











 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkEl ataque de fitopatógenos es uno de los aspectos más preocupante en el cultivo de tomate (Solanum lycopersicum), tanto cuando se cultiva a cielo abierto o en sistema protegido bajo malla o invernadero. Entre las enfermedades de mayor importancia, se encuentra la pudrición de la corona (PCT) causada por el hongo saprófito Fusarium oxysporum f. sp. radicis-lycopersici (Forl), llegando a reducir los rendimientos de cosecha hasta un 50% (Apodaca et al., 2004). El patógeno puede sobrevivir en el suelo por largos períodos de tiempo, infectando las plantas, a través de heridas en la raíz o durante la formación de nuevas raíces (Szczechura et al., 2013). Una buena estrategia de control de la enfermedad, es el uso de variedades resistentes y el uso de injertos (Carrillo-Fasio et al., 2003). Sin embargo, el control de esta enfermedad depende principalmente del control químico (McGovern, 2015), existiendo reportes que después que el hongo penetra al tejido vegetal, el control químico resulta sin efecto (Apodaca et al., 2002). Un método con potencial para reducir la severidad de la enfermedad causada por Forl, es la inducción de resistencia de la planta al patógeno (Ojha y Chatterjee, 2012). Existen reportes de plantas que presentan susceptibilidad a patógenos, las cuales, después de aplicar agentes bióticos y abióticos que afectan la fisiología del hospedero, inducen a una rápida y coordinada activación del sistema de defensa (El-Kallal, 2007; Arzoo et al., 2012). Los extractos de plantas o sus componentes, han sido reportados como inductores de defensa en distintos cultivos (Baysal et al., 2002; Zaker y Mosallanejad, 2010; Arzoo et al., 2012). Particularmente interesante, son los extractos capaces de inducir al ácido salicílico (AS) y ácido jasmónico (AJ), moléculas señal que tienen una función importante en regular la red de señalización implicada en inducir las respuestas de defensa de la planta contra patógenos (Ryan y Moura 2002; Van Loon et al., 2006). La defensa de la planta es más efectiva cuando la resistencia adquirida es sistémica, ya que resulta eficiente contra un amplio espectro de patógenos (Gozzo y Faoro, 2013). La resistencia sistémica adquirida, implica el incremento de señales móviles como el AS y el AJ, compuestos que están involucrados en la expresión de genes que activan los mecanismos de defensas en los tejidos alejados al sitio donde inició el ataque, extendiendo la defensa hacia las partes no infectadas de la planta (Ojha y Chatterjee, 2012). Por lo tanto, inducir la defensa de la planta, la predispone a resistir efectivamente ataques de otros fitopatógenos (Kachroo y Robin, 2013). En este contexto, la inducción de resistencia es una técnica no convencional de control de patógenos, donde el agente inductor de la resistencia, puede no tener efecto directo sobre el patógeno (Arzoo et al., 2012). Para este estudio se aplicó un extracto de ajo a plantas de tomate para inducir la resistencia contra Forl, cuantificando la acumulación de AS y AJ como parámetros bioquímicos y la severidad de la enfermedad, altura de la planta y el contenido de clorofila como parámetro de resistencia.
MATERIALES Y MÉTODOS
Material vegetal
Se utilizaron plántulas de tomate (Solanum lycopersicum) var. Tiny tim cherry de 21 día de crecimiento, las cuales fueron propagadas a partir de semillas certificadas en un invernadero comercial. Las plántulas se trasladaron a una cámara de crecimiento a temperatura de 25 °C, fotoperiodo de 16 h luz/ 8 h oscuridad (100 µmol m-2 s-1 de intensidad lumínica) y 80% de humedad relativa. Cuando las plántulas completaron 28 días después de germinación, se aplicó un enraizador (Magic root) al 2%. Al completar 45 días después de germinación, las plántulas fueron inoculas con Forl e inmediatamente trasplantaron a bolsas de 3 kg, conteniendo sustrato preparado (Nutrigarden). Después de que las plántulas fueron trasplantadas, se procedió a designar y aplicar aleatoriamente los tratamientos a cada grupo de plantas.
