











 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkEl chile manzano (Capsicum pubescens R y P) es un producto agrícola con altas posibilidades de comercialización a nivel mundial por su demanda para consumo como verdura, o bien por sus propiedades químicas para la obtención de derivados de capsaicina (Pérez-Grajales et al., 2004). El chile manzano fue introducido en México a principios del siglo XX, es el único tipo de chile que se cultiva entre de 1700 y 2500 m; crece como arbusto grande y puede llegar hasta los 3 metros, el fruto es esférico, verde en estado tierno y se torna rojo, amarillo y naranja al madurar. Las principales zonas productoras de chile manzano se encuentran ubicadas en los estados de Michoacán, Puebla, Edo. de México y Veracruz, en menor escala Chiapas y Oaxaca (Espinosa-Torres y Villa-Galindo, 2008). El período de siembra es en los meses de abril y mayo, el consumo es insustituible para las zonas de la sierra norte de Puebla, Toluca y Michoacán (principalmente Morelia), donde forma parte de los hábitos alimenticios. En cuanto a su cosecha a cielo abierto es de 9 a 14 t/ha e invernadero 40 a 70 t (Espinosa-Torres y Ramírez-Abarca, 2016). Sin embargo, el cultivo puede ser afectado por patógenos. Entre las enfermedades que afectan el cultivo se encuentran la marchitez del chile manzano, causada por diversos fitopatógenos, tales como Botrytis, Fusarium spp., Phytium, etc., los cuales ocasionan pérdidas significativas en la producción en varias partes del mundo, incluyendo México. Fusarium sambucinum [teleomorfo Gibberella pulicaris (Fr.) Sacc (Leslie y Sumerell, 2006)] es una de las especies de hongos que causa podredumbre seca o pudrición negra en papa (Solanum tuberosum L.), afectando en campo como en almacenamiento (Hanson et al., 1996; Peters et al., 2008). F. sambucinum ha sido reportado en Europa, Estados Unidos de América y, recientemente, en China afecta a frutos en almacenamiento y que produce micotoxinas, tales como tricotecenos (El-Banna et al., 1984; Altomare et al., 1995; Du et al., 2012). La identificación correcta de un fitopatógeno es esencial para proponer las alternativas más efectivas para su manejo (Agrios, 2005). En este contexto, el objetivo, fue determinar el agente causal de lesiones hundidas en frutos de chile manzano por medio de caracterización morfológica y molecular con factor de elongación (EF-1∞) y pruebas de patogenicidad en frutos.
En noviembre del 2015 se llevó acabo un muestreo dirigido a frutos de chile manzano procedentes del Estado de México, municipio de Coatepec de Harinas en donde se colectaron frutos con síntomas de pudrición y lesiones hundidas. Las muestras se trasladaron en bolsas de plástico (Ziploc®) etiquetadas y colocadas en una hielera al laboratorio de postcosecha del Instituto de Fitosanidad del Colegio de Postgraduados.
Para aislar al agente causal, se desinfestaron fragmentos de 1 cm de la zona de avance de la pudrición causada por hongos, para lo cual se realizaron cortes de tejido de 40 frutos y se utilizó una solución de hipoclorito de sodio al 0.2% durante 2 min, enseguida se lavaron con agua destilada estéril dos veces, se dejaron secar sobre sanitas estériles y se colocaron en cajas Petri con 20 mL de PDA. Las cajas se colocaron en incubación a 27+2 °C, bajo un régimen de 24 h luz; una vez esporulados a los seis días de crecimiento, se realizaron cultivos monospóricos y se transfirieron a cajas de Petri con medio de cultivo PDA. La conservación de los aislados se llevó acabo con la transferencia de 10 discos miceliales (5 mm de diámetro) a tubos criogénicos de 2 mL, con 1.5 mL de glicerina al 20% (v/v) almacenados a -80 °C y en tubos de ensaye con aceite mineral.
La caracterización morfológica se realizó a partir de sus estructuras de reproducción asexual. Para lo cual los aislados fueron transferidos a medio de cultivo PDA, SNA y Clavel-Agar con tres repeticiones para cada una de las colonias fúngicas, posteriormente fueron incubadas a 25± 2 °C en condiciones de luz continua. Se midió el crecimiento micelial cada 48 h hasta que el hongo llenó por completo la caja donde se evaluó el color de la colonia, tipo de micelio, crecimiento del micelio y esporas (conidios, clamidosporas y esporodoquios). Después de 15 días de incubación se hicieron preparaciones en glicerina al 50 % con la finalidad de reconocer las estructuras de los hongos con ayuda de un microscopio compuesto (Velab Ve-B6, México). Se observaron 50 macroconidios y se registró su tamaño, coloración y número de septos con el programa Motic Images plus 2.0 (Group Co., Ltd). La identificación morfológica de los hongos a nivel de género se llevó acabo empleando las claves especializadas de Barnett y Hunter (1998) y para especie de Leslie y Summerell (2006).
