











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkEn México se cultivan 18,543.5 ha de jamaica (Hibiscus sabdariffa) y se producen 7,683.2 t de cálices deshidratados, con un valor de la producción superior a $ 293 millones de pesos (SIAP, 2018). En dicho país, los estados productores son: Guerrero (14,079 ha), Michoacán (1,714 ha) y Oaxaca (1,605 ha), quienes aportan el 93%, de la producción nacional (SIAP, 2018). En Guerrero, los principales municipios productores son: Ayutla de Los Libres, Tecoanapa, Acapulco de Juárez, San Luis Acatlán, Juan R. Escudero y San Marcos, los que aportaron 64% de la producción nacional en 2018. El sistema de producción maíz-jamaica constituye el eje de la economía de las familias campesinas, el maíz se usa como alimento básico de autoconsumo y la jamaica como cultivo comercial para obtención de ingresos económicos. La enfermedad “manchado de cáliz” (MCJ) causada por Corynespora cassiicola, es considerara una de las más severas en Guerrero, afecta el 60% de la superficie sembrada (Cesavegro, 2018).
No obstante, se reportan en Guerrero, a los hongos Coniellla musaiaensis var., hibisci, Phomopsis sp., Corynespora cassiicola y Lasiodiplodia theobromae asociados al MCJ, provocando manchas muy similares en el cáliz de color negro, ligeramente hundidas de forma irregular, conforme la infección avanza incrementan en tamaño y número e invaden el cáliz (Martínez, 2010). Correa et al. (2011), determinaron que el manchado acuoso del follaje de jamaica en el estado de Tabasco, fue inducido por Pilidiella diplodiella (sin. Phoma diplodiella) donde causa síntomas con manchas foliares circulares e irregulares color marrón claro, presencia de necrosis con apariencia acuosa en toda la lámina foliar. Conforme avanza la necrosis, se muestran anillos concéntricos marrón claro de consistencia suave; así como, pequeños cuerpos (picnidios) de color marrón oscuro agrupados que sobresalen de la cutícula de la hoja. La necrosis alcanza el peciolo de la hoja, esta se debilita y se desprende de la planta. Las manchas inician en las hojas inferiores y avanzan a las hojas superiores, y cuando el nivel de daño es severo, ocurre defoliación. Mientras que Ortega-Acosta et al. (2015) reporta a Corynespora cassiicola como agente causal del MCJ en Guerrero, el cual causa manchas circulares irregulares en las hojas con centros de color pajizo, bordes negros y anillos de color purpura, al crecer se unen en grandes áreas necróticas; en los cálices se observan puntos hundidos necróticos, circulares a irregulares, causando un tizón en el ápice que se extiende a la base del cáliz. Barrón-Coronado et al. (2019), en muestras procedentes de Ayutla, reportan la patogenicidad de Coniella javanica como el causante del tizón de hojas y cálices, describiendo síntomas con manchas irregulares, de color marrón claro, que luego aumentaron de tamaño y causaron una necrosis con la presencia de picnidios, en etapas avanzadas de la enfermedad, se observó un marchitamiento general de la planta.
El conocimiento de la disponibilidad y dispersión de inóculo de los fitopatógenos, de los factores ambientales que favorecen la liberación de esporas y el desarrollo de la planta, son aspectos fundamentales para determinar los períodos de riesgo de infección, que permita implementar medidas preventivas de control. Por lo que, el objetivo de este trabajo fue estudiar las relaciones entre fluctuación estacional de esporas de Corynespora y Coniella hongos patógenos al manchado de cálices y al tizón de hojas y cálices, las condiciones de clima y la severidad del manchado de cálices (MCJ) en Ayutla, Gro. México.
Materiales Y Métodos
Área de estudio. El trabajo se realizó en una plantación de jamaica localizada en Pozolapa, Mpio. de Ayutla de los Libres, Guerrero, México (16° 53’ 35.2’’ N, 99° 05’ 59.7’’ O), a una altitud de 275 msnm, con antecedentes de alta incidencia de la mancha acuosa de hojas, tizón cálices y MCJ, durante dos ciclos de producción en primavera-verano 2017 y 2018. El municipio presenta clima cálido subhúmedo (Aw1) (García, 2004). La plantación bajo estudio fue de una hectárea. La siembra se realizó el 04 y 10 de julio de 2017 y 2018 respectivamente, con espeque, con cuatro a seis semillas por mata del genotipo denominado “Coneja”, a distancia de 1.0 m entre surcos y matas. Cuando las plantas tuvieron 10 cm de altura, se realizó un raleo, para dejar dos a tres plantas por mata, con una densidad aproximada de 25,000 plantas ha-1. La fertilización se realizó con la fórmula 45-30-20 en dos aplicaciones (Alejo, 2016). En la plantación no se aplicaron agroquímicos para el control de plagas y enfermedades.