Extracto de ajo (EA).
El extracto se preparó a partir de 10 g de dientes de ajo fresco (var. Río sonora), se licuó con 14 mL de agua destilada durante 1 min a temperatura ambiente. El homogenado se centrifugó por 20 minutos a 15 000 x g a 4 °C, se adicionaron 6 mL de etanol al sobrenadante y se centrifugó nuevamente a 8000 x g por 20 min a 4 °C (Jansen et al., 1987). El sobrenadante representó el extracto de ajo.
Obtención de esporas de Forl
La cepa de Fusarium oxysporum f. sp. radicis-lycopersici fue identificada molecularmente empleando la técnica de secuenciación de ITS1-5.8S ADNr-ITS2 y/o dominios D1/D2 del 28S ADNr en el Centro de Biotecnología Genómica del Instituto Politécnico Nacional (Núm. de acceso:90436915:DO452455.1 en base NCBI).La cepa de Forl fue donada para este estudio por el Laboratorio de Biotecnología Vegetal del CIBAA en ITSON centro. Para su crecimiento se utilizó como medio de cultivo agar papa dextrosa (PDA) y se mantuvo a 28 °C por 7 días en incubadora. Se resembró tomando un pequeño pedazo de micelio e inoculándolo sobre PDA fresco a 28 °C por 7 días o hasta que el hongo alcanzó una esporulación suficiente para preparar una solución de 1x107 esporas/mL. La suspensión de esporas se preparó adicionando 5 mL de agua destilada conteniendo 0.01% de Tween 80 al 0.02% sobre la superficie de la placa con el hongo, se raspó con una varilla de vidrio, el líquido conteniendo las esporas del hongo se colectó y filtró sobre gaza para eliminar micelio y el filtrado fue centrifugado a 5000 x g. Las esporas colectadas fueron re-suspendidas en agua estéril hasta lograr una concentración de 1x107 esporas/mL cuantificadas con un cámara de Neubauer.
Inoculación de plántulas de tomate con Forl
Las plántulas de tomate de 45 días después de germinadas, se extrajeron del semillero, se lavaron suavemente las raíces para eliminar el sustrato. En las raíces adventicias, se hicieron pequeñas heridas con un bisturí previamente esterilizado, inmediatamente, se sumergieron durante 30 minutos en una solución inoculante (1X107 esporas/mL) de Forl. Posteriormente se realizó el trasplante, como se describió en párrafo anterior.
Tratamientos aplicados para la inducción de resistencia en plantas de tomate
Se prepararon grupos de 10 plántulas por tratamiento con tres repeticiones. Se evaluaron seis tratamientos que se muestran en el Cuadro 1. Los primeros tres tratamientos fueron usados como controles. La inoculación se realizó con Forl (1x107 esporas mL-1) previo al trasplante. La aplicación de los tratamientos del EA, se llevó a cabo por aspersión al follaje a los 7 y 14 días después de la inoculación con Forl y trasplante.
Cuadro 1 Tratamientos utilizados para inducir resistencia en plantas de tomate infectadas con Forl.
| Tratamiento | Plantas |
| 1 | Testigo absoluto |
| 2 | Raíces heridas |
| 3 | Raíces heridas e inoculadas con Forl |
| 4 | Raíces heridas e inoculadas con Forl + EA 0.5% |
| 5 | Raíces heridas e inoculadas con Forl + EA 1% |
| 6 | Raíces heridas e inoculadas con Forl + EA 2% |
Evaluación de la severidad de la enfermedad.