Una vez obtenidos los aislados puros de los hongos, se inició la identificación molecular con la extracción de ADN genómico de nueve aislados e identificados morfológicamente como Fusarium para lo cual se siguió el protocolo indicado en el kit de extracción Plant DNeasy® Minikit (Quiagen). Se colocó una muestra de micelio de 0.030 g en tubos Eppendorf macerando con aguja hipodérmica. Se adicionaron 400 μL de Buffer AP1 y 4 μL de RNasa y se agitó. Las muestras fueron incubadas en baño María por 10 min a 65 °C, se agitó por inversión cada 2 min, después se adicionaron a la muestra 120 μL del Buffer AP2, y se incubó por 5 min en hielo. La mezcla se colocó en la columna QIAshredder Mini spin lila y se centrifugó por 2 min a 14000 rpm. Posteriormente se retiró el sobrenadante para colocarlo en un nuevo tubo Eppendorf al que se agregaron 675 μL del Buffer AP3/E. La mezcla fue colocada en un tubo DNeasy Mini spin centrifugándolo a 8000 rpm por 1 min. La columna fue reemplazada por otra suplementaria del protocolo y se agregaron 500 μL del Buffer AW en la membrana del Mini spin DNeasy y se centrifugó a 8000 rpm por un min; nuevamente se cambió la columna con el filtrado, se adicionaron 500 μL del Buffer AW a la membrana y se centrifugó a 14000 rpm por 2 min. Finalmente, se transfirió la columna DNeasy Mini spin a un nuevo tubo Eppendorf y se agregaron 100 μL del Buffer AE. Se incubó por 5 min a temperatura ambiente y se centrifugó a 8000 rpm por 1 min. Los productos de extracción de ADN se enviaron al laboratorio de Macrogen® Inc. (Corea) para su purificación, amplificación y secuenciación. Las secuencias obtenidas se procesaron en el programa BioEdit v7.0.9. Se realizaron las pruebas de PCR Factor de elongación (EF-1∞) con las siguientes condiciones: desnaturalización inicial de 5 min a 94 °C, 35 ciclos de 45 s a 94 °C, 45 s a 63 °C, 1 min a 72 °C y una extensión final de 10 min a 72 °C (O’Donnell et al., 1998).
Las pruebas de patogenicidad se llevaron a cabo sobre 80 frutos de chile manzano sanos en estado de madurez fisiológica, los cuales se inocularon con nueve aislados identificados morfológicamente como Fusarium sp. Los frutos se desinfectaron con solución de hipoclorito de sodio 0.5% durante 5 min, se sumergieron en agua destilada estéril durante 10 min y se colocaron sobre toallas de papel (Kimberly-Clark) estériles hasta secarse totalmente. Se inocularon cuatro frutos los cuales se colocaron en charolas de unicel con toallas de papel y humedad con agua destilada estéril que se colocaron dentro de bolsa de plástico para proporcionar humedad. Las charolas con los frutos se colocaron en una cámara de crecimiento bajo condiciones de incubación de 27+2 °C. La inoculación se hizo en frutos con herida (punción con aguja) y frutos sin herida sobre los cuales se colocó un disco de 5 mm de diámetro que contenían el crecimiento micelial del hongo de cada uno de aislados de siete días de edad. Como testigo se usaron chiles sin herida e inoculados con discos de PDA sin micelio del hongo. Se midió el diámetro de la lesión en los chiles inoculados y sin inocular con los aislados de Fusarium sp. cada 48 h. Las pruebas de patogenicidad se repitieron dos veces para cada tratamiento y el testigo. Para información adicional sobre el tamaño de lesión, se realizó comparación de medias (prueba de Tukey) utilizando el programa SAS V.9.1 para Windows.