Identificación de hongos. En 2017 se colectaron dos hojas y dos cálices con manchas irregulares con puntos hundidos necróticos de la parte intermedia de 14 plantas, los cuales se cortaron en 56 fragmentos de 2.0 mm, se desinfectó la superficie con hipoclorito de sodio al 1.0% durante 30 s y se enjuagaron en tres cambios sucesivos de agua destilada estéril y se sembraron en medio de cultivo papa-dextrosa-agar (PDA), y se incubó durante seis días a 26 °C por 12 horas de fotoperiodo. De estas colonias en crecimiento se obtuvieron 28 subcultivos y se realizaron cultivos monoconidiales por la técnica de rayado en agar al 2%. Luego de seis días se procedió a identificar hasta nivel de género las colonias fungosas obtenidas en cada muestra, empleando las claves Barnett y Hunter (1998).
De los aislamientos realizados se seleccionaron dos colonias de Phoma, se purificaron mediante transferencias de puntas de hifas. Para la extracción de ADN se realizaron transferencias a medio líquido papa-dextrosa, se incubaron por 24 horas en agitación y se colectó el micelio. La extracción del ADN se realizó mediante el método de Dellaporta et al. (1983), su concentración, calidad y pureza se cuantificó en un nanodrop One marca Thermo Scientific®, colocando 2 uL de ADN por muestra. Para confirmar la identificación de las cepas, se amplificaron las regiones del espacio transcrito interno (ITS) usando los iniciadores ITS1 / ITS4 (White et al., 1990), las PCR se realizaron con el kit de QIAGEN Top Taq Master mix, en el termociclador Bio-RadT-100®. Las condiciones para la amplificación fueron una etapa inicial de pre-incubación por dos min a 95 °C seguida de 35 ciclos de desnaturalización por 30 s a 95 °C, hibridación por 30 s a 55 °C y extensión por un minuto a 72 °C y una etapa final de extensión durante 10 min a 72 °C (White et al., 1990). Los fragmentos amplificados fueron visualizados mediante gel de agarosa en TAE al 1.0%, teñidos con GelRed. El fragmento de ADN amplificado se envió a secuenciar en ambas direcciones con el método de didesoxinucleótidos marcados en el secuenciador 3130 Genetic Analyzer (Applied Biosystems®) en el Laboratorio Nacional de Biotecnología Agrícola, Médica y Ambiental del Instituto Potosino de Investigación Científica y Tecnológica, A.C. (IPICYT). Las secuencias obtenidas se compararon para su identificación con una búsqueda de secuencias en las bases de datos del banco de genes del National Center for Biotechnology Information (NCBI) utilizando el algoritmo BLAST (Altschul et al., 1997; Zhang et al., 2000).
Para confirmar la patogenicidad de dos cepas seleccionadas, se utilizaron plantas sanas de jamaica susceptibles de los genotipos Ayutla y Tecoanapa. La cepa de C. cassiicola proveniente de H. sabdariffa (MF000878.1) fue obtenida de la colección de micología del Colegio de Postgraduados México (Dr. Javier Hernández-Morales) y la segunda cepa, identificada por morfología como Phoma, seleccionada para su identificación con marcadores ITS. Cada cepa se consideró un tratamiento y la unidad experimental fue una planta. Seis plantas por genotipo fueron inoculadas con C. cassiicola y otras seis por genotipo con Phoma y se usaron tres plantas por genotipo como testigo (aspersión con agua destilada estéril) de 28 días de edad en un bioespacio ubicado en el Campo Experimental Iguala en 2017 (28 °C y 66% de humedad relativa). La inoculación se realizó rociando las hojas sanas con una suspensión de esporas (6x107 esporas/mL para C. cassiicola y 1x107 esporas/mL para Phoma. Posteriormente, las plantas se cubrieron durante tres días con una bolsa plástica, desinfectada con alcohol, para mantener una temperatura de 28.9 °C y 88% de humedad relativa, se evaluaron las 12 primeras hojas para determinar la incidencia de los síntomas. Una segunda inoculación se realizó a los 77 días de edad de las plantas, la temperatura fue 27.3 °C y 70% humedad relativa, los botones de flor fueron asperjados con una suspensión de esporas (6x107 esporas/mL para C. cassiicola y 10x107 esporas/mL para Phoma) y el tratamiento testigo se asperjó con agua destilada estéril, se evaluaron los tres primeros cálices por planta. La incidencia de la enfermedad se calculó dividiendo el número de hojas y cálices con síntomas entre el número total de hojas y cálices multiplicado por 100. Las evaluaciones en hojas y cálices se hicieron a los 3, 8 y 15 días después de la inoculación. Una hoja y cáliz por planta fue sembrado en PDA por tratamiento con la metodología anteriormente descrita y se volvió a aislar de los tejidos con manchas, las colonias y sus estructuras reproductivas, las que se compararon con las características de las colonias inoculadas originalmente.
Muestreo de esporas. En 2017 y 2018, durante julio a noviembre, se capturó y se contaron las esporas presentes en el aire, en la plantación experimental, se utilizó una trampa volumétrica tipo Burckard, la que trabaja mediante un proceso de succión de aire continuo, con un registro de siete días (Gadoury y MacHardy, 1983). La trampa de esporas se colocó en el centro de la plantación, a 1.7 m de altura por encima del dosel de la jamaica. Las esporas succionadas fueron impactadas sobre un tambor cilíndrico, cubierto por una cinta Janel transparente recubierta con vaselina, la cinta fue cortada en secciones de 39.5 mm correspondientes a cada período de 24 horas y montada sobre un portaobjetos. El conteo de esporas fue en tres transectos a 400x de aumento, calculando el promedio observado por 24 h. Cuando menos de cinco esporas fueron observadas, otros tres transectos fueron contados y se calculó el promedio; para reportar la concentración de esporas acumuladas en siete días. El monitoreo de esporas se realizó durante 133 y 122 días en 2017 y 2018 respectivamente, en las etapas fenológicas de crecimiento vegetal, desarrollo de yemas florales, fructificación y previo a cosecha.
