











 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkLa cantidad y calidad de la producción en el cultivo de jitomate disminuye por efecto de las enfermedades provocando pérdidas millonarias (Johnson et al., 2015; Collinge et al., 2010; Saville et al., 2015; Childers et al., 2015). La interacción patógeno-hospedero es afectada por factores bióticos y abióticos. Los primeros incluyen resistencia, heterogeneidad y demografía del hospedero, así como densidad, frecuencia y relación genética de genotipos del patógeno coexistente (Zhan y McDonald, 2013). El principal patógeno del jitomate es Phytophthora infestans y diversos factores pueden incrementar su patogenicidad (Dudler, 2013; Schornack et al., 2013; Berdúo- Sandoval et al., 2019). La migración de sus dos tipos de apareamiento (A1 y A2) provocan su reproducción sexual incrementando su variabilidad genética (Skelsey et al., 2009). En tierras altas de México, la variabilidad genética de P. infestans, bajas temperaturas (10-15 °C) y alta humedad relativa (> 90% 10 h día-1) son favorables para su expresión patogénica (Díaz et al., 2014). El patrón de dispersión del patógeno también depende de la ubicación del campo, distribución espacial, duración de supervivencia de esporas (Skelsey et al., 2010), y radiación solar (Skelsey et al., 2009). La liberación de esporangios está correlacionada estrecha y directamente con niveles bajos de radiación solar, temperatura y alta humedad relativa (Díaz et al., 2014).
La temperatura y humedad relativa tienen un impacto significativo en el desarrollo del tizón tardío. Los efectos diferenciales, como lluvia, viento, humedad relativa, temperatura y luz pueden influir en la eficacia de esporangios para iniciar la infección (Ojiambo et al., 2015) siendo importantes para la epidemiología y manejo de la enfermedad (Danies et al., 2013). En condiciones climáticas favorables para el patógeno, el cultivo de jitomate puede destruirse en cuestión de días (Hu et al., 2012) y cuando las condiciones son desfavorables puede reducirse su incidencia y/o severidad (Bostock et al., 2014). Predecir la enfermedad necesita una sólida comprensión de la relación entre datos climáticos y desarrollo de la enfermedad (Yuen y Mila, 2015). “LATEBLIGHT” es un modelo predictivo que evalúa enfermedades policíclicas de hojas y predice el nivel de resistencia genética (Forbes et al., 2008; Yuen y Forbes, 2009). Estudios no definen el momento y la cantidad de infección inicial por varias razones, incluidas las limitaciones de tiempo relacionadas con el inicio de la epidemia después del trasplante (Andrade-Piedra et al., 2005a; Díaz et al., 2014). El área bajo la curva de progreso de la enfermedad (AUDPC) permite cuantificar el grado de resistencia genética a P. infestans en condiciones de campo, a través de la estimación del área bajo la curva de infección real. AUDPC relativo (RAUDPC) se usa para comparar el progreso de la enfermedad en diferentes condiciones ambientales o cuando se comparan más de una estación de crecimiento (Díaz et al., 2014).
Phytophthora infestans exhibe un amplio rango de diversidad genética en Chapingo, México (Alarcón-Rodríguez et al., 2014). Esta región tiene una temporada de lluvias de verano constante, lo que garantiza la infección natural del tizón tardío. Bajo estas condiciones y evaluando la resistencia genética de varias líneas de jitomate, se realizó el presente estudio con el objetivo de documentar y cuantificar el progreso de la infección foliar por P. infestans en diferentes líneas de jitomate con diferentes niveles de resistencia genética, bajo tres escenarios: (1) esperado, basado en el AUDPC derivado de promedios históricos de variables; (2) observaciones directas de campo, y (3) simulado, utilizando el modelo matemático predictivo “LATEBLIGHT” versión LB2004.
Materiales Y Métodos
Sitio de estudio. Chapingo se encuentra a 2,250 m sobre el nivel del mar (19° 30’ latitud norte y 98° 53’ longitud oeste) en el Estado de México. Tiene clima Cb (Wo) con temperatura anual promedio de 15.2 °C y precipitación anual de 635.5 mm (García, 1987). Las temporadas de cultivo en esta región dependen de la lluvia y se extienden desde mediados de junio hasta mediados de octubre.
Patosistema Jitomate/Tizón. El patosistema de P. infestans-jitomate se estudió bajo condiciones climáticas de Chapingo, Estado de México durante dos ciclos de cultivo (2017 y 2018) en un periodo de cinco semanas que comprendieron los meses de agosto y septiembre, utilizando tres escenarios: (1) esperado, (2) observado y (3) simulado. En el experimento se utilizó un control (río grande) y seis líneas de jitomate (L-03, L-51, L-72, L-92, L-43, L-77). El escenario esperado consiste en la obtención de la cantidad de posibles ciclos de infección de P. infestans que podrían existir durante el ciclo de cultivo. Este se obtiene del análisis de la asociación de variables meteorológicas con factores que influyen en el progreso de P. infestans (Díaz et al., 2014). El escenario observado es el progreso de infección real de P. infestans durante un ciclo de cultivo determinado. Este consiste en la evaluación de lecturas de infección del follaje utilizando la escala de Henfling para así obtener el AUDPC (Henfling, 1987). El escenario simulado consiste en la predicción del AUDPC utilizando el algoritmo “LATEBLIGH” el cual se ejecuta con el suministro de datos de parámetros climáticos, resistencia del hospedero y variables epidemiológicas (Díaz et al., 2014; Andrade-Piedra et al., 2005b; Andrade-Piedra et al., 2005c).