Para evaluar la respuesta de las plantas de tomate al patógeno, se utilizó una escala hedónica propuesta por Apodaca et al. (2004) y Clavijo-Castro (2014). La clasificación de las plantas se realizó posterior a la aparición de la 5a hoja verdadera, se evaluó cada planta en una escala del 1 al 5 (Cuadro 2) según la intensidad de la enfermedad. Para cuantificar la severidad de la enfermedad, se evaluaron en las plantas los parámetros: índice de clorofila (UC) y altura (cm) cada 7 días después de la aplicación del tratamiento durante 49 días. El contenido de clorofila, se midió empleando el equipo SPAD 502 (Minolta). El equipo mide la absorción en longitudes de onda de 650 y 940 nm para estimar niveles de la clorofila. La medición se realizó en la tercera hoja por triplicado para obtener el índice de clorofila. La altura de la planta se midió con un flexómetro desde la base del tallo hasta el ápice de la planta.
Cuadro 2 Escala hedónica para evaluar la severidad de la enfermedad producida por Forl en plantas de tomate.
| Clase | Intensidad de la enfermedad |
| 0 | Sin síntomas visibles a la enfermedad |
| 1 | Puntos necróticos en hipocotílo |
| 2 | Hojas marchitas, oscurecimiento en la base del hipocotílo o disminución en el crecimiento de la planta |
| 3 | Marchitez, lesión necróticas 1-5 cm y disminución en el crecimiento de la planta |
| 4 | Lesión necrótica 6-10 cm, defoliación y disminución en el crecimiento. |
| 5 | Muerte de la planta |
La severidad se calculó utilizando la fórmula propuesta por Towsend y Heuberguer (1943) (1).
Severidad= (Σnv/ 5N) x 100 (1)
Donde:
S= Severidad
n= Plantas por categoría
v= Valor de la categoría
N= Plantas por unidad experimental
Cuantificación de ácido salicílico y ácido jasmónico
El contenido de AS y AJ se determinó en hojas, antes y después del trasplante, tanto en plantas tratadas y en las no tratadas, evaluando cada 7 días, durante 49 días. Las hojas se cortaron de la planta y fueron envueltas en papel aluminio, inmediatamente fueron congeladas en nitrógeno líquido y almacenadas a -80 °C hasta su análisis. Para la cuantificación de los metabolitos se siguió la metodología propuesta por Guzmán-Téllez et al. (2014) y Arbona y Gómez-Cárdenas (2008). Se partió de 1 g de hojas que se colocaron en tubos (30 mL), se les agregaron 10 mL de agua des-ionizada. Posteriormente se homogeneizó con Ultra-Turrax. Los tubos se centrifugaron (Galaxy 7D centrifuga, VWR, USA) a 4000 g durante 45 min a 4 °C. Se colectó el sobrenadante y se ajustó a un pH de 3.0, con ácido acético al 15%. Se prepararon dos muestras de 3 ml c/u del sobrenadante pH 3 y se agregaron 2 mL de éter etílico a cada tubo. Se agitó y la fase orgánica se recuperó y se llevó a sequedad bajo chorro de nitrógeno gaseoso. El residuo seco resultante se resuspendió con 500 µL de la fase móvil A/B (50/50) preparada para la determinación en HPLC. Después se filtró utilizando un filtro de PTFE de 0.45 µm (Pall Gelman Acrodisc filtros). El filtrado se utilizó para la determinación del ácido salicílico y del ácido jasmónico, utilizando un cromatógrafo de líquidos (Agilent Tecnologies modelo 1260 infinity), controlado por el programa OpenLab ChemStation (Agilent Tecnologies). El equipo consta de una bomba cuaternaria, un detector de arreglo de diodos y un inyector manual Rheodyne 7725i, la columna utilizada fue C18 (Phenomenex Luna) de 250 x 4.6 mm de 5 µm de partícula. La fase móvil consistió de una fase A compuesta de 94.9% H2O, 0.1% HCOOH (ácido fórmico) y 5% CH3CN (acetonitrilo) y una fase B de 5% H2O, 94.9% CH3CN y 0.1% HCOOH, en una proporción 60 (A):40 (B), la cual se trabajó de manera isocrática a 1 mL por minuto durante 20 min a temperatura ambiente. La longitud de onda de detección fue 303 nm para los dos compuestos. Se utilizaron compuestos puros de ácido salicílico y ácido jasmónico (Sigma Aldrich) para realizar curvas de calibración, determinar los tiempos de retención de cada compuesto y calcular la concentración. Los rangos de las curvas de calibración fueron: para AS, 0.02-0.1 µg g-1 y de 0.1-1.0 µg g-1, para AJ, 0.05-0.1 µg g-1 y 0.1-1.0 µg g-1.