Mediciones meteorológicas. Los factores climáticos de precipitación, temperatura, humedad relativa, radiación global y velocidad del viento se registraron en una estación meteorológica Davis Vantage PRO2, desde julio a diciembre, en 2017 y 2018 respectivamente. Con los registros radiación global se estimó el número de horas con luz solar por día.
Monitoreo de la severidad de la enfermedad. El nivel de daño de la enfermedad se determinó en 25 plantas seleccionadas al azar, en cada una se seleccionó una hoja y cáliz por rama de la base, intermedia y superior de la planta por punto cardinal, contabilizando 12 hojas y cálices por planta y con una escala diagramática de Ortega-Acosta et al. (2016), con rango de 0 al 5, en hoja 0=sano (0% de daño) y 5=daño severo (>57% de daño); en cáliz 0=sano (0% de daño) y 5=daño severo (>77% de daño) para determinar la severidad. El periodo de evaluación para hojas fue en septiembre previo a la emergencia de flores y para cálices en noviembre antes de la cosecha de 2017 y 2018.
Análisis estadístico. Los registros de captura de esporas se agruparon en períodos de siete días y se analizaron con estadísticas descriptivas y la correlación de Spearman (SAS, 2010) entre densidad de esporas acumuladas semanalmente y datos de clima. También se utilizó la correlación de Spearman para relacionar la cantidad de esporas acumuladas semanalmente con severidad de la enfermedad en hojas y cálices.
Resultados
Identificación de hongos. Un crecimiento de micelio color blanco que se tornó grisáceo, a los ocho días estas colonias formaron conidiómatas picnidiales de color hialino y posterior color marrón oscuro de forma redonda cubriendo la caja Petri. Los conidióforos muy densos, delgados, simples y a veces ramificados. Las esporas inmaduras hialinas unicelulares, elípticos, posteriormente color marrón claro, lisas, elipsoidales, de 10-16.2x 5-7 µm características que los ubican dentro del género Phoma (Barnett y Hunter (1998). Dos aislados representativos se depositó en la Colección de Cultivos de Hongos Fitopatógenos en el Laboratorio de Fitopatología, Campo Experimental Centro de Chiapas, CIRPAS-INIFAP, como Gro-1701 y Gro-1702. La secuenciación de ADN de ésta cepa tuvo 99% de similitud con Coniella diplodiella (=Phoma.diplodiella) (KC771899.1).
El segundo crecimiento de micelio septado con conidióforos cilíndricos cortos y conidios terminales individuales o en cadenas cortas con formación acropetala, color café, multicelulares, con exospora gruesa incolora y una prominente cicatriz oscura, características que coinciden con el género Corynespora (Barnett y Hunter, 1998). El tercer crecimiento micelial fue rápido y abundante, en 1 a 3 días, de color gris a oliváceo, conidiomas picnidiales se observaron después de 13 dias, de color negro, obpiriformes, ostiolados, con conidios hialinos inmaduros a los 17 días, elipsoidales a subovoides, a los 22 días se observaron los conidios maduros de color café oscuro, elipsoidales a ovoides y con estriaciones longitudinales irregulares características morfológicas que coinciden con el género Lasiodiplodia (Sutton, 1980; Barnett y Hunter, 1998).
De tejido enfermo, en hojas, los hongos Coniella sp. y Corynespora sp. mostraron la más alta frecuencia de aislamientos con 71.4 y 21.4% respectivamente y en cálices, ambos hongos con 57.1 y 35.7% respectivamente (Cuadro 1). El de menor aislamiento fue Lasiodiplodia sp. se aisló con baja frecuencia en hojas y cálices de 7.1%.
El hongo C. diplodiella causó los síntomas típicos del MCJ en los genotipos Ayutla y Tecoanapa, con 88.9 y 22.2% de síntomas en hojas y cáliz de las plantas respectivamente. El hongo C. cassiicola también causó síntomas similares de MCJ en ambos genotipos, con 56.3 y 25% de síntomas en hojas y cálices. Con ambos hongos, los síntomas en hojas iniciaron con manchas irregulares y la formación de un halo rojo y centro necrótico de 1-3 mm, a las 72 y 192 h después de la inoculación con C. diplodiella y C. cassiicola respectivamente. En cáliz se presentaron las manchas de color negro ligeramente hundidas, de forma irregular de aproximadamente 1-2 mm de diámetro a los 53 y 91 días después de la segunda inoculación con C. diplodiella y C. cassiicola respectivamente. Sin embargo, en hojas, el aislamiento de C. diplodiella fue más agresivo al causar mayor incidencia en el genotipo Tecoanapa y Ayutla, mientras que C. cassiicola tuvo una menor incidencia e incluso hubo tres plantas del genotipo Ayutla que no mostraron síntomas. En contraste, en el cáliz la incidencia de la enfermedad fue ligeramente mayor, C. cassiicola con una incidencia del 25.0% y C. diplodiella fue de 22.2%. La infección de la hoja y cáliz fue confirmada con estos aislamientos, volviendo a re-aislar los dos hongos de las manchas típicas.