Manejo agronómico del cultivo. En 2017 la semilla se sembró el 13 de mayo y fueron trasplantadas el 16 de junio. En 2018 la semilla se sembró el 14 de mayo y trasplanto el 21 de junio. En ambas fechas la semilla se plantó en charolas de unicel de 200 cavidades. El trasplante para ambas fechas se realizó en unidades experimentales conformadas por tres surcos con un área de 12 m2 de cada unidad experimental. El experimento se ubicó en el lote X-2 del campo agrícola experimental Xaltepa de la Universidad Autónoma Chapingo. Se realizaron riegos de auxilio cuando el cultivo lo requería mediante la utilización de cintilla (calibre ocho mil, con goteros cada 20 cm y con un gasto por emisor de 1.02 litro•h-1.); se aplicó fertilización granulada N, P, K (46-00-00; 18-46-00; 00-00-60). Se aplicó fertilización foliar de micro elementos (Tradecorp AZ). Para el control de plagas se aplicó flonicamid y cipermetrina. El control de malezas fue manual. De acuerdo a la etapa fenológica del cultivo se le dio soporte a la planta con rafias de nilón.
Escenario esperado. Se analizaron seis variables meteorológicas sobre el desarrollo del tizón tardío (Cuadro 1), mediante la aplicación regresión lineal múltiple (SAS, 2002). La selección del mejor modelo, con variabilidad mínima de independencia, se basó en el criterio del coeficiente de determinación más alto. Los datos meteorológicos se asociaron con condiciones que influyen en la supervivencia, permanencia y diseminación de P. infestans (Andrade-Piedra et al., 2005c; Iglesias et al., 2010; Johnson et al., 2009; Morales et al., 2011; Nowicki et al., 2012; Skelsey et al., 2009; Skelsey et al., 2010). Temperatura (T), humedad relativa (HR) y radiación solar (RS) fueron seleccionadas para determinar los escenarios esperados considerando su mayor contribución al desarrollo de la enfermedad.
Cuadro 1. Valores límites mínimos y máximos de los intervalos de las variables meteorológicas registradas de Junio 1 a septiembre 30, de 2007 a 2016 en Chapingo, México.
| Variable | Abreviación | Unidad | 80%≤HR≤90% Rango | HR>90% Rango |
|---|---|---|---|---|
| Temperatura | T | °C | 12.7-17.6 | 12.7-16.3 |
| Punto de roció | PR | 12.1-16.7 | 12.3-16 | |
| Precipitación | PP | mm | 0-47.1 | 2.8-21.4 |
| Velocidad del viento | VV | ms-1 | 0-1 | 0-1.5 |
| Velocidad máxima del viento | VMV | ms-1 | 0-1 | 0-6.3 |
| Radiación solar acumulada | RS | W m2 | 3018y | 1701z |
y acumulado durante 23.5 h, de 00:00 a 23:30, 3,606 W m-2 ≈ 12.982 MJ m-2.
z acumulado durante 3 h, de 06:00 a 09:00, 1,025 W m-2 ≈ 3.69 MJ m-2
Escenario observado. Se evaluaron seis líneas de jitomate y un control con dos niveles de susceptibilidad (S = susceptible, R = resistente, C= control); L-03 (S), L-51 (S), L-72 (S), L-92(S), L-43 (R), L-77(R), y río grande (C) (Leyva et al., 2013). Durante el ciclo del cultivo se fertilizaron las plantas, se realizó el control de malezas y plagas. En campo fueron evaluados con un diseño experimental de bloques completos al azar con cuatro repeticiones (5 m de largo, 0.80 m entre filas y 0.40 m entre plantas). Las lecturas semanales de infección de follaje se evaluaron con la escala de Henfling (Henfling, 1987) en un periodo de cinco semanas comprendidas entre agosto y septiembre de los años 2017 y 2018 una vez que el patógeno se presentó de forma natural. Con base en síntomas iniciales después del trasplante, el AUDPC se calculó progresivamente (Shaner y Finney, 1977). Los valores derivados del AUDPC se usaron para calcular RAUDPC (Fry, 1978; Bonierbale et al., 2010). Finalmente, los valores de RAUDPC se usaron para calcular RaRAUDPC, utilizando como referencia la línea 92 que fue susceptible durante el experimento, siguiendo el procedimiento descrito por Yuen y Forbes (2009) y Taipe et al. (2011).
Escenario simulado. Se elaboró un algoritmo denominado LATEBLIGH en el “Sistema SAS para Windows 9.0” (SAS, 2002) considerando parámetros climáticos y nivel de resistencia del hospedero dentro de 2.4 m2 de plantas de jitomate, basado en formularios reportados por Díaz et al. (2014).