Diseño experimental y análisis de datos
Se aplicó un diseño experimental completamente al azar con 6 tratamientos y tres repeticiones. El experimento se repitió dos veces para su validación. Los datos se analizaron con ANOVA (de una vía) en el paquete estadístico NCSS (Number Cruncher Statistical System, Kaysville, Utah) versión 2007. Los datos de las 7 semanas de índice de clorofila, altura, severidad y la concentración de ácido salicílico y ácido jasmónico fueron las variables respuestas. Se realizó una comparación de medias por el método de Tukey-Kramer con un nivel de probabilidad de p≤0.05. El índice de clorofila y altura, se analizaron mediante estadística descriptiva.
RESULTADOS y DISCUSIÓN
Contenido de clorofila en plantas de tomate.
El contenido de clorofila en las plantas de tomate después de una semana del trasplante fue entre 41-44 UC para todas las plantas (Figura 1). Estos valores se encuentran en el rango (41.3-56.11 UC) reportado para plantas de tomate medidos en condiciones similares a las nuestras (Mendoza et al., 1998; Peñuelas-Rubio et al., 2017). El contenido de clorofila presentó una disminución a partir de la tercera semana después del trasplante en las plantas testigo absoluto y control con raíces heridas, comportamiento asociado a la demanda de nitrógeno durante el rápido crecimiento vegetativo de las plantas (Nurzyeki, 2013). Sin embargo, las plantas control con raíces heridas e inoculas con Forl, presentaron una disminución significativa en las UC, a partir de la segunda semana después del trasplante, con valores de clorofila significativamente menores (p≤0.05) que los otros dos grupos de controles (Figura 1). Pudiendo atribuirse esta disminución en las UC, a la presencia de Forl, como lo reportaron Mendoza et al. (1998) y Hiderman et al. (1992).
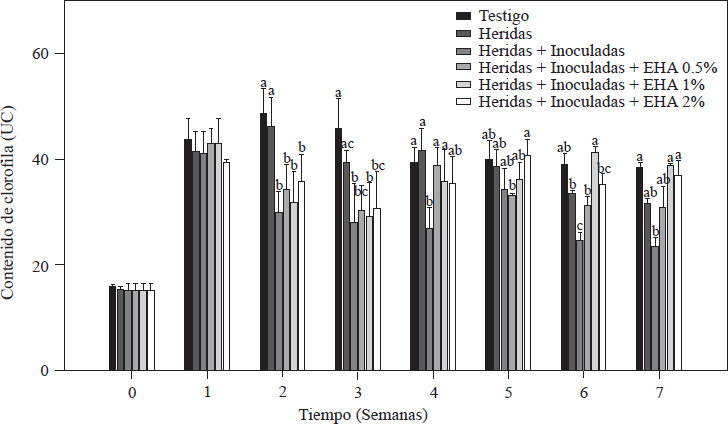
Figura 1 Contenido de clorofila en plantas de tomate inoculadas y no-inoculadas con Forl, y tratadas con diferentes concentraciones de extracto de ajo. Los valores son la media de tres repeticiones de 10 plantas cada una. Las barras verticales representan la desviación estándar (n=3). Letras diferentes significa diferencia significativa entre tratamientos de acuerdo con Tukey-Kramer para p≤0.05. El experimento se repitió dos veces.