Muestreo de esporas. Del muestreo de esporas en el aire se identificaron cinco géneros de hongos por sus características morfológicas: Coniella, Corynespora, Lasiodiplodia, Curvularia y Alternaria (Barnett y Hunter 1998). Los géneros Curvularia, Alternaria y Lasiodiplodia tienen habilidades patogénicas reconocidas en numerosos hospedantes. Lasiodiplodia fue reportada en cálices frescos de jamaica por Martínez (2010) mientras que Ruíz (2014) reportó Alternaria en cálices deshidratados. No obstante, en la etiología del manchado de cálices de jamaica, ha sido consistente la presencia y patogenicidad de Pilidella (=Coniella) diplodiella (Correa et al., 2011), Coniella javanica (Barrón et al., 2019) y Corynespora cassiicola (Ortega-Acosta et al., 2015). Por tal razón el estudio se centró a éstos patógenos.
En el Cuadro 2 se presenta la abundancia de esporas desde la siembra a la cosecha. El género Coniella, con 19.3 y 32.3 esporas/semana en 2017 y 2018 respectivamente (con un máximo de 223.6 esporas/semana del 19 al 25 de septiembre 2018). Con una detección de 44.4 y 52.0% para 2017 y 2018 respectivamente y promedio de 48.2% en ambos ciclos. El género Corynespora con menores concentraciones de 3.9 y 5.3 esporas/semana en 2017 y 2018 (con un máximo de 12.4 esporas/semana del 24 al 30 de noviembre 2018). Con una detección de 9.0 y 8.5% en 2017 y 2018 respectivamente y promedio de 8.7% en ambos ciclos. Las esporas de Lasiodiplodia fueron menos abundantes y con bajo porcentaje de detección en ambos ciclos.
Cuadro 1. Frecuencia de aislamientos en hoja y cáliz de jamaica con síntomas de manchado. Pozolapa, Mpio. Ayutla de los libres, Guerrero, México. 2017.
| Tipo de espora | Frecuencia en hoja (%) | Frecuencia en cálices (%) |
|---|---|---|
| Coniella sp. | 71.4 | 57.1 |
| Corynespora sp. | 21.4 | 35.7 |
| Lasiodiplodia sp. | 7.1 | 7.1 |
Mediciones meteorológicas. La precipitación más alta fue de 141.2 mm el 14 de septiembre 2017 y de 71.9 mm el 10 de octubre 2018 (Cuadro 3). Las temperaturas máximas se presentaron en 2018, con una media de 32.7°C mientras que en 2017 fue de 31.9°C. La menor temperatura se presentó en 2018 con 14.9°C y en 2017 fue de 15.8°C. La mayor radiación global que fue de 600.5 W/m2 se presentó el 11 de agosto de 2018 y de 553.7 W/m2 el 31 de julio de 2017. El mayor rango de velocidad del viento alcanzó hasta 3.2 km hr-1 en 2017, mientras que en 2018 fue de 2.7 km hr-1. La media de humedad relativa fue de 86.4 y 84.4% en 2017 y 2018 respectivamente.
Cuadro 2. Abundancia y porcentaje de detección de esporas promedio por semana, en el sistema de producción de jamaica. Pozolapa, Mpio. Ayutla de los Libres, Guerrero, México.
| Hongo | Primer ciclo 2017 | Segundo ciclo 2018 | Promedio | ||
|---|---|---|---|---|---|
| Abundanciaz | % | Abundancia | % | 2017-2018 (%) | |
| Coniella | 19.3 | 44.3 | 32.3 | 52.0 | 48.2 |
| Corynespora | 3.9 | 9.0 | 5.3 | 8.5 | 8.7 |
| Lasiodiplodia | 2.1 | 4.8 | 1.5 | 2.4 | 3.7 |
| Curvularia | 13.2 | 30.4 | 18.9 | 30.3 | 30.3 |
| Alternaria | 5.0 | 11.5 | 4.2 | 6.8 | 9.1 |
| Total | 43.5 | 100 | 62.2 | 100 | 100 |
z Periodo de julio a noviembre
Distribución temporal de inoculo en el aire. La captura de esporas dentro del sitio de estudio en el periodo de julio a noviembre de 2017 y 2018, mostró presencia de Coniella y Corynespora durante todo el ciclo del cultivo (Figura 1). La detección de esporas fue a partir de la emergencia de las primeras hojas verdaderas en las plantas de jamaica y continuó hasta la cosecha, observándose clara variación mensual.