De la identificación morfológica se obtuvieron nueve aislados del material procesado de los frutos de chile manzano que presentaron un crecimiento promedio de 2 cm de diámetro por día en medio de cultivo PDA. Cada aislado mostró diferente coloración en el medio de cultivo PDA por lo que se formaron tres grupos a los cuales se les asigno una clave. El primer grupo mostró crecimiento micelial de color blanco-rosa que con el paso del tiempo se tornó de color rosa en el medio de cultivo (claves FS24, FS26 y FS27). El segundo grupo mostró crecimiento blanco-crema con crecimiento afelpado, algodonoso y abundante (claves FS25, FS28 y FS32). El tercer grupo presentó un crecimiento inicialmente blanco, pero con el paso del tiempo se tornó de color rojo (claves FS30, FS31 y FS33). Estos resultados coinciden con estudios reportados por Baturo Ciesniewska et al. (2015) quienes observaron diferentes pigmentos en el medio de cultivo causados por Fusarium sambucinum que se colectó del cultivo de Solanum tuberosum. Así mismo se observó que los nueve aislados que crecieron en el medio de cultivo PDA formaron estructuras llamadas macroconidios y en el medio de cultivo SNA se tuvo formación de macroconidios en forma abundante y esporodoquios de color naranja. En el medio de cultivo de Clavel-Agar sólo se desarrolló micelio del hongo, este comportamiento fue en general para los nueve aislados. En cuanto a la forma de los macroconidios estos fueron falcados hialinos con pared delgada y por lo general bastante uniformes en tamaño presentando célula basal papilada y la célula terminal en forma de pie. El promedio del tamaño de los conidios fue 24.8-35.3 μm de largo y 3.67-4.46 μm de ancho, se observaron de tres a cinco septos por conidio (Figura 1), no hubo presencia de microconidios ni formación de clamidosporas, con estas características morfológicas los nueve aislados se identificaron como Fusarium sambucinum. Todas las características coincidieron con las descritas por Leslie y Summerell (2006) para F. sambucinum. A diferencia de F. torulosum y F. venetanum que producen clamidosporas, F. sambucinum no las produce. En cuanto F. culmorum solo se distingue por el marcador molecular con Factor de elongación (EF-1∞) que confirmó a F. sambucinum.
Las secuencias de los nueve aislados se compararon con las secuencias de las bases de datos del GenBank mostrando un 99 % de identidad con F. sambucinum en cadena de la polimerasa con Factor de elongación (EF-1∞). Las secuencias consenso se denominó CHF24 y se obtuvo mediante alineación de avance y retroceso de la secuencia, y fue depositada en Gen Bank con el número de accesión (KX632088.1). Lo anterior confirmó los resultados de la caracterización morfológica.
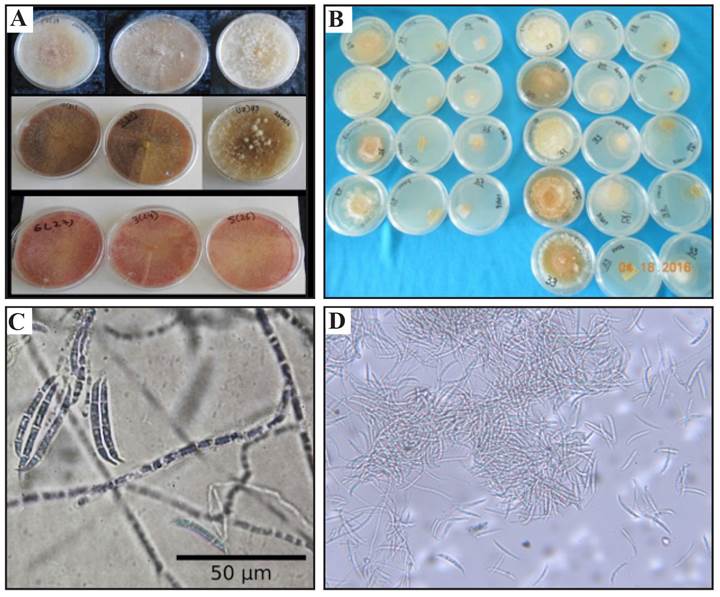
Figura 1. Caracterización cultural y morfológica de F. sambucinum (A). Colonias en medio de PDA de ocho días de edad. (B) Macroconidios de siete días de edad producidos en medio PDA 40X. (C) Crecimiento de colonias en medio de cultivo SNA y Clavel-Agar. (D) Esporodoquios producidos en medio SNA 10 X.