Resultados Y Discusión
Escenarios esperados. Al seleccionar el modelo de regresión lineal múltiple para la realización de los escenarios esperados se encontró que temperatura (T), humedad relativa (HR) y radiación solar (RS) fueron variables independientes que presentaron el mayor coeficiente determinación (R2 = 0.44). Estas variables están asociadas con la presencia, diseminación y efectividad de P. infestans. Se registraron variables meteorológicas en un periodo de 10 años (2007-2016) y se promediaron en el gráfico de tendencia. En el Cuadro 1, se muestran los registros promedio mínimo y máximo de variables meteorológicas derivadas de la aplicación de dos criterios de selección para HR (80% ≤HR≤90% y HR> 90%). El límite superior de los registros promedio de T, HR y RS bajo el criterio del 80% ≤RH≤90% se utilizó para explicar tanto la variabilidad como el efecto de estas variables en el desarrollo del tizón tardío. Esta comparación se hizo utilizando los parámetros meteorológicos para los escenarios esperados con HR durante 23.5 h y T 15.9 °C favoreciendo la presencia, difusión y efectividad de P. infestans, en comparación con los límites superiores de la media (Andrade-Piedra, et al., 2005c; Iglesias et al., 2010). Basado en las condiciones obtenidas a través de T, HR y RS como el mejor modelo lineal múltiple, mayor pico fluctuación y selección de criterio 80% ≤RH≤90%, se identificaron cinco escenarios esperados con rangos de cuatro a diez ciclos de infección de longitud variable de 6 a 12 h por la noche, en la madrugada y al amanecer (19:00 a 08:00). En la Figura 1, se presentan los ciclos de infección de los escenarios esperados (I a V). Las curvas de tendencia (picos altos y bajos) se unen para las tres variables (T, HR y RS). El principio como el final de cada ciclo de infección está determinado por la disminución de la HR e incremento en la T, basado en intervalos diarios de 60 min. Se requiere clima frío y húmedo para el desarrollo de una mayor cantidad de inóculo en un corto período de Tiempo (Kamoun y Smart, 2005). La oscuridad favorece la esporulación (Nowicki et al., 2012) y la expresión de los patógenos en Chapingo (Díaz et al., 2014). La severidad fue favorecida con T de 9.9 a 16.1 °C y HR de 80.4 a 100% durante 8 h en el cultivo de jitomate desde el 1 de junio hasta el 18 de septiembre (Cuadro 2). Estas condiciones favorecen el inicio de una epidemia con temperaturas nocturnas por debajo del punto de condensación durante al menos 4 h y baja temperatura nocturna igual o superior a 10 °C (Díaz et al., 2014; Morales et al., 2011; Nowicki et al., 2012). Se observaron diferencias significativas (p<0.05) entre los registros promedio para las variables meteorológicas T y HR en los cinco escenarios esperados analizados.

Figura 1 Ciclos de infección de P. infestans de cinco escenarios esperados en jitomate bajo las condiciones climáticas de Chapingo, Estado de México. (Temperatura (°C), Humedad relativa (%), Radiación solar (Wm-2).)
Escenarios observados. En 2017, la tolerancia genética de Río grande y Línea 77 fue mejor a las Líneas susceptibles (Línea 92 y Línea 72) las cuales fueron utilizadas como referencias en el progreso inicial de P. infestans y en 2018, la expresión de la línea tolerante (Río Grande) fue mayor a líneas susceptibles (Línea 43, Línea 51). En 2017, la infección observada en campo inició el mismo día, dando la idea de que ambas líneas son susceptible a P. infestans bajo condiciones favorables de HR y TR. De igual manera en 2018, la infección en la línea 92 y Río grande inició el mismo día con una velocidad similar de infección en comparación con 2017 acorde con el plan meteorológico de condiciones de ese año (Figura 2). Las condiciones climáticas fueron favorables para la infección de hojas en líneas susceptibles. A los 90-91 días después del trasplante (ddt), las plantas de las líneas susceptibles fueron 73% infectadas durante 2017. Un retraso similar en las tasas de infección también fue informado por Cadena-Hinojosa et al. (2007).
Cuadro 2. Escenariosesperados y ciclos de infección por escenario para P. infestans en jitomate bajo las condiciones climáticas de Chapingo.