En julio de 2017, a los 10 días después de la siembra, se registró la presencia de esporas de Coniella y Corynespora con un promedio de 20 y 4.5 esporas/semana respectivamente. En el desarrollo vegetativo, ambos hongos, bajaron sus niveles poblacionales. Coniella con 0.7 esporas/semana en promedio, del 25 de agosto al 13 de octubre de 2017 y 0.2 esporas/semana en promedio entre el 29 de agosto al 11 de septiembre de 2018. Corynespora presentó poblaciones un poco más altas con 1.9 esporas/semana en promedio entre el 25 de agosto al 13 de octubre de 2017 y 0.7 esporas/semana en promedio entre el 29 de agosto al 11 de septiembre de 2018. Sin embargo, durante el desarrollo de yemas florales, fructificación y madurez fisiológica de cálices incrementaron sus niveles poblacionales. Coniella con 41.2 esporas/semana en promedio del 13 de octubre al 24 de noviembre de 2017 y 49.5 esporas/semana en promedio desde el 12 de septiembre al 30 de noviembre 2018. Mientras que Corynespora registró poblaciones promedio más bajas con 7.1 esporas/semana del 13 de octubre al 24 de noviembre de 2017 y 7.2 esporas/semana del 12 de septiembre al 30 de noviembre de 2018.
Cuadro 3. Rango y promedio diario de variables climáticas del 01 de julio al 10 de diciembre, de los años 2017 y 2018. Pozolapa, Mpio. Ayutla de los Libres, Guerrero, México.
| Variables climáticas | 2017 | 2018 | ||
|---|---|---|---|---|
| Rango | Media | Rango | Media | |
| Precipitación (mm) | 0-141.2 | 10.4 | 0-71.9 | 4.8 |
| Temperatura máxima (°C) | 22.1-36.8 | 31.9 | 21.5-36.2 | 32.7 |
| Temperatura mínima (°C) | 15.8-22.7 | 20.5 | 14.9-22.4 | 19.5 |
| Temperatura media (°C) | 21.2-28.1 | 24.9 | 20.2-28.7 | 25.8 |
| Radiación global promedio (W/m2) | 40.9-553.2 | 436.1 | 61.4-600.5 | 460.9 |
| Velocidad Viento promedio (Km h-1) | 0.0-3.2 | 1.2 | 0.24-2.7 | 1.4 |
| Humedad Relativa (%) | 52.5-99.4 | 86.4 | 67.8-99.8 | 84.4 |

Figura 1. Distribución semanal de esporas de Coniella y Corynespora presentes en el aire, desde julio hasta noviembre de 2017 y 2018. La fenología de la jamaica se muestra en la parte inferior. Los periodos típicos de siembra, floración y cosecha en Guerrero se indican con líneas negras.
Correlación entre esporas y clima. Las concentraciones de captura de esporas de los hongos fueron correlacionadas con las variables meteorológicas mediante la prueba de Spearman y los valores se presentan en el Cuadro 4. Las poblaciones individuales de esporas de los cinco géneros estudiados y el total de esporas mostraron sensibilidad a las variaciones climáticas. La lluvia indujo un efecto negativo significativo solamente para la concentración de inoculo de Corynespora. La temperatura fue la variable que más favoreció la dinámica de población de los hongos en el aire. La temperatura máxima mostró una correlación positiva significativa con las fluctuaciones poblacionales de inóculo de Coniella y Curvularia y el total de esporas, mientras que Alternaria, presentó correlación negativa. Y la temperatura mínima con una correlación negativa significativa solamente con la dinámica de población de Corynespora. La temperatura diaria promedio manifestó una correlación positiva con la fluctuación de esporas de Coniella y Curvularia y el total de esporas. De la misma manera, la velocidad máxima del viento se correlacionó negativamente con la cantidad de esporas de Corynespora. En este estudio la radiación global fue otra variable con influencia en la dinámica de población de hongos en el aire, la radiación global promedio mostró una influencia positiva sobre la dinámica de población de Coniella y Curvularia y el total de esporas, mientras que el número de horas de radiación solar mostraron una influencia negativa con las esporas de Corynespora.
Cuadro 4. Coeficiente de correlación de Spearman entre concentración de esporas acumulada semanalmente y variables meteorológicas. Ciclos 2017-2018. Pozolapa, Mpio. Ayutla de los Libres, Guerrero, México.
| Hongo | PPy | Tmax | Tmin | Tmed | VVmax | RGprom | HRG |
|---|---|---|---|---|---|---|---|
| Coniella | -0.3009z | 0.3957 | -0.0512 | 0.4976 | -0.1500 | 0.41278 | 0.0002 |
| 0.0837* | 0.0205 | 0.7736 | 0.0027 | 0.3971 | 0.0153 | 0.9991 | |
| Corynespora | -0.4259 | 0.0120 | -0.3631 | 0.0178 | -0.3553 | -0.03883 | -0.5536 |
| 0.0120 | 0.9463 | 0.0348 | 0.9205 | 0.0392 | 0.8274 | 0.0007 | |
| Lasiodiplodia | -0.1513 | 0.1353 | -0.0903 | 0.1047 | 0.2997 | 0.12448 | 0.0940 |
| 0.3930 | 0.4457 | 0.6116 | 0.5555 | 0.0850 | 0.483 | 0.5970 | |
| Curvularia | -0.2682 | 0.3408 | -0.0785 | 0.4318 | -0.1039 | 0.38521 | 0.0104 |
| 0.1251 | 0.0486 | 0.6591 | 0.0108 | 0.5588 | 0.0245 | 0.9533 | |
| Alternaria | -0.0492 | -0.3526 | 0.0019 | -0.2675 | 0.1998 | -0.20753 | 0.0694 |
| 0.7824 | 0.0409 | 0.9916 | 0.1261 | 0.2572 | 0.2389 | 0.6966 | |
| Total de esporas | -0.3184 | 0.3451 | -0.0780 | 0.4521 | -0.1220 | 0.3839 | -0.0108 |
| 0.0665 | 0.0457 | 0.6611 | 0.0073 | 0.4920 | 0.025 | 0.9518 | |
y PP=precipitación (mm); Tmax=temperatura máxima (°C); Tmin=temperatura mínima; Tmed=temperatura promedio; VVmax=velocidad del viento máxima (km hr-1); RGprom=radiación global promedio(W/m2) y HRG=horas de radiación global.
z Coeficiente de correlación; * Nivel de significancia; número de observaciones 34.