Las plantas tratadas con EA, lograron incrementar las UC. Específicamente, las plantas que recibieron los tratamientos de 1 y 2% de EA, incrementaron significativamente (p≤0.05) los valores de UC, logrando en las dos últimas semanas de la evaluación, alcanzar los valores del testigo absoluto, que presentó un valor de clorofila de 41.3 UC, y las plantas con EA 1 y 2% con valores de 41.5 y 40.5 UC, respectivamente. Massacci et al. (2008) reportó que las plantas infectadas con F. oxysporum no logran llevar a cabo procesos fotosintéticos, reduciendo la tasa de asimilación fotosintética. Coincidiendo con los datos obtenidos en esta evaluación, donde los menores valores de clorofila (24.9 UC) se presentaron en el grupo de plantas control con raíces heridas e inoculadas que no recibieron tratamiento de EA, y no lograron recuperar los valores de clorofila, como lo hicieron las plantas que recibieron los tratamientos con EA. Las UC es una medida indirecta de la cantidad de N en la planta (Vázquez et al., 2012), por lo que se asume que, las plantas tratadas con el EA, mantuvieron un buen desarrollo, aun cuando fueron infectadas con Forl.
Altura de las plantas de tomate
Los resultados de las mediciones de la altura de las plantas de tomate se muestran en la Figura 2. Se puede observar que las tres primeras semanas después de la inoculación y trasplante, todas las plantas crecieron sin mostrar diferencias significativas entre las plantas tratadas con EA y las no tratadas (p≤0.05). Después de la cuarta semana, los grupos de plantas, marcaron las diferencias en el crecimiento, siendo las plantas que recibieron el tratamiento con el EA en concentraciones de 1 y 2%, las que presentaron el mayor crecimiento, alcanzando los valores de 67.8 cm para las plantas tratadas con EA al 2%, 62.2 cm las plantas con EA al 1%, valores similares (p≤0.05) al 61.9 cm al que presentó el grupo de testigo absoluto. Las plantas tratadas con EA al 0.5%, alcanzaron una altura promedio de 53.5 cm, valor superior al 34.9 cm, que presentó el control de raíces heridas + inoculadas, ambos grupos presentan valores estadísticamente menores (p≤0.05) que los de los tratamientos con 1 y 2% de EA (Figura 2). La disminución en el crecimiento, es un síntoma característico de plantas infectadas por Forl (Apodaca et al., 2004; Peñuelas-Rubio et al., 2017), debido a la proliferación del patógeno en los haces vasculares de la planta, diseminando las esporas hacia el ápice mediante el flujo del xilema (Hadian et al., 2011). Concordando con los resultados observados en las plantas control con heridas en la raíz + inocularon con Forl, sin recibir tratamiento con EA, estableciéndose la enfermedad y propagándose intercelularmente. El tratamiento de EA al 0.5%, indujo la menor resistencia a la planta de tomate ya que el grupo de plantas que recibió este tratamiento, presentó la menor altura de los tres tratamientos de EA. El efecto del ajo como promotor de crecimiento vegetativo y reproductivo en plantas infectadas con patógenos, esta reportado por Fatema y Ahmad (2005), en plantas de cacahuate infectadas con nematodos, y en plantas de tomate infectada con Fol por Chohan y Perveen (2015).En ambos casos las soluciones acuosas de ajo mejoraron la respuesta en el crecimiento de la planta y controlando al patógeno. En este estudio las concentraciones del EA de 1 y 2% mejoraron el desarrollo de las plantas induciendo la resistencia a Forl.
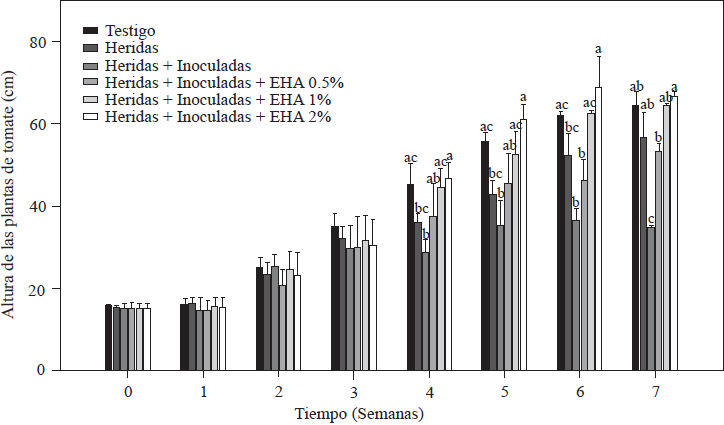
Figura 2 Evaluación de la altura de plantas de tomate inoculadas y no-inoculadas con Forl y tratadas con extracto de ajo con diferentes concentraciones. Los valores son la media de tres repeticiones de 10 plantas cada una. Las barras verticales representan la desviación estándar (n=3). Letras diferentes significa diferencia significativa entre tratamientos de acuerdo con Tukey-Kramer para P≤0.05. El experimento se repitió dos veces.
