Fleming, A.E. (1992). The determinants and measurement of abalone growth. In: S.A. Shepherd, M.J. Teagner and S.A. Guzman del Proo (eds.), Abalone of the World: Biology, Fisheries and Culture. Blackwell, Oxford. pp.141-181. [ Links ]
Day, R.W., Williams, M.C. and Hawakes, G.P. (1995). A comparison of fluorochromes for marking abalone shells. Mar. Freshwater Res., 46: 599-605. [ Links ]
Diario Oficial de la Federación (1981), 21 de septiembre. Acuerdo mediante el cual se establecen medidas de regulación para la extracción de abulón. [ Links ]
Diario Oficial de la Federación (1987), 29 de junio. Acuerdo mediante el cual se establecen medidas de regulación para la extracción de abulón. [ Links ]
Diario Oficial de la Federación (1993a), 21 de septiembre Norma Oficial Mexicana-005 Pesca-1993. [ Links ]
Diario Oficial de la Federación (1993b), 31 de diciembre Norma Oficial Mexicana-009-Pesca-1993. [ Links ]
Diario Oficial de la Federación (1999), 28 de septiembre. Ley de Pesca y su Reglamento. [ Links ]
Diario Oficial de la Federación (2000), 28 de agosto. Anexo de la Carta Nacional Pesquera. [ Links ]
García-Juárez, A.M. y Ortiz-Quintanilla, M. (1992). Madurez sexual en abulón azul Haliotis fulgens (Mollusca:Gasterópoda) de Isla de Cedros, Baja California. Pag. 65-76. En: Guzmán del Proo (ed.), Memorias del Taller México-Australia sobre reclutamiento de recursos bentónicos de Baja California. IPN-Secretaria de Pesca. México, 212 pp. [ Links ]
Guzmán del Proo S., Mille-Pagaza, S.R., Guadarrama-Granados, R. de La Campa, S. Carrillo-Laguna, J., Pereira-Corona, A., Belmar-Pérez, J., Parra-Alcocer, M.J. y Luque-Guerrero, A.C. (1991). La comunidad bentónica de los bancos de abulón Haliotis spp. mollusca:gaterópoda) en Bahía Tortugas, Baja California Sur, México. An. Esc. Nac. Cienc. Biol. Méx., 36: 27-59. [ Links ]
Guzmán del Proo, S., Salinas, F., Zaytsev, O., Belmar-Pérez, J. and Carrillo-Laguna, J. (2000). Potential dispersion of reproductive products and larval stages of abalone Haliotis spp. As a function of the hydrodynamics of Bahia Tortugas, México. J. Shellfish Res., 19(2): 869-881. [ Links ]
Guzmán del Proo S., Belmar-Pérez, J., Carrillo-Laguna, J., Carreón-Palau, L., Herrera, R. and Castro, A. (2001). Growth of juvenile abalone in Bahia Tortugas, Baja California, Mexico. Abstract. CalCOFI Annual Conference 2001, 5-8 nov 2001, San Diego, Ca. U.S.A. [ Links ]
Ladah, L.B., Zertuche-González, J. and Hernández-Carmona, G. (1999). Giant kelp (Macrocystis pyrifera, Phaeophycea) recruitment near its southern limit in Baja California after mass disappearance during ENSO 1997-1998. J. Phycol., 35: 1106-1112. [ Links ]
León-Carballo G. y Muciño-Díaz, M. (1996). Pesquería de abulón. En: M. Casas-Valdez y G. Ponce Díaz (eds.), Estudio del Potencial Pesquero y Acuícola de Baja California Sur. Volumen I. pp. 15-41. [ Links ]
Lin Lai H., Gallucci, V. and Gunderson, D.R. (1996). Age determination in fisheries: Methods and applications to stock assessment. In: V. Gallucci, S. Saila, D. Gustafson and B. Rothschild (eds.), Stock Assessment, Quantitative Methods and Applications for Small-scale Fisheries. pp. 82-178. [ Links ]