Correlación entre severidad de MCJ y esporas de Coniella y Corynespora. En 2017, la severidad del MCJ en las hojas alcanzó un índice de 2.6, en septiembre y la concentración de esporas de Coniella y de Corynespora fue de 121.2 y 9.1 esporas/semana respectivamente, en la última semana de este mes (Figura 2a); en septiembre se presentó la más alta precipitación acumulada de 936.6 mm. La severidad de la enfermedad en los cálices alcanzó un índice de 3.9 en noviembre, la concentración de esporas de Coniella y de Corynespora fueron de 34.7 y 8.3 esporas/semana respectivamente, en la segunda semana de noviembre (Figura 2a); en la cuarta semana de octubre a la tercera semana de noviembre se presentó una precipitación acumulada de 9.7 mm. En 2018, la severidad del MCJ en las hojas fue alta, con 3.2 de la escala, en septiembre y la concentración de esporas de Coniella y de Corynespora fue más alta con 223.6 y 9.1 esporas/semana respectivamente, en la última semana de este mes (Figura 2b); en septiembre se presentó una precipitación acumulada de 48.3 mm. La severidad de la enfermedad en los cálices alcanzó un índice de 3.55 en noviembre, la concentración de esporas de Coniella y de Corynespora fueron de 0.6 y 12.4 esporas/semana respectivamente, en la tercera semana de noviembre (Figura 2b); de la tercera semana de octubre a la primera semana de noviembre se presentaron precipitaciones acumuladas de 261.7 mm.
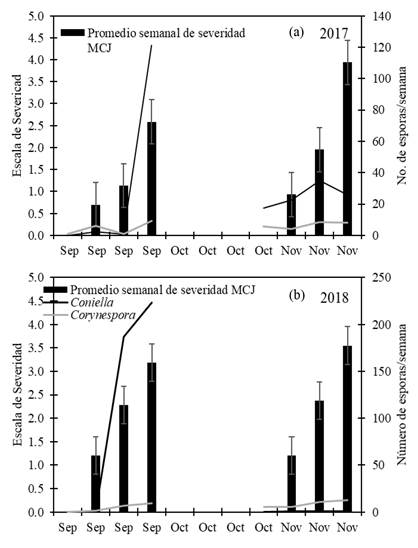
Figura 2. Promedio de severidad semanal del manchado de hoja (septiembre) y cáliz (noviembre) en jamaica (MCJ) estimada con la escala de 0 al 5, donde 0=sano y 5=daño severo en plantas no tratadas en 2017 (a) y 2018 (b). Número de esporas por semana de Coniella y Corynespora atrapadas en la parcela por una trampa de esporas volumétrica de Burkard. Las barras verticales representan errores estándar.
Cuadro 5. Correlación de Spearman entre severidad del manchado del cáliz de jamaica y la concentración de esporas acumuladas semanalmente. Pozolapa, Mpio. de Ayutla de los Libre, Guerrero, México.
| Hongo | Estratos de la plantax | ||
|---|---|---|---|
| Base | Medio | Superior | |
| Coniella | 0.56423y | 0.51853 | 0.50442 |
| 0.0228* | 0.0841 | 0.0945 | |
| 16z | 12 | 12 | |
| Corynespora | 0.76691 | 0.70424 | 0.66199 |
| 0.0005 | 0.0106 | 0.019 | |
| 16 | 12 | 12 | |
| Total de esporas | 0.68794 | 0.61269 | 0.58452 |
| 0.0032 | 0.0342 | 0.0459 | |
| 16 | 12 | 12 | |
x Base=evaluación de severidad en hojas en septiembre; Medio y Superior=evaluación severidad en cálices en noviembre.
y Coeficiente de correlación; * Nivel de significancia y z Número de observaciones.
Las variaciones de severidad de la enfermedad MCJ mostraron alta correlación con las fluctuaciones de población de esporas de Coniella y Corynespora y en el total de esporas (Cuadro 5). La correlación entre la severidad de la enfermedad en las hojas del estrato base de la planta (evaluadas en septiembre) y el número acumulado de esporas de Coniella y Corynespora y el total de esporas fue positiva y significativa para ambos años. En el estrato medio y superior de la planta se evaluó la severidad de la enfermedad en cálices y se correlacionó positivamente con las poblaciones de esporas de Corynespora y con el total de esporas en el aire.
Discusión
El estudio sobre dinámica de las esporas presentes en el aire es importante para comprender el patrón de distribución espacial y temporal del inóculo de los patógenos, para proponer estrategias de manejo integrado de las enfermedades.
