











 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkCarrot (Daucus carota) is a root vegetable that is grown worldwide and used for human consumption in salads, juices, stews and as a garnish. In Mexico, 353,750 t were produced in 2019 with a value of 54,738,948 US dollars. In 2018, Mexico was the 24th largest producer of carrots worldwide (FAO, 2020; SIAP, 2020). As with all crops, carrot is affected by a large number of pathogens e.g., bacteria, fungi, viruses, phytoplasmas and nematodes (Davis and Nuñez, 2007; Seo et al., 2015; Groves et al., 2020). The carrot cyst nematode, Heterodera carotaeJones (1950a), is a cyst-forming nematode (CFN) that is distributed throughout the carrot-growing areas of Europe, North America and South Africa (Subbotin et al., 2010; Yu et al., 2017; Escobar-Avila et al., 2018; Handoo and Subbotin, 2018). This CFN has a limited host range: wild and cultivated carrots and a number of wild members of Apiaceae Family, such as Torilis spp. (Jones, 1950b; Greco, 1986; Mugniery and Bossis, 1988). This nematode completes its life cycle in 36 days at 20 °C and, in an Italian field study, had four generations per year and caused up to 20% yield losses in carrot crops (Greco, 1986; Greco and Brandonisio, 1986). Recently, H. carotae was found to be widely distributed in several municipalities of the Tepeaca Valley, Puebla, Mexico, where its life cycle was completed in 73 days at 20-25 °C (Escobar-Avila et al., 2017; Escobar-Avila et al., 2018). The difference in duration of the life cycle between European and Mexican populations of H. carotae has led us to believe that the number of generations per year and the population dynamics will also be different. Understanding population dynamics is key to design management strategies. The objectives of the present study were: i) to determine the population dynamics of one population of H. carotae in soil and in carrot roots over a ten-month cultivation period under greenhouse conditions; ii) to determine the number of generations of the nematode and its population multiplication capacity; and iii) to determine carrot yield losses at the nematode population densities evaluated.
In June 2017, 90 pots (3 L volume) were placed under greenhouse conditions at 13-20 °C (greenhouse temperature). Sixty of these pots each contained 3 kg of naturally nematode-infested soil and ten carrot seeds (cv. Christian). The naturally nematode-infested soil came from an infested field from Santa Maria Actipan, Acatzingo, Puebla, Mexico (18° 58’ 486’’ N; 97° 50’ 295’’ W; 2246 masl). The soil was thoroughly homogenized and then second-stage juveniles were extracted using the centrifugal flotation technique (Jenkins, 1964) to determine average initial inoculum of H. carotae which was 1200 second stage juveniles (J2) per pot. The thirty remaining pots were controls; each contained 3 kg of the same soil, except that it had been tindalized, and 10 carrot seeds (cv. Christian) placed. The experiment was set up in a completely randomized design. All pots were maintained in the greenhouse over a 10-month period (June 2017 - April 2018) and were watered every 3 days with 20 mL of tap water. Immediately after the seeds had germinated, pots were destructively sampled to determine nematode population densities; three treatment pots were evaluated every 15 days and three control pots were evaluated every 30 days. The roots and soil were evaluated separately from each pot. To determinate the fresh weight of carrot roots, these were gently shaken, washed and then foliage was removed. From each pot, the roots of each plant were individually weighed and then a mean per pot was determined. As control pots were only evaluated monthly, only data from treatment pots sampled at the same time as control pots were used in statistical analysis of fresh weight. A t-test was performed to identify any significant differences between the weight of the roots when nematodes were present or absent (p<0.05). Analysis was done using the SygmaPlot 12.0 program. Carrot yield loss was calculated as the percentage loss between the root weight of control pots and infested pots. After weighing, all of the roots from each pot were carefully washed with tap water and then stained using acid fuchsin lactoglycerol to observe and count nematode stages inside and on the surface of roots (J2, J3, J4, females and cysts) (Byrd et al., 1983). All nematode stages were recorded and a mean nematode density per stage and date was calculated. The total mean nematode density was taken as the 100 per cent for each date. On the other hand, the soil from each pot was thoroughly homogenized and then J2s and cysts were extracted from 200 cm3 of soil using the centrifugal flotation technique (Jenkins, 1964) and Fenwick’s method (Fenwick, 1940), respectively. The mean number of cysts were used to calculate the population multiplication factor by dividing the final population (Pf) density from each date by the initial population (Pi) density. The value of Pi was taken from the immediate previous date of sampling.