| Escenario | Ciclo de infección | Número de horasy | Días Julianos | Horario | Variables meteorológicas | |||
|---|---|---|---|---|---|---|---|---|
| H_hz | T | HR | RS | |||||
| I | 1 | 673-681 | 11-12 | 24:00-08:00 | 8 | 14.4-15.2 | 84.6-96 | 166.8 |
| 2 | 692-704 | 12-13 | 19:00-07:00 | 12 | 14.3-15.5 | 85.7-92.9 | 289.4 | |
| 3 | 716-727 | 13-14 | 19:00-06:00 | 11 | 13-14.5 | 98.4-100 | 83.7 | |
| 4 | 743-753 | 15-16 | 22:00-08:00 | 10 | 12.5-15.5 | 80.4-91.7 | 269 | |
| 5 | 766-777 | 16-17 | 21:00-08:00 | 11 | 14.1-15.6 | 90.8-94.8 | 16.6 | |
| Promedio | 10.4 | 13.6b-15.2a | 87.9a-95b | 165.1 | ||||
| II | 1 | 1244-1257 | 35-36 | 19:00-08:00 | 13 | 10.3-13.9 | 100-100 | 298.4 |
| 2 | 1268-1281 | 36-37 | 19:00-08:00 | 13 | 12.1-14.3 | 91.1-97.9 | 46.05 | |
| 3 | 1293-1306 | 37-38 | 20:00-09:00 | 13 | 12.1-14.4 | 91.3-99.5 | 264.65 | |
| 4 | 1318-1330 | 38-39 | 21:00-09:00 | 12 | 12-13.7 | 98.5-100 | 331.85 | |
| 5 | 1341-1352 | 39-40 | 20:00-07:00 | 11 | 12.7-15.2 | 82.2-97.2 | 219.95 | |
| 6 | 1365-1375 | 40-41 | 20:00-07:00 | 11 | 12-16.1 | 83.3-94.7 | 91.02 | |
| Promedio | 12 | 11.8b-14.6a | 91a-98.2ab | 190.04 | ||||
| III | 1 | 1897-1905 | 62-63 | 00:00-09:00 | 8 | 11.9-15.3 | 85.4-97.6 | 323.84 |
| 2 | 1916-1927 | 63-64 | 19:00-06:00 | 11 | 11.4-15.2 | 85-95.9 | 8.39 | |
| 3 | 1943-1951 | 64-65 | 22:00-06:00 | 8 | 8.9-14.2 | 80.1-99.1 | 166.28 | |
| 4 | 1967-1975 | 65-66 | 22:00-06:00 | 8 | 10.2-14.8 | 85.1-96.1 | 65.7 | |
| 5 | 1992-2000 | 66-67 | 23:00-06:00 | 8 | 10.3-13.7 | 85.2-99.7 | 97.02 | |
| Promedio | 8.6 | 10.5C-14.6a | 84.1a-97.6ab | 137.246 | ||||
| IV | 1 | 2137-2146 | 73 | 00:00-09:00 | 9 | 13-15.1 | 97.1-100 | 31.63 |
| 2 | 2157-2169 | 73-74 | 20:00-07:00 | 11 | 12.7-15.6 | 80.8-100 | 0 | |
| 3 | 2181-2191 | 74-75 | 20:00-06:00 | 10 | 12.5-13.3 | 99.6-100 | 60.76 | |
| 4 | 2204-2216 | 75-76 | 19:00-07:00 | 12 | 10.8-16.2 | 81.8-99.7 | 6.9 | |
| 5 | 2228-2241 | 76-77 | 19:00-08:00 | 13 | 12.2-16 | 89.5-99.4 | 575.75 | |
| 6 | 2254-2265 | 77-78 | 21:00-08:00 | 11 | 12.9-14.7 | 88.4-96.7 | 16.3 | |
| 7 | 2277-2290 | 78-79 | 20:00-09:00 | 13 | 12.9-14.5 | 99.1-100 | 134.21 | |
| 8 | 2301-2313 | 79-80 | 20:00-08:00 | 12 | 12.7-17 | 81.2-93.6 | 0 | |
| 9 | 2325-2337 | 80-81 | 20:00-08:00 | 12 | 12.8-15.9 | 87.3-100 | 0 | |
| 10 | 2349-2362 | 81-82 | 20:00-09:00 | 13 | 11.6-13.9 | 99.4-100 | 32.37 | |
| Promedio | 12.2 | 12.4b-15.2a | 90.4a-98.9a | 36.57 | ||||
| V | 1 | 2545-2551 | 90 | 00:00-06:00 | 6 | 9.9-14.9 | 81.1-98.3 | 66.48 |
| 2 | 2567-2576 | 90-91 | 22:00-07:00 | 9 | 10.9-13.9 | 82.7-93.1 | 45.02 | |
| 3 | 2589-2600 | 91-92 | 20:00-07:00 | 11 | 11.5-15.7 | 86.1-99.3 | 30.8 | |
| 4 | 2613-2625 | 92-93 | 20:00-08:00 | 12 | 10.3-17.4 | 85.2-100 | 441.03 | |
| Promedio | 10.5 | 10.6c-15.4a | 83.7a-97.6ab | 123.98 | ||||
* Los promedios con letras similares no son significativamente diferentes (prueba t de relación K de Waller-Duncan, p<0.05).
y No. 0.5 h = número de segmentos de 30 min acumulados, del 1 de junio al 18 de septiembre (104 días) de cada año.
z H_h horas acumuladas para RH> 80%, temperatura mínima y máxima T (°C), humedad relativa mínima y máxima HR (% ), RS Radiación solar acumulada (Wm-2).
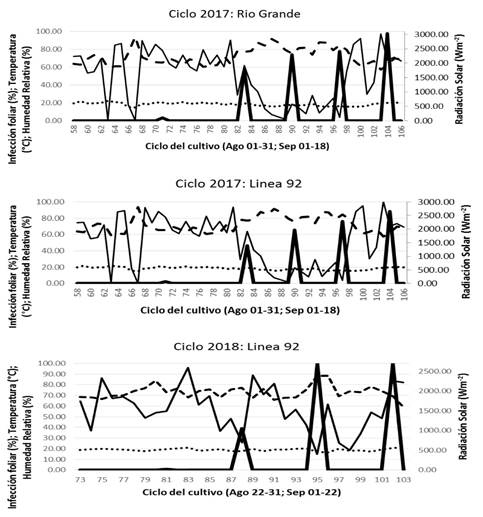
Figura 2. Progreso de la infección por P. infestans en una línea de jitomate susceptible durante dos temporadas de campo (2017, 2018) y una tolerante (2017) bajo las condiciones climáticas de Chapingo. (Temperatura (°C), Humedad relativa (%), Radiación solar (Wm-2), P. infestans).
El progreso de la infección en la Línea 92 (plantas muertas en 100-104 ddt demostró la presencia de condiciones ambientales favorables para la propagación del patógeno, con temperatura de 10.6-15.4 °C, HR entre 83-98% durante 12 h, durante la noche, madrugada y parte de la mañana (20:00 a 08.00 h) coincidiendo con el escenario VI-4 esperado. Sin embargo, en 2018, la infección inició y progresó más tarde (en aproximadamente 79-81 ddt) en comparación a 2017, con 12-16.2 °C, 90-99% HR, que coincide con el escenario 6, 7, 8 (Cuadro 3).