Los nueve aislados de F. sambucinum que se utilizaron para inocular los frutos de chile manzano con heridas fueron patogénicos en un 100%, en los tratamientos sin herida solo el 20% de los frutos inoculados presentaron síntomas, mientras que los frutos de chile testigos fueron asintomáticos. En frutos de chile con herida, se observaron lesiones circulares hundidas a las 96 h después de la inoculación (hdi), además se desarrolló micelio superficial a las 144 hdi y esporulación de conidios de a las 192 hdi (Figura 2). El hongo fitopatógeno se reaisló a partir de los frutos de chile manzano infectados con cada aislado de F. sambucinum, para confirmar los postulados de Koch; se observaron las mismas características morfológicas de crecimiento del hongo tanto en los reaislamientos como en las inoculaciones originales. En la comparación de medias de los tratamientos con herida no se observaron diferencias significativas entre el diámetro de la lesión en los nueve aislados evaluados (p >0.05) (Cuadro 1).

Figura 2 Síntomas de hundimiento observados a las 96 hdi en frutos de chile manzano inoculados con discos de F. sambu cinum (A). Presencia de micelio blanquecino a las 144 hdi (B y C). Presencia de esporulación de conidios a los 192 hdi estos fueron con herida (D). Micelio y esporulación de F. sambucinum en frutos con herida artificial (192 hdi) (E).
En este estudio se observó que los nueve aislados de F. sambucinum resultaron patogénicos. En cuanto a los dos métodos de inoculación con herida y sin herida se observó que en el primer método el 100% de los frutos inoculados presentaron síntomas como lesiones circulares, hundidas con micelio blanquecino y esporulación del hongo; mientras que, en los frutos de chile inoculados sin herida se observó que solo 20% presentaron los síntomas característicos mencionados con anterioridad en frutos de chile manzano. Estos resultados fueron similares a los reportados por otros autores (Nelson et al., 1981; Secor y Salas, 2001; Peters et al., 2008; Gachango et al., 2012; Baturo-Ciesniewska et al., 2015) quienes encontraron que el patógeno causa podredumbre seca en tubérculos de Solanum tuberosum con heridas artificiales.
Cuadro 1. Diámetro de lesión inducido por de F. sambucinum en frutos de chile manzano con heridas artificiales.
| Aislados | Diámetro de la lesión (mm) |
|---|---|
| FS24 | 18.05aZ |
| FS25 | 18.85 a |
| FS26 | 16.98 a |
| FS27 | 12.03 a |
| FS28 | 14.96 a |
| FS30 | 14.81 a |
| FS31 | 17.19 a |
| FS32 | 21.07 a |
| FS33 | 14.32 a |
ZValores con la misma letra, dentro de la columna son estadísticamente iguales (Tukey, α=0.05) / Values with the same letter within the column are statistically equal (Tukey, α=0.05).
Además, se observó que los nueve aislados cultivados en PDA formaron tres grupos de color de micelio de los cuales dos grupos coinciden con los reportados por Baturo-Ciesniewska et al. (2015) quienes agruparon los aislados en color rosa, salmón y crema brillante. Para este estudio se presentaron colonias con una coloración blanco cremoso, rosa y rojiza marrón.
En cuanto a las características morfológicas se observó que los aislados de F. sambucinum cultivados en PDA formaron macroconidios y en SNA formaron abundantes esporodoquios y macroconidios con coloración naranja, no hubo presencia de microconidios ni formación de clamidosporas dichas características coinciden con las reportadas por Leslie y Summerell (2006).
Las secuencias obtenidas a partir del DNA de los nueve aislados mostraron similitud del 99% con otras secuencias de F. sambucinum, en la base de datos del Gen Bank con número de accesión KP710620 y KP674193 las cuales se reportaron como agente causal de la podredumbre seca en tubérculos de papa (Stefancyk et al., 2016). En esta investigación todos los aislados se identificaron como F. sambucinum y una de las secuencias consenso realizadas (número de accesión KX632088.1) se depositó en Gen Bank; sin embargo, es necesario realizar un muestreo en distintas regiones del país en donde se cultiva chile manzano para conocer la distribución del patógeno en México.
En conclusión se tiene que los resultados del presente trabajo, de muestran que la identidicación morfológica, caracterización molecular y pruebas de patogenicidad del agente causal de los síntomas de lesiones hundidas con micelio blanquecino en el fruto de chile manzano es Fusarium sambucinum y las heridas favorecen la penetración del patógeno en postcosecha.
Este es el primer estudio que se realiza en México de Fusarium sambucinum en frutos de chile manzano en postcosecha.