Overall, the fresh weight of carrot roots was significantly smaller in the nematode treatment pots than in the control pots (p<0.05) (Table 1). From the beginning of the experiment and up to 60 days post germination (DPG), the fresh root weight was not significantly different between the treatment and control pots (p>0.05). However, from 90 DPG, there was a significant difference in the root fresh weight between treatment and control pots on every sampling time. At the end of the experiment (300 DPG), an average 70.8% yield loss was observed. Inside carrot roots, J2 were observed at 15 DPG, but as DPG increased the number of J3, then J4 and female stages increased in number (Figure 1). By 75 DPG, all nematode stages were present in various proportions, but cysts were the predominant stage (Figure 1). Five generations of the nematode were completed during the entire experimental period; peaks of females were observed at 60, 105, 195, 225 and 255 DPG with means of 172, 224, 228, 264 and 235 females, respectively (Figure 2A). As for nematode population in soil, three maximum peaks in J2 populations were observed at 105, 150 and 195 DPG with a mean of 735, 882 and 967 J2/200 cm3 of soil respectively. A similar trend in J2 populations was observed inside carrot roots (Figure 2B). The multiplication factor calculated from cysts was inversely related to the initial population densities and varied between 230 and 558 cysts/200 cm3 of soil (Figure 2C). At low nematode Pi densities, the multiplication factor was the greatest. In contrast, when nematode Pi densities were at their highest, the multiplication factor was at its smallest (Figure 2C). A multiplication factor of 1.6 was calculated at the end of the experiment.
Table 1 Fresh root weight of Heterodera carotae infested and control carrots over a ten-month culture period under greenhouse conditions.
| Fresh carrot root weight (g) | |||||
|---|---|---|---|---|---|
| DPG | Control | Infested with H. carotae | p | Yield loss (%) | |
| 30 | 0.006 ± 0.0008 | 0.005 ± 0.00008 | 0.751 | 16.7 | |
| 60 | 0.019 ± 0.0033 | 0.018 ± 0.0087 | 0.849 | 5.3 | |
| 90 | 0.066 ± 0.003 | 0.039 ± 0.0126 | 0.023* | 40.9 | |
| 120 | 3.547 ± 1.5 | 0.242 ± 0.11 | 0.019** | 93.2 | |
| 150 | 7.039 ± 0.985 | 1.519 ± 1.06 | 0.003** | 78.4 | |
| 180 | 8.402 ± 2.559 | 0.528 ± 0.0599 | 0.006** | 93.7 | |
| 210 | 9.963 ± 3.619 | 1.745 ± 0.223 | 0.017** | 82.5 | |
| 240 | 10.192 ± 3.467 | 1.406 ± 0.352 | 0.012** | 86.2 | |
| 270 | 13.712 ± 4.255 | 1.659 ± 0.297 | 0.008** | 87.9 | |
| 300 | 16.599 ± 5.379 | 3.168 ± 0.367 | 0.012** | 80.9 | |
Each value is the mean of three replicates + standard deviation. Significant differences were determined using a t-test. *= significant (p0.05); **= highly significant (p0.01). Average yield loss = 70.8%. DPG = days post germination.
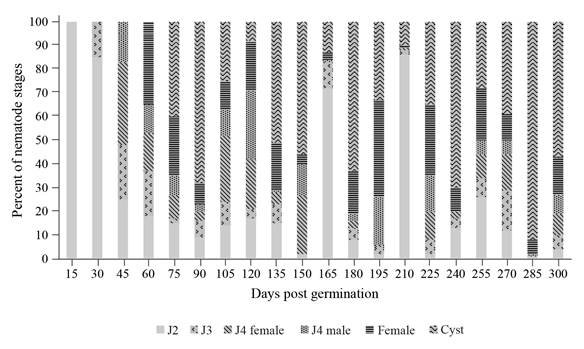
Figure 1 Stages of the cyst nematode Heterodera carotae present within roots of carrots over a ten-month culture period under greenhouse conditions. The total mean nematode density from the three replicates was taken as the 100 per cent for each date.