En 2018, las plantas no presentan infección a los 95-96 ddt, 10-14.6 °C, y 100% HR durante 12 h en la noche, madrugada y al amanecer (20:00-08:00 h) siendo similar al escenario esperado IV-4. En general, durante los dos ciclos de cultivo (2017 y 2018), el progreso de la infección por P. infestans fue favorecido por escenarios con T de 10-16.7 °C y HR 80-100% durante 8-13 h en la noche, madrugada y parte de la mañana (19:00-08:00 h) (Andrade-Piedra, et al., 2005c; Iglesias et al., 2010). El promedio de registros de 60 minutos durante el ciclo de cultivo de 102-104 días en 2017 y 2018, no mostraron diferencias significativas entre magnitudes de T (14.6 °C vs 15.4 °C) y HR (80% vs 83%). Sin embargo, condiciones favorables para la progresión natural de la enfermedad fueron observados y esperados (basado en los escenarios modelados). En 2017, el inicio observado de la infección coincidió con los escenarios I-2; en 2018, coincidió con los escenarios II-3 y IV-5. En 2017, el punto observado donde las plantas no presentan infección coincide con el escenario IV-6, y en 2018 con el escenario II-1 (Cuadro 3).
Cuadro 3. Progreso de los niveles de infección por P. infestans en los escenarios observados y esperados en jitomate Línea 92, bajo las condiciones ambientales de Chapingo, México.
| Ciclo del cultivox | DAT_ty | Horario | Variables meteorológicasz | Infección (%) | Escenario Esperado | ||
|---|---|---|---|---|---|---|---|
| h | T | HR | |||||
| 2017 I | 70-71 | 19:00-07:00 | 12 | 9.8-17 | 84.4-100 | 3.1 | I-2 |
| 2017 M1 | 82-83 | 19:00-08:00 | 13 | 12.2-16 | 87.2-100 | 56.6 | IV-4, IV-5 |
| 2017 M2 | 89-90 | 19:00-08:00 | 13 | 11.8-16 | 83.8-99.2 | 73.8 | |
| 2017 M3 | 96-97 | 19:00-08:00 | 13 | 12.1-15 | 83.3-100 | 77.5 | |
| 2017 F | 103-104 | 21:00-08:00 | 11 | 11.5-16.7 | 83-99.3 | 97.5 | IV-6 |
| 2018 I | 80-81 | 19:00-08:00 | 13 | 12-16.2 | 82.8-98.7 | 0.9 | II-3, IV-5 |
| 2018 MI | 87-88 | 19:00-08:00 | 13 | 11.9-15.8 | 83-100 | 39.1 | II-2 |
| 2018 F | 94-95 | 19:00-08:00 | 13 | 10-14.6 | 98.9-100 | 100 | II-1 |
x I Fase temprana de la infección, M fase intermedia de la infección, F Fase final de la infección, 2017 I= 14 agosto 2017; 2017 M1= 25 al 26 agosto; 2017 M2= 1 al 2 septiembre; 2017 M3= 08-09 septiembre; 2017 F= 15 al 16 septiembre; 2018 i= 29 al 30 agosto; 2018 M= 05 al 06 septiembre; 2018 F= 12 al 13 septiembre.
y DAT_t1 = primera evaluación cuando se observaron lesiones causadas por P. infestans (número de días después del transplante).
z H_h rango diario de horas con RH≥80%, rango de temperatura diaria de T (°C), rango de humedad relativa diaria de HR (%).
El AUDPC, RAUDPC y RaRAUDPC fueron evaluados en un diseño experimental aleatorizado con cuatro bloques durante 2017 y 2018. Los resultados validaron el modelo propuesto, siendo este confirmado por el desempeño homogéneo de la varianza a través de un análisis de residuos y la prueba de normalidad de Shapiro-Wilk, (SAS, 2002) (Cuadros 4 y 5).
En 2017, la estadística de las pruebas produjo el valor de 0.45 para AUDPC, RAUDPC y RaRAUDPC con Pr <W 0.0001 y en 2018, los tres valores fueron de 0.94. La normalidad de Shapiro-Wilk valida el modelo propuesto con un rendimiento homogéneo de varianza durante las dos temporadas de campo. AUDPC, RAUDPC, y RaRAUDPC fueron analizados individualmente sin encontrar diferencias entre tratamientos con P <0.0001. Durante cada ciclo de cultivo, el efecto de orden de mérito AUDPC, RAUDPC, y RaRAUDPC fue mayor en las líneas susceptibles (línea 92) en comparación con tolerantes (Río grande). El t-test (LSD) y Tukey’s Valores de Studentized Test (HDS) para la variable AUDPC y sus derivados indican que todas las posibles comparaciones entre líneas tolerantes y susceptibles fueron importantes en 2017.
Cuadro 4 Valores medios de AUDPC, RAUDPC, y RaRAUDPC de las líneas de jitomate evaluadas en el campo temporada 2017 en Chapingo, México.
| Línea | AUDPC | Dev St | RAUDPC | Dev St | RaRAUDPC | Std. Dev. |
|---|---|---|---|---|---|---|
| Río grande (RG) | 1752.9a | 156.7 | 0.53a | 0.05 | 7.1a | 0.64 |
| Línea 3 | 1819.3a | 106.6 | 0.55a | 0.03 | 7.4a | 0.44 |
| Línea 51 | 1866.4a | 214.3 | 0.57a | 0.06 | 7.6a | 0.88 |
| Línea 72 | 1932.3a | 151.1 | 0.59a | 0.05 | 7.9a | 0.62 |
| Línea 92 | 1956.0a | 86.5 | 0.59a | 0.03 | 8.0a | 0.35 |
| Línea 43 | 1920.0a | 223.7 | 0.58a | 0.07 | 7.8a | 0.92 |
| Línea 77 | 1775.3 | 146.4 | 0.54a | 0.04 | 7.2a | 0.60 |
| Media | 1860.3 | 0.56 | 7.6 | |||
| DSM | 372.4 | 0.1 | 1.5 | |||
| CV | 8.7 | 8.7 | 8.7 | |||
| R2 | 0.2 | 0.2 | 0.2 | |||
* Los promedios con letras similares no son significativamente diferentes (Waller- Prueba t de relación K de Duncan, p<0,05) DMS diferencia significativa mínima.