Severidad de la enfermedad de plantas de tomate infectadas con Forl
Los resultados de la evaluación de la severidad de la enfermedad provocada por Forl en las plantas de tomate, se muestra en la Figura 3. Todas las plantas inoculadas con Forl, presentaron en diferentes grados, síntomas característicos de la enfermedad, incluyendo, marchitez en las hojas y defoliación, disminución en el crecimiento y afectaciones en el tallo como puntos y lesiones necróticas en los haces vasculares. Como se esperaba, las plantas testigo absoluto y las de raíces heridas, no presentaron síntomas a la enfermedad ya que no fueron inoculadas con el hongo (Figura 3). El valor mínimo de severidad de la enfermedad fue del 26.4%, que se presentó en el grupo de plantas que recibieron el tratamiento con el EA al 1 y 2%, seguido del tratamiento con EA 0.5% con 33.4%. Las plantas más afectadas fueron el grupo control de raíces heridas + inoculadas, que mostraron síntomas visibles desde la segunda semana después de la inoculación, presentando una severidad del 100% en la séptima semana. Estos resultados muestran el efecto positivo de la aplicación del EA, donde las plantas tratadas presentaron una reducción de daños entre el 66 y 73 %, comparado con la severidad presentada por las plantas control (heridas + inoculadas), lo que significa que los síntomas ocasionados por el hongo disminuyeron en las plantas tratadas. Este efecto en la reducción de la severidad de la enfermedad provocada por Forl en plantas de tomate tratadas con EA, podría atribuirse a la acumulación de metabolitos como el AS y AJ que inducen los mecanismos de defensa, como lo demostraron Shah y Zeier (2013) y El Oirdi et al. (2011) donde, estos metabolitos, indujeron la expresión de genes involucrados en la defensa en condiciones de estrés. Basado en este resultado se puede inferir que la aplicación exógena del EA funcionó como activador de la resistencia sistémica adquirida en las plantas de tomate.
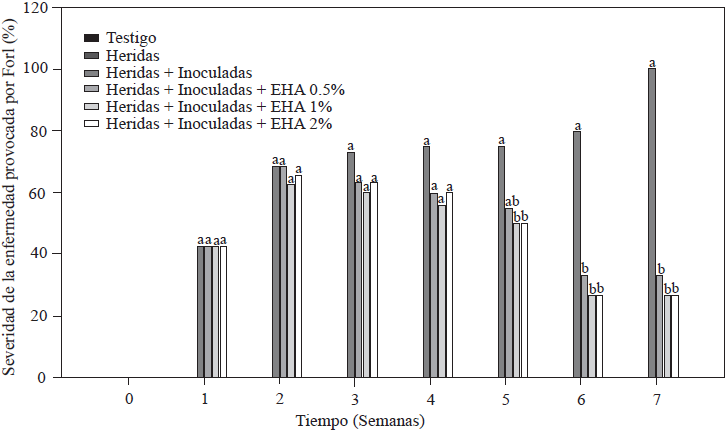
Figura 3 Porcentaje de severidad de la enfermedad provocada por Forl en plantas de tomate inoculadas y no-inoculadas con Forl y tratadas con diferentes concentraciones de extracto de ajo. Los valores son la media de tres repeticiones de 10 plantas cada una. Letras diferentes significa diferencia significativa entre tratamientos de acuerdo con Tukey-Kramer para P≤0.05. El experimento se repitió dos veces.