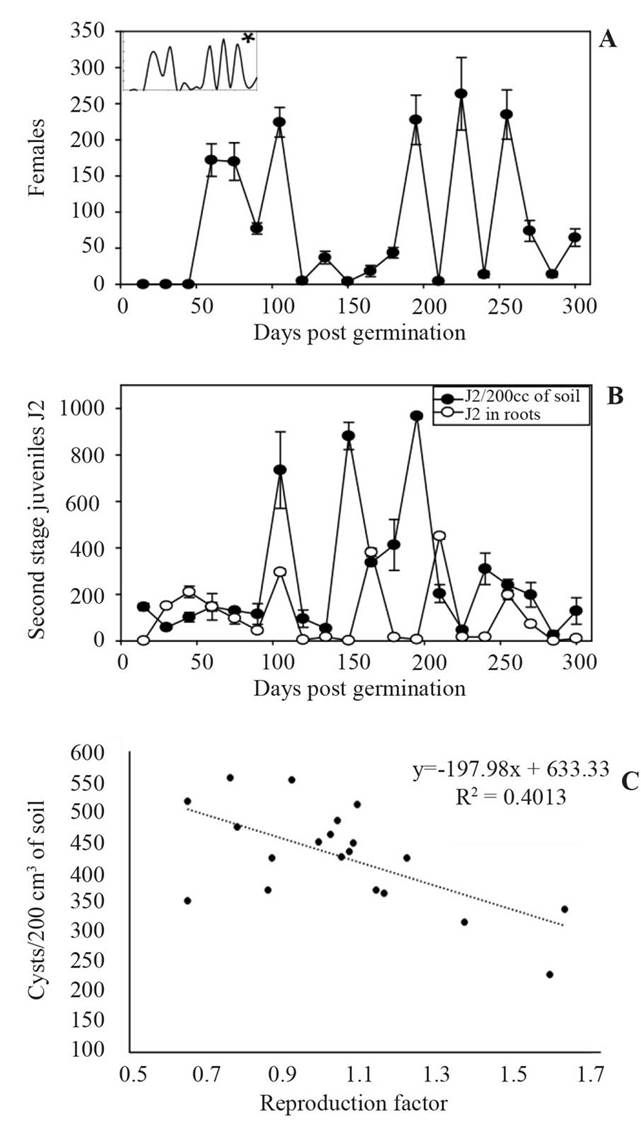
Figure 2 Population dynamics and multiplication factor of Heterodera carotae in soil and roots of carrot cv. Christian over a ten-month culture period under greenhouse conditions. A) Population dynamics of Heterodera carotae females in carrot roots. Average number of females from three replicates; * = Simple spline curve representation. B) Population dynamics of second stage juveniles (J2) of H. carotae in soil and carrot roots. Average number of J2 from the three replicates. C) Relationship between H. carotae multiplication factor and initial population densities on carrot.
In this study, the carrot cyst nematode Heterodera carotae caused an average 70.8% yield loss in carrot over a ten-month culture period from June 2017 to April 2018 under greenhouse conditions. This is much higher than those previously reported by Greco et al. (1993), which was a maximum of 20% under Italian conditions. Ambrogioni and Marinari-Palmisano (1976) determined a tolerance limit, i.e. the maximum nematode density at which no yield losses occur (Seinhorst, 1986: Schomaker and Been, 2006), of 0.19 eggs/cm3 of soil in a field trial, while Greco and Brandonisio (1980) determined a tolerance limit of 0.8 eggs/cm3 of soil in a microplot experiment. In the latter study, yield losses of 40 and 70% occurred at densities of 16 and 32 eggs/cm3 of soil, respectively (Greco and Brandonisio, 1980). In our study, the initial nematode density was 1200 J2/three kg of soil, which is equivalent to 0.4 J2 per g of soil. It is difficult to compare our results with previous work because the initial nematode inoculum concentrations were not the same. However, the initial nematode density in our study was lower than the one reported by Greco and Brandonisio (1980) and yet still caused a yield loss of 70%. In addition, the inoculum concentration used in our study was, theoretically, within the tolerance limit, and so no yield losses should have been observed (Greco and Brandonisio, 1980). The difference in our results and previous reports may be due to a number of factors. Firstly, the plant varieties used in the experiments were different and it is known that some varieties of carrots are more susceptible to nematodes than others (Barrot, 2018). It is possible that carrot cv. Christian is more susceptible than cv. Vilmorin 66, which Greco and Brandonisio (1980) used. Secondly, temperature and type of soil play an important role in emergence and development of H. carotae (Greco and Brandonisio, 1986). The optimum temperature for emergence of H. carotae J2 is between 15 and 20 °C, although they can invade plants and develop at 10 °C (Greco and Brandonisio, 1986; Greco, 1987). Our study was conducted at 13-20 °C, therefore there were no temperature restrictions on emergence and development. Thirdly, the specific Mexican population (Santa Maria Actipan) of H. carotae used in this experiment could be more virulent than the European population. Further studies on virulence of the Mexican population are needed. Inside carrot roots, J2, J3, J4, females and cysts were observed after 15, 30, 45, 60 and 75 DPG, respectively at 13-20 °C. These results agree with those obtained by Escobar-Avila et al. (2017) who evaluated the same population in carrot cv. Mexicana and observed J2, J3, J4, females and cysts after 10, 24, 38, 59 and 73 DPG, respectively, under greenhouse conditions at 20-25 °C. In contrast, in Italy the life cycle of this nematode species was completed within 36 days at 20 °C in carrot cv. Selection 92 (Greco and Brandonisio, 1986). According to Greco and Brandonisio (1986) the temperature range for development of H. carotae is 10-20 °C and that above 25 °C nematode development stops. Therefore, in the months when the average temperatures are high, the life cycle may be longer (Greco, 1987; Mugniery and Bossis, 1988). However, this was not a factor in our study in which the temperature was maintained between 13 and 20 °C. Factors that might influence the duration of the life cycle (twice as long for the Mexican population compared with the Italian population) is the carrot variety used and