Lynn, R.J., Baumgartner, T., Garcia, J., Collins,C.A., Hayward, T.L., Hyrebach, K.D., Mantyla, A.W., Murphree, T., Shankle, A., Schwing, F.B., Sakuma, K.M. and Teagner, M.J. (1998). The state of the California current, 1997-1998: transition to El Niño condition. CalCOFI Rep., 39: 25-49. [ Links ]
Marin, V.A. (1981). Parámetros poblacionales y diagnóstico de la pesquería de abulón amarillo Haliotis corrugata en Bahía Tortugas, B.C.S. Ciencia Pesquera, 1 (2): 67-80. [ Links ]
McShane, P.E. (1992). Early life of abalone: a review. In: S.A. Shepherd, M.J. Teagner and S.A. Guzmán del Proo (eds.), Abalone of the World: Biology, Fisheries and Culture. Blackwell, Oxford. pp. 120-140. [ Links ]
McShane, P.E. (1995). Recruitment variation in abalone: its importance to fisheries management. In: S.A. Shepherd, R.W. Day and A.J. Butler (eds.), Progress in Abalone Fisheries Research CSIRO. 1995, Australia. pp. 555-570. [ Links ]
Muciño-Díaz, M., León-Carballo, G., Vélez-Barajas, A., Turrubiates-Morales, J.R. y Reinecke-Rodríguez, M.A. (1996). Aspectos reproductivos del recurso abulón en las zonas II, III y IV y su relación con la aplicación del periodo de veda. En: Resúmenes II Simposium sobre investigación en Biología Pesquera y Oceanografía en México. 25-27 sep. 1996. [ Links ]
Muciño-Díaz, M., Sierra-Rodríguez, P., Vélez-Barajas, A., Zárate-Becerra, E., Talavera-Maya, J., Turrubiates-Morales, J.R., Caballero-Alegría, F. y Reinecke, M.A. (2000). Abulón. En: Díaz de León-Corral y M.A. Cisneros-Mata (eds.), Sustentabilidad y Pesca Responsable en México Evaluación y Manejo 1999-2000. INP-SEMARNAP, pp.119-157. [ Links ]
Muñoz-López, T. (1976). Resultados preliminares de un método para determinar la edad en abulones (Haliotis spp.) en Baja California. Mem. I Simp. Nal. Rec. Pesq. Mas. de México. Vol. Especial abulón/langosta SC/INP, pp.281-301. [ Links ]
Nikolskii, G. (1969). Theory of Fish Population Dynamics, as the Biological Background for Rational Exploitation and Management of Fishery Resources. Oliver & Boyd LTD. First English language edition 1969. 323 p. [ Links ]
Ponce-Díaz, G., Vega-Velázquez, A., Ramade-Villanueva, M., León-Carballo, G. y Franco-Santiago, R. (1998). Socioeconomic characteristics of the abalone fishery along the west coast of the Baja California Peninsula. J. Shellfish Res., 17(3): 853-857. [ Links ]
Prince, J.D. (1992). Using a spatial model to explore the dynamics of an exploited stock of the abalone Haliotis rubra. In: S.A. Shepherd, M.J. Tegner and S. Guzmán del Proo (eds.), Abalone of the World. Biology, Fisheries and Culture. Oxford, Fishing News Books. pp. 305-317. [ Links ]
Prince, D.G. and Guzmán del Proo, S. (1993). A stock reduction analysis of the Mexican abalone (Haliotid) fishery. Fish Res, 16: 25-49. [ Links ]
Reinecke-Rodríguez, M.A. (1997). Estructura poblacional, abundancia y aspectos reproductivos de abulón azul Haliotis fulgens de Cabo San Lázaro en Isla Magdalena, B.C.S. Memoria Profesional para obtener el título de Biólogo Marino, UABCS. La Paz, B.C.S., México. 68 pp. [ Links ]
Ricker, W.E. (1975). Computation and interpretation of biological statistics of fish populations. Bull. Fish. Res. Board Can. 191: 382 pp. [ Links ]
SEMARNAP (1996). Anuario Estadístico de Pesca 1996. Dirección de Informática y Estadística de SEMARNAP, México. [ Links ]