Cuantificación de ácido salicílico (AS)
Los resultados obtenidos en la cuantificación de AS en las plantas de tomate se muestran en la Figura 4. Puede observarse que las variaciones en el contenido de AS presentaron cambios desde la semana de la inoculación de las plantas, cuantificando niveles basales de 0.10 µg g-1 en las plantas testigo absoluto, valor dentro del rango (0.10-10 µg g-1) reportado para plantas Solanáceas (Rivas-San Vicente y Plasencia, 2011). En las plantas heridas e inoculadas se cuantificó una concentración inicial de 0.24 µg g-1, dos veces la concentración del control absoluto. En las plantas inoculadas con Forl y tratados con EA 1 y 2%, mostraron un incremento significativo (p≤ 0.05) del contenido de AS en la tercera semana después de la inoculación. En las plantas con EA al 2%, se cuantificaron valores 11 y 6 veces más altos que los que se presentaron en los controles para esa misma semana y en el tratamiento con EA al 1%, se cuantificaron 8 y 4.5 veces más los valores que sus respectivos controles. Después de este incremento, los niveles de AS decrecieron las semanas posteriores, volviendo a incrementarse en la sexta semana (Figura 4). Las plantas control (testigo absoluto y heridas + inoculadas), a partir de la quinta semana mostraron un incremento en la acumulación de AS, alcanzando la máxima concentración de 0.78 y 0.82 µg g-1, respectivamente. Estos incrementos en los controles, se presentaron dos semanas posterior al incremento que se presentó en el grupo de plantas tratadas con el EA al 1 y 2%. Los incrementos en el contenido de AS entre 2 a 5 veces los valores basales, son reportados en plantas durante el inicio de la floración, así como en plantas infectadas con patógenos necróticos como Fusarium oxysporum (Swarupa et al., 2014). Pudiendo ser esta la razón de los incrementos del AS en las plantas control en las semanas de la quinta a la séptima. Sin embargo, los incrementos de más de 5 veces la concentración de AS que se presentaron en la tercera semana en las plantas tratadas con EA al 1 y 2%, pudiera estar asociada al tratamiento recibido más que a la presencia del patógeno, debido a que los valores máximos de AS en los tratamientos con el EA de 1 y 2%, concuerda con la disminución de los síntomas observados durante la evaluación de la severidad de la enfermedad (Figura 3). Apoyando está deducción, Ojha y Chatterjee (2012), aplicando AS a plantas de tomate infectadas con F. oxysporum, significativamente incremento la resistencia de la planta al ataque del patógeno al aumentar la actividad de enzimas relacionadas con el sistema de defensa, presentándose los mayores incrementos en la actividad de las enzimas, a los 21 días después de la aplicación del AS. Coincidiendo con el resultado obtenido en este estudio, los valores más altos de AS se cuantificaron a la tercer semana después de la aplicación del tratamiento. El efecto protector contra Forl, que mostraron las plantas tratadas con el EA, manifestado en mejora en el crecimiento y desarrollo de la planta, también fue reportado por Hayat et al., (2012), al aumentar concentraciones endógeno de AS, mejoró significativamente las características de crecimiento de las plantas incluyendo la altura, el número y color de hojas, el diámetro del vástago, el peso seco y fresco de la planta, así como también la tasa fotosintética.
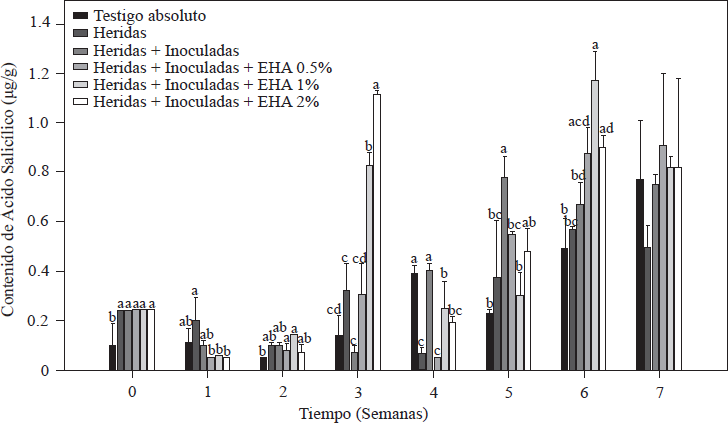
Figura 4 Concentración de ácido salicílico en plantas de tomate inoculadas y no-inoculadas con Forl y tratadas con diferentes concentraciones de extracto de ajo. Los valores son la media de tres repeticiones de 10 plantas cada una. Las barras verticales representan la desviación estándar (n=3). Letras diferentes significa diferencia significativa entre tratamientos de acuerdo con Tukey-Kramer para p≤0.05. El experimento se repitió dos veces.