Cuadro 5. Valores medios de AUDPC, RAUDPC, y RaRAUDPC de las líneas de jitomate evealuadas en el campo temporada 2018 en Chapingo, México.
| Línea | AUDPC | Dev St | RAUDPC | Dev St | RaRAUDPC | Std.Dev. |
|---|---|---|---|---|---|---|
| Río grande (RG) | 1333.2a | 79.8 | 0.4a | 0.03 | 7.4a | 0.45 |
| Línea 3 | 1380.3a | 90.4 | 0.4a | 0.03 | 7.7a | 0.51 |
| Línea 51 | 1410.9a | 47.4 | 0.5a | 0.02 | 7.9a | 0.27 |
| Línea 72 | 1354.5a | 140.6 | 0.4a | 0.05 | 7.6a | 0.79 |
| Línea 92 | 1330.0a | 48.9 | 0.4a | 0.02 | 7.4a | 0.27 |
| Línea 43 | 1426.2a | 115.8 | 0.5a | 0.04 | 8.0a | 0.65 |
| Línea 77 | 1362.8a | 59.9 | 0.4a | 0.02 | 7.6a | 0.34 |
| Media | 1371.1 | 0.4 | 7.7 | |||
| DSM | 205.6 | 0.07 | 1.1 | |||
| CV | 6.5 | 6.5 | 6.5 | |||
| R2 | 0.1 | 0.1 | 0.1 | |||
* Los promedios con letras similares no son significativamente diferentes (Waller- Prueba t de relación K de Duncan, p<0,05) DMS diferencia significativa mínima.
Cuadro 6. Factores y niveles de condición de los parámetros utilizados para el modelo epidemiológico LATEBLIGHT para simular y predecir las epidemias de tizón tardío en Chapingo.
| Factory | Nivel de condiciones para | ||
|---|---|---|---|
| Phytophthora infestansz | |||
| 0 | 0.5 | 1 | |
| LP | 2.8 | 3.1 | 3.4 |
| LGR | 4.1 | 3.7 | 3.4 |
| SR | 2.9 | 2 | 1.2 |
| IE | 1 | 0.9 | 0.8 |
| IL | 500 | 50 | 5 |
| DOI | 1 | 11 | 21 |
y LP Tiempo de latencia mínimo (días), LGR proporción máxima de desarrollo de la lesión (m día-1, × 10−3), SR proporción máxima de esporulación (esporangio m−2 día−1, × 108), factor de eficiencia de infección IE ( La proporción de esporangios y zoosporas que infectan al huésped con un máximo de 0,1 infecciones por esporangio por día y de 0,01 infecciones por zoosporas germinadas por día), la lesión inicial de IL (lesiones m−2 del suelo) no se aplica en condiciones de infección natural y el día de la inoculación DOI (días después de la emergencia) no se aplica en condiciones de infección natural.
z 0 = favorable (cultivar Tomasa susceptible); 0.5 = intermedio (cultivar intermedio Yungay); 1 = menos favorable (cultivar resistente Amarilis) Andrade-Piedra et al. 2005c.
RaRAUDPC se utilizó para generar una escala de susceptibilidad (1-8) del grado de resistencia o susceptibilidad de las líneas en dos ciclos de cultivo (Yuen y Forbes, 2009). Este sistema escalar es práctico y fácil de usar porque permite la mínima diferencia entre los promedios observados de 7.6 vs 7.7 en el ciclo 2017 y 2018, respectivamente. Este valor se convierte en el valor máximo (≤ 8) para las líneas susceptibles e intermedias y el valor mínimo (<1) para el cultivar resistente (Cuadros 4 y 5). El valor mínimo no se observó en los dos ciclos de cultivo debido a que no existieron diferencias estadísticas en RaRAUDPC de las líneas por su valor alto de AUDPC.
Escenario simulado. El modelo “LATEBLIGHT” involucró el uso de Procedimientos específicos, incluyendo magnitudes de parámetros y variables epidemiológicas (Cuadro 6), parámetros meteorológicos del entorno específico, y la estimación del inóculo inicial del patógeno (Andrade-Piedra et al., 2005c). Estos procedimientos hicieron posible obtener valores de AUDPC observados y simulados, además de predecir epidemias en condiciones naturales de infección en Chapingo. En 2017, se utilizó el modelo para obtener valores de AUDPC simulado y observado para cada tratamiento (Figura 3). Además, el modelo predijo la incidencia de epidemias días antes del inicio de la infección natural en el campo, lo que permite la implementación de medidas de control integral. Por lo tanto, al predecir la epidemia, el modelo incluye magnitudes de variables y parámetros epidemiológicos (Cuadro 6) además de los parámetros meteorológicos de T entre 10.6 y 15.4 °C, HR entre 80 y 100% para períodos continuos de 9 a 13 h durante la noche, madrugada y mañana.
Sin embargo, la magnitud del umbral de humedad relativa no se mantuvo fijo en 85 o 90% en este modelo, debido a la variación causada por las condiciones ambientales de Chapingo. La HR requiere calibración dependiendo del grado de susceptibilidad de los cultivares; específicamente, al 80% para cultivares susceptibles, y al 88% para cultivares resistentes. El límite superior del 88% existe porque la humedad no supera el 90% dentro del dosel de la hoja (Andrade-Piedra et al., 2005b). Las líneas susceptibles tienen valores de AUDPC > 1956 comparado a la línea tolerante (AUDPC < 1753) (Cuadro 7).