SEMARNAP (1997). Anuario Estadístico de Pesca 1997. Dirección de Informática y Estadística de SEMARNAP, México. [ Links ]
Servier-Zaragoza, E., Gómez-López, D. and Ponce-Díaz, G. (1998). The natural diet of the green abalone (Haliotis fulgens Philippi) in the southern part of its range, Baja California Sur, Mexico, assessed by an analysis of gut contents. J. Shellfish Res., 17(3): 777-782. [ Links ]
Shepherd, S.A., Guzmán del Proo, S., Turrubiates, J., Belmar, J. Baker, J. and Sluczanowski, P. (1991). Growth, size at sexual maturity, and egg-per-recruit analysis of the abalone Haliotis fulgens in Baja California. The Veliger 34(4): 324-330. [ Links ]
Shepherd, S.A and Brown, L.D. (1993). What is an abalone stock: Implications for the role of refuge in conservation. Can. J. Fish. Aquat. Sci., 50: 2001-2009. [ Links ]
Shepherd, S.A., Al-Wahaibi, D. and Rashid Al-Azri, A. (1995). Shell growth checks and growth of the Omani abalones Haliotis mariae. In: Shepherd, S.A., R. W. Day and A. J. Butler (eds.), Progress in Abalone Research. Mar. Freshwater Res., 46: 575-582. [ Links ]
Shepherd, S.A. and Ávalos-Borja, A. (1997). The shell microstructure and chronology of the abalone Haliotis corrugata. Moll. Res., 18: 197-207. [ Links ]
Shepherd, S.A and Turrubiates-Morales, J.R. (1997). A practical chronology for the abalones Haliotis fulgens. Moll. Res., 18: 219-226. [ Links ]
Shepherd, S.A., Turrubiates-Morales, J.R. and Hall, K. (1998). Decline of the abalone fishery at La Natividad, Mexico: overfishing or climate change?. J. Shellfish Res., 17(3): 839-846. [ Links ]
Troynikov, V. S., Day, R.W. and Leorke, A.M. (1998). Estimation of seasonal growth parameters using a stochastic Gompertz model for tagging data. J. Shellfish Res., 17(3): 833-838. [ Links ]
Tegner, M.J., Demartini, J.D. and Krpov, K.A. (1992). The California red abalone fishery: a case study in complexity. In: S.A. Shepherd, M.J. Tegner and S. Guzmán del Proo (eds.), Abalone of the World. Biology, Fisheries and Culture. Oxford, Fishing News Books. pp. 370-383. [ Links ]
Turrubiates-Morales, J.R. and Castro-Ortiz, J.L. (1992). Growth of Haliotis fulgens in Bahía Tortugas, Baja California, Mexico. In: S. Guzman del Proo, Tegner, M. J. and Shepherd, S.A. Abalone of the World: Biology, Fisheries and Culture. Fisheries Research Papers. Department of Fisheries (South Australia) 24: 10-15. [ Links ]
Wells, F.E. and Keesing, J.K. (1990). Population characteristics of the abalone Haliotis roei on intertidial platforms in the Perth metropolitan area. J. Malacol. Soc. Australia, 11: 65-71. [ Links ]
Wells, F.E. and Mulvay, P. (1995). Good and bad fishing areas for Haliotis leavigata: a comparison of population parameters. In: S.A. Shepherd, R.W. Day, and J. Butler (eds.), Progress in Abalone Fisheries Research. Mar. Freshwater Res., 46: 519-695. [ Links ]
Wells, F.E. and Keesing, J. (1997). How many juvenile abalone are there? The example of Haliotis roei. Moll. Res. 18: 209-218. [ Links ]
Worhtington, D.G., Andrew, N.L. and Hamer, J. (1995). Covariation between growth and morphology suggests alternative size limits for the blacklip abalone, Haliotis rubra, in New South Wales, Australia. Fish.Bull., 93: 551-561. [ Links ]