Identificación de hongos. En este trabajo, el género Coniella (=Pilidiella) fue el más frecuente en el cultivo y abundante en el aire, mientras que Corynespora tuvo una menor frecuencia en el cultivo y en el muestreo del aire. Lasiodiplodia con mucha menor frecuencia y abundancia sobre el cultivo. En cambio, las esporas de Curvularia y Alternaria fueron abundantes en el aire, en el caso de Curvularia ha sido asociada al MCJ (Trujillo-Tapia y Ramírez-Fuentes, 2015). No obstante, el estudio se centró en los géneros Coniella y Corynespora, debido a que han sido reportados consistentemente como agentes patógenos en jamaica con las especies P. (=Coniella) diplodiella, como causante de mancha acuosa en hojas y defoliación de la planta (Correa et al., 2011), C. javanica causante del tizón de las hojas, cálices y un marchitamiento general de la planta (Barrón-Coronado et al., 2019) y Corynespora cassiicola reportado como agente causal del manchado de cáliz con puntos circulares a irregulares hundidos necróticos, causando un tizón del ápice a la base del cáliz y en hojas manchas circulares a irregulares con centros de color pajizo, bordes negros y anillos púrpuras que al unirse forman áreas necróticas (Ortega-Acosta et al., 2015). Las pruebas de patogenicidad realizadas con C. diplodiella y C. cassiicola confirmaron sintomatología muy similar entre ambos hongos.
Clima y patógenos. Las esporas de Coniella y Corynespora fueron atrapadas sobre el cultivo durante todo el ciclo productivo, desde los primeros 10 días después de la siembra. Las primeras lluvias provocaron la germinación de semilla que quedo en el terreno del ciclo anterior, lo que permite una buena disponibilidad del tejido hospedante; por otro lado, Hernández-Morales et al. (2018) menciona que Corynespora cassiicola tiene otros hospedantes alternos como malezas y plantas cultivadas del área de producción, lo cual puede explicar los niveles de inóculo desde el inicio del cultivo. No obstante, las mayores poblaciones de esporas se presentaron durante el desarrollo de las yemas florales a cosecha, en los meses de octubre y noviembre. El inicio de emergencia de las flores se presenta a finales de septiembre, y coincide con días cortos de menos de 12 horas de exposición a la luz, lo cual ha sido bien documentado por Muslihatinn y Daesusi (2014). La concentración de esporas en el aire sobre el cultivo estuvo estrechamente relacionada con la precipitación, temperatura, velocidad del viento y radiación global ambientales previas a su captura. Sin embargo, se encontraron correlaciones negativas entre la lluvia y la concentración de esporas de Corynespora, al parecer cuando ocurrieron lluvias torrenciales > 138 mm en el mes de septiembre 2017, la densidad de esporas en el aire se redujo. Este comportamiento se ha reportado para otros hongos, como Botrytis cinerea, quienes indican que las gotas de lluvia reducen las esporas en el aire (Blanco et al., 2006). Una situación similar reporta Pakpour et al. (2015) en un estudio urbano, donde indican que las concentraciones de esporas en el aire eran mayores cuando llovía poco y con altas temperaturas; las esporas de hongos persisten en el aire durante los períodos secos y la lluvia pueden disminuir la cantidad de esporas presentes en el aire. En contraste Ganthaler y Mayr (2015) indican que las condiciones húmedas del suelo y las hojas que persisten después de un evento de lluvia contribuyen a un aumento en la densidad de esporas de hongos en el aire.
La temperatura máxima y temperatura media diaria correlacionaron positivamente con la concentración de esporas de Coniella, en este estudio, el período con mayores capturas de esporas correspondió con períodos de temperatura entre 30 -33.6 °C. Lo cual no corresponde a lo reportado para otros hongos como Botrytis cinerea por Sosa-Alvarez et al. (1995), quienes informaron que se produce una gran cantidad de esporas a una temperatura cercana a los 15-22 °C después de 7 días de humedad continua. No obstante, la producción y dispersión de las esporas son dos eventos diferentes y es probable que estas temperaturas máximas facilitaron la dispersión en el aire y por ello la mayor captura, puesto que las temperaturas mínimas diurnas de estos mismos días fueron de 19.4-20.4 °C (Cuadro 3 y Figura 1). Esta situación es posible que pueda ser explicada por los resultados encontrados con Botrytis cinerea donde se distinguen dos fases: la primera, hay una liberación de esporas del conidióforo mediada por un mecanismo higroscópico que controla la turgencia del conidióforo y, en segundo lugar, se presenta un episodio de transporte conidial mediado por agentes físicos (viento y las gotas de lluvia), situación que a menudo ocurre temprano en la mañana cuando la humedad relativa cambia y a medida que aumenta la temperatura (Blanco et al., 2006). Al contrario, las poblaciones de Corynespora en este estudio mostraron una correlación significativamente negativa con la temperatura mínima, las mayores poblaciones se presentaron a temperaturas entre 17.2 a 20.7°C, lo cual difiere a lo reportado por Kamei et al. (2018), quienes indican que C. cassiicola provoca el progreso del manchado foliar en tomate a una temperatura óptima entre 24 y 25°C, junto con el 80-85% de humedad relativa. En cambio, Sharma (2017) reportan un rango más amplio de temperaturas de 16-32 °C, donde el alargamiento de la duración de la humedad de la hoja aumenta el número de lesiones de mancha foliar en algodón causado por C. cassiicola a todas las temperaturas probadas de 16, 20, 24, 28 y 32 °C, las mayores lesiones fueron a 28 °C con 48 horas de humedad de la hoja. Los patrones de distribución de esporas dentro del dosel de jamaica rara vez se han estudiado, se necesitan más datos y estudios para comprender completamente la epidemiología del complejo de enfermedades de la jamaica.