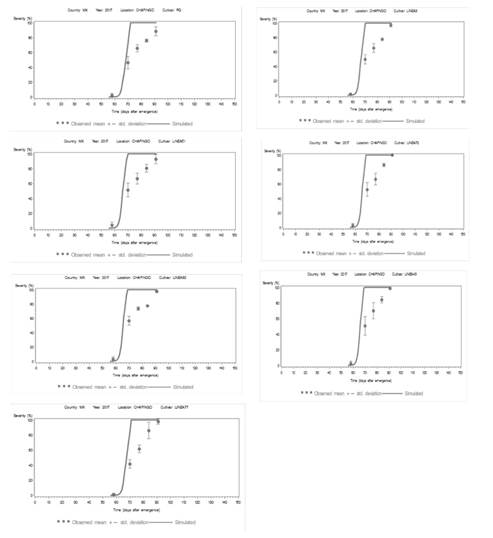
Figura 3. Área bajo la curva de progreso de la enfermedad (AUDPC) de los datos observados (círculos) y simulados (línea continua) de siete epidemias de tizón tardío en condiciones de campo de Chapingo, Edo. de México en 2017. La curva de progreso simulado se desarrolló utilizando la versión LB2004 de “LATEBLIGHT”. Las líneas verticales representan la desviación estándar de la media para la gravedad del tizón tardío observado.
Durante el ciclo 2018, la aplicación del algoritmo del modelo generó valores de AUDPC simulados cercanos a los observados en el campo (Figura 4), el modelo predijo la incidencia de epidemias dos días antes de la infección natural observada en el campo. La predicción epidémica se desarrolló con las magnitudes de las variables y parámetros epidemiológicos (Cuadro 6), así como los parámetros meteorológicos de T entre 10.6 y 15.4 °C, HR entre 80 y 100% para una longitud continúa de 5 a 13.5 h en la noche y parte del amanecer. Sin embargo, la magnitud umbral de la humedad relativa no se mantuvo fija en 85 o 90%, según el modelo, por la variación causada por las condiciones ambientales en Chapingo. Como resultado, la HR se calibró a 80-85% para las líneas susceptibles y 86% para las líneas tolerantes. Este valor se utilizó porque la HR máxima no superaba el 90% de humedad dentro del dosel foliar (Andrade-Piedra et al., 2005b). El AUDPC distingue a las líneas susceptibles (AUDPC > 1426) de las líneas tolerantes (AUDPC < 1333) (Cuadro 8).
Cuadro 7 Parámetros y variables LATEBLIGHT para AUDPC en el ciclo de cosecha 2017.
| Líneaw | AUDPCx | LB2004 Variablesy | Eval0 | Horario | Variables meteorológicasz | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Obs. | Sim. | HR | Mic | t0 | t1 | H_h | T | HR | |||
| Línea92 S | 1956.1 | 2559 | 80 | 209 | 50-51 | 58 | Ago. 06 | 19:00-04:00 | 9 | 10.1-17 | 84.4-100 |
| Línea72 I | 1932.3 | 2531 | 80 | 190 | 50-52 | 58 | Ago. 06 | 19:00-04:00 | 9 | 10.1-17 | 84.4-100 |
| Línea43 I | 1920 | 2523 | 80 | 160 | 50-53 | 58 | Ago. 06 | 19:00-04:00 | 9 | 10.1-17 | 84.4-100 |
| Línea51 I | 1866.4 | 2494 | 80 | 130 | 50-54 | 58 | Ago. 06 | 19:00-04:00 | 9 | 10.1-17 | 84.4-100 |
| Línea03 I | 1819.4 | 2446 | 80 | 98 | 50-55 | 58 | Ago. 06 | 19:00-04:00 | 9 | 10.1-17 | 84.4-100 |
| Línea77 I | 1775.3 | 2363 | 80 | 59 | 50-56 | 58 | Ago. 06 | 19:00-04:00 | 9 | 10.1-17 | 84.4-100 |
| RG R | 1753 | 2285 | 80 | 37 | 50-57 | 58 | Ago. 06 | 19:00-04:00 | 9 | 10.1-17 | 84.4-100 |
w S susceptible, I intermedio, R resistente v..
x obs = AUDPC observado; sim = AUDPC simulado.
y HR = RH_threshold = umbral de humedad relativa para estimar el período diario de humedad de la hoja (%); Mic = InMicCol = microcolonias iniciales utilizadas en la simulación, t0 = última evaluación de lesiones no observadas de P. infestans (días después de la emergencia) para estimar el inóculo inicial, t1 = primera evaluación de lesiones observadas de P. infestans (días después de la emergencia) para estimar el inóculo inicial; Eval0 = fecha de evaluación de la gravedad del patógeno sin lesiones observadas en el campo
z T Rango diario de la temperatura mínima del aire (°C), H_h período diario cuando la humedad relativa> 80%, calculado de 24:00 a 24:00, RH rango diario de humedad relativa ≥80%..