the particular characteristics of the Mexican nematode population evaluated. Heterodera carotae has an interesting biology. Approximately half of the eggs are retained within the cyst, while the other half are laid in a gelatinous matrix secreted by the anal glands of the female (termed egg masses). Under suitable conditions, eggs in the egg masses hatch as soon as embryogenic development is completed. In contrast, hatching of eggs within the cysts is delayed; these eggs usually require a two-month period and the presence of host plant root exudates before they hatch. Therefore, H. carotae is capable of hatching all year round (Winslow, 1955; Aubert, 1986; Greco and Brandonisio, 1986; Greco, 1987; Baldwin and Mundo-Ocampo, 1991). This is unlike other species of cyst nematode that only complete one generation per year and require a dormancy period before hatching (e.g. the cereal cyst nematodes H. avenae, H. filipjevi, H. latipons and the potato cyst nematode Globodera rostochiensis). The peculiar biology of H. carotae explains why all nematode stages where found at the same time inside carrot roots during the experiment; after the first appearance of females, eggs within egg masses were continuously hatching and infesting carrot roots. Five generations of the nematode were completed during the ten-month culture period. In the first four months of the experiment, two nematode generations were completed (60 and 105 DPG), which corresponds to one typical carrot growing season. After sixth months, another three nematode generations were observed with a 30-day period between each one (195, 225 and 255 DPG). It seems that, at first, the nematode required some time to establish its population leading to long life cycles (75 days), but then the life cycle shortened and generations were completed every 30 days. During the experiment, the highest numbers of J2 in the soil were observed at 105, 150 and 195 DPG; a similar trend was observed inside carrot roots. In the first generation of females the number of J2 in soil was constant. From the second generation of females the J2 numbers peaked at 105 DPG and the timing suggests they were most probably derived from egg masses. At 150 days, a second peak of J2 was observed but there was no increase in the numbers of females or cysts in the roots; this second peak of J2 (150 days) was observed 60 days after the largest number of cysts was observed in roots (90 days). As the eggs inside cysts require a two-month period before they can hatch it is likely that this second peak of J2 was derived from the cysts (Greco and Brandonisio, 1986). The timing of the third peak of J2 suggests they were derived from egg masses of the third generation of females (195 days). These results demonstrate the high multiplication capacity of this species, especially in continuous carrot crops, as is done for carrot seed production. The multiplication factor was inversely related to initial population densities. This means that when the initial population is high, the multiplication factor is low. This is due to competition for space and food in large nematode populations (Seinhorst, 1966; Hajihasani et al., 2010). Studies on population dynamics of the carrot cyst nematode in soil and roots are scarce. Moreover, they have generally been conducted over a shorter period (4-5 months only). Therefore, our findings provide novel and important information that can be used in the development of integrated management strategies and decision- making tools to optimize the timing of control applications.
CONCLUSIONS
To be able to develop control strategies that are effective against the Carrot Cyst Nematode (CCN), Heterodera carotae, the knowledge of population dynamics is indispensable. The results obtained in this study showed three second-stage juveniles (J2’s) peaks at 105, 150 and 195 post-germination days, at which time, control strategies that are effective against this stage of the CCN can be applied. Additionally, a multiplication factor of 1.6 was calculated and five generations of the nematode were observed. The CCN caused an average 70.8% yield loss during the experiment.