La presencia de vientos con velocidades máximas >4.4 km hr-1 y el tiempo con luz solar diario <12 hrs estuvieron correlacionados negativamente con las bajas densidad de poblaciones de esporas de Corynespora en el aire. Mientras que la radiación global solar promedio diaria fue de >249 (W/m2) lo que favoreció el incremento de inóculo de Coniella en el aire y el total de esporas. El efecto de la luz solar sobre la esporulación de los hongos ha sido documentado y estudiado experimentalmente para algunos hongos, el principal efecto fotomorfogenético es a través de la inducción de la esporulación (Ensminger 1993). Sin embargo, las longitudes de onda que inducen la esporulación a cantidades bajas pueden inhibir la esporulación en cantidades más altas (Paul 2000) y estas mismas longitudes de onda tienen efectos estimulantes o inhibidores sobre el desarrollo del conidio y afectan la fotomorfogénesis fúngica (Maddison y Manners 1973).
La variación anual del clima, principalmente en la temperatura y en menor grado las precipitaciones pueden determinar las concentraciones de esporas en el aire. Al parecer en el año con menos precipitación resultó con mayores concentraciones de esporas lo que puede tener implicaciones para el control de la enfermedad y de cambio climático.
Correlación entre severidad de MCJ y esporas de Coniella y Corynespora. Los resultados del presente trabajo mostraron que la interacción entre el hospedante, clima e inóculo, que prevalecieron durante las etapas fenológicas del cultivo de la jamaica, favoreció el desarrollo de la enfermedad.
En el presente estudio hubo correlaciones significativas positivas entre la severidad de MCJ y la concentración de esporas de Coniella y Corynespora y el total de esporas. Los niveles de inóculo de Coniella tuvo una mayor relación con la severidad en hojas y por el contrario Corynespora sus incrementos de inóculo sobre el cultivo se relacionó con la severidad de la enfermedad en el desarrollo de flores. Ambos géneros de hongos han sido reportados como agentes causales de enfermedades en jamaica: Correa et al. (2011) reportó a Pilidiella (=Coniella) diplodiella con síntomas del manchado acuoso de hojas, Barrón-Coronado et al. (2019) a Coniella javanica patógeno del tizón de hojas y cálices, Ortega-Acosta et al. (2015) reportaron a Corynespora cassiicola como patógeno del manchado de hojas y cálices.
Durante la etapa de crecimiento vegetativo, desde la germinación de la semilla a inicio de yemas florales, la presencia de poblaciones de esporas de Coniella, y de una continua presencia de tejido susceptible (hojas jóvenes y maduras), de julio a principios de septiembre, no es limitante en cuanto a disponibilidad de tejido hospedante. No obstante, en septiembre, previo al inicio de la emergencia floral se observó una defoliación importante de hojas afectadas por manchas foliares, aunado a la poca concentración de inoculo en el aire, debido a la disminución de esporas presentes en el aire por las lluvias torrenciales en este mes, el SMN (2019) reporta una precipitación normal en septiembre de >434.8 mm. Cuando se presenta la floración de la jamaica (octubre y noviembre), coincide con la fase de crecimiento acelerado de la enfermedad presente en el follaje joven, con niveles altos de inóculo (Coniella y Corynespora con >100 y 8 esporas/semana respectivamente sobre el cultivo) y con las condiciones atmosféricas idóneas para la epidemia (temperatura media por día 25.3-26.6 °C, radiación promedio de 235.9-317.1, con humedades relativas >84% y con 12-11.2 hrs de radiación solar por día).
Estos resultados pueden tener implicaciones para el control de la enfermedad, donde se han reportado hasta siete aplicaciones de fungicidas (mancozeb, clorotalonil y benomilo alternando con oxicloruro de cobre), programados en prefloración, floración y fructificación, las cuales detiene las infecciones por Corynespora cassiicola, y por consiguiente una menor severidad de la enfermedad y mayor peso fresco y seco de cálices (Ortega-Acosta et al., 2019). Sin embargo, es importante intentar reducir las aplicaciones de fungicidas inoportunas y potencialmente innecesarias, a través de información de muestreo de inóculo transportado por el aire, en tiempo cercano al real, que permita estimar el riesgo de acumulación potencial de esporas y su germinación, lo que ayudaría a ajustar los intervalos de aplicación de fungicidas.
Conclusiones
Las esporas de Coniella y Corynespora fueron los géneros comunes presentes en el aire sobre el cultivo de jamaica. Las variaciones del clima, temperatura y precipitación principalmente, determinan las concentraciones de esporas sobre el cultivo. La severidad del manchado del cáliz estuvo correlacionada positiva y significativamente con el número de esporas de Coniella y Corynespora acumulados semanalmente.