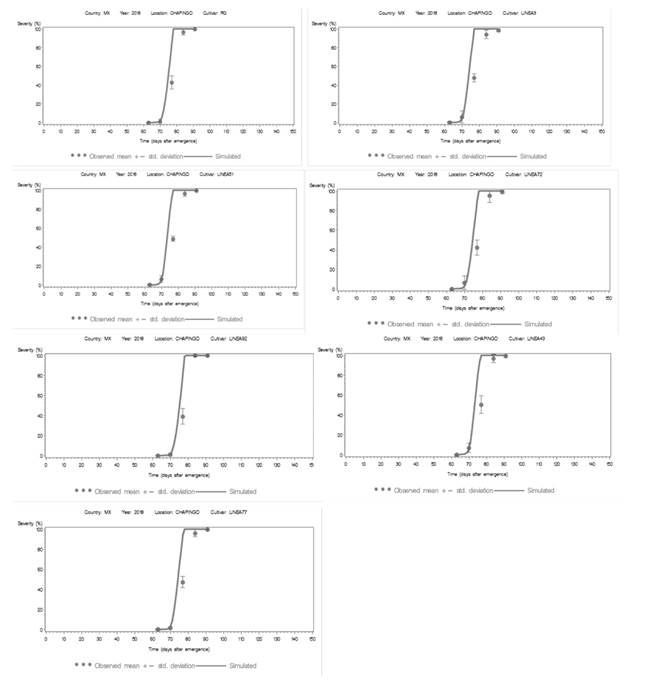
Figura 4. Área bajo la curva de progreso de la enfermedad (AUDPC) de los datos observados (círculos) y simulados (línea continua) de siete epidemias de tizón tardío en condiciones de campo de Chapingo, Edo. de México en 2018. La curva de progreso simulado se desarrolló utilizando la versión LB2004 de “LATEBLIGHT”. Las líneas verticales representan la desviación estándar de la media para la gravedad del tizón tardío observado.
Los escenarios esperados se basaron en (1) un mejor modelo de regresión lineal múltiple; (2) Un gráfico de tendencia, curvas; y (3) criterio de selección 80% ≤RH≤90% de T y RH. Estos criterios llevaron a la identificación de cinco escenarios favorables para epidemias de P. infestans con cuatro a diez ciclos de infección por P. infestans con una duración de 5 a 12 h para las condiciones ambientales en Chapingo. Las tasas de infección observadas durante los ciclos de cultivo 2017 y 2018 confirmaron que la incidencia real de condiciones favorables de P. infestans fue influenciado por condiciones ambientales (T y HR). Se encontró que la infección por P. infestans en el ciclo observado en campo en 2017 coincidió con los escenarios esperados IV-4 y III-1, y en el 2018 coincidió con el escenario V-2. El modelo del experimento de campo es válido con un coeficiente de variación de <16%.
Cuadro 8 Parámetros y variables LATEBLIGHT para AUDPC en el ciclo de cosecha 2018.
| Líneaw | AUDPCx | LB2004 Variablesy | Eval0 | Horario | Variables meteorológicasz | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Obs. | Sim. | HR | Mic | t0 | t1 | H_h | T | HR | |||
| Línea 43 | 1426.2 | 1789 | 80 | 77 | 55-56 | 63 | Ago. 16 | 22:00-08:00 | 10 | 12.8-15.4 | 82.1-96.3 |
| Línea 51 | 1410.9 | 1773 | 80 | 69 | 55-56 | 63 | Ago. 16 | 22:00-08:00 | 10 | 12.8-15.4 | 82.1-96.3 |
| Línea 03 | 1380.3 | 1736 | 80 | 53 | 55-56 | 63 | Ago. 16 | 22:00-08:00 | 10 | 12.8-15.4 | 82.1-96.3 |
| Línea 77 | 1362.8 | 1697 | 80 | 41 | 55-56 | 63 | Ago. 16 | 22:00-08:00 | 10 | 12.8-15.4 | 82.1-96.3 |
| Línea 72 | 1354.5 | 1670 | 80 | 34 | 55-56 | 63 | Ago. 16 | 22:00-08:00 | 10 | 12.8-15.4 | 82.1-96.3 |
| RG | 1333.2 | 1637 | 80 | 27 | 55-56 | 63 | Ago. 16 | 22:00-08:00 | 10 | 12.8-15.4 | 82.1-96.3 |
| Línea 92 | 1330 | 1615 | 80 | 23 | 55-56 | 63 | Ago. 16 | 22:00-08:00 | 10 | 12.8-15.4 | 82.1-96.3 |
w S susceptible, yo intermedio, R resistente.
x Obs = AUDPC observado; sim = AUDPC simulado.
y HR = RH_threshold = umbral de humedad relativa para estimar el período diario de humedad de la hoja (%); Mic = InMicCol = microcolonias iniciales utilizadas en la simulación, t0 = última evaluación de lesiones no observadas de P. infestans (días después de la emergencia) para estimar el inóculo inicial, t1 = primera evaluación de lesiones observadas de P. infestans (días después de la emergencia) para estimar el inóculo inicial; Eval0 = fecha de evaluación de la gravedad del patógeno sin lesiones observadas en el campo.
z T Rango diario de la temperatura mínima del aire (°C), H_h período diario cuando la humedad relativa> 80%, calculado de 24:00 a 24:00, RH rango diario de humedad relativa ≥80%.
Conclusiones
El número de ciclos de infección del escenario esperado se relacionó de manera directa con la severidad que presentó P. infestans en los escenarios observados. Entre mayor es el número de ciclos de infección asociados con las condiciones ambientales favorables, mayor es la severidad de tizón tardío. Los escenarios simulados con el modelo LATEBLIGHT predijeron la severidad de los escenarios observados con dos días de antelación. De esta manera se obtuvo que entre los escenarios esperados, observados y simulados existe una estrecha relación para predecir epidemias de P. infestans en el cultivo de jitomate en Chapingo. En conclusión, el modelo demostró ser capaz de predecir epidemias de la infección en el campo, facilitando la aplicación oportuna de los métodos de control integral, lo que generaría beneficios económicos y reducir los impactos ambientales de agroquímicos.