Para el caso de los resultados de este estudio, el incremento en la concentración de AS coincide con la mejora del desarrollo de la planta, así como la resistencia al patógeno, lo que concuerda con el papel importante reportado del AS en la acción protectora contra patógenos. Probablemente por la capacidad que tiene de inducir la transcripción de proteínas de defensa (PR) en las que codifican enzimas quitinasas, glucansas, endohidrolasas, y β-1, 3-glucanasa para contribuir con la resistencia sistémica adquirida (RSA) contra diversos patógenos por su actividad antifúngica (Heil y Bostock, 2002; Hayat et al., 2010).
Cuantificación de ácido jasmónico (AJ)
Los resultados obtenidos en la cuantificación de AJ en las plantas de tomate se muestran en la Figura 5. Se puede observar que tanto las plantas con raíz herida, como las de raíz herida + inoculadas con Forl y a las que se les aplicaron los EA, provocaron un incremento en el contenido de AJ en las plantas de tomate, cuantificando valores iniciales de 0.89 µg g-1 en plantas con herida y/o inoculadas con Forl, valor significativamente mayor (p≤0.05) que el de las plantas testigo absoluto con 0.74 µg g-1. A partir de la segunda semana, la aplicación de EA al 1 y 2 % indujeron un incremento de 2.5 veces el valor del testigo absoluto y de 1.8 veces el del control con herida + Forl. Estos valores de AJ se mantuvieron significativamente (p≤0.05) altos en la tercera semana después de la inoculación y trasplante en las plantas tratadas con EA al 1 y 2%. En todos los tratamientos, con excepción del testigo absoluto, el contenido de AJ disminuyó gradualmente a partir de la cuarta semana hasta el final de la evaluación, conservando niveles altos, lo que nos indica que la planta está respondiendo a alguna situación de estrés (Ryan y Moura, 2002). De este resultado podemos inferir que las concentraciones del EA del 1 y 2% es suficiente para incrementar concentraciones endógenas de AJ en las plantas de tomate, el cual a su vez puede inducir respuestas de defensa en la planta (Kravchuk et al., 2011). El AJ es reconocido como un regulador clave para estimular en las plantas las respuestas de defensa en sinergia con el AS, reduciendo los síntomas de las enfermedades ocasionados por patógenos (Pieterse et al., 2012), mediante la activación en la expresión de genes involucrados en la defensa (Ryan, 2000). Específicamente, en plantas de tomate se ha reportado que el AJ induce inhibidores de proteasas y polifenoloxidasa y compuestos de la ruta de los fenilpropanoides, mostrando un efecto en la disminución de patógenos (El Oirdi et al., 2011). Es bien aceptado que las plantas requieren activar la resistencia sistémica inducida dependiente de AJ, para combatir patógenos necrotróficos (Glazebrook, 2005). Por lo que la protección de las plantas de tomate contra Forl, observada en este estudio, puede estar relacionada con la rápida inducción de AJ y las concentraciones endógenas cuantificadas (Rahman et al., 2012). Sin embargo, la activación simultanea de los sistemas de defensa dependiente de AS y AJ, resultan en un incremento de la resistencia a patógenos (Mur et al., 2006). Lo anterior concuerda con los resultados observados en los incrementos endógenos de AS y AJ y la disminución de los síntomas de la enfermedad a producida por Forl en plantas de tomate.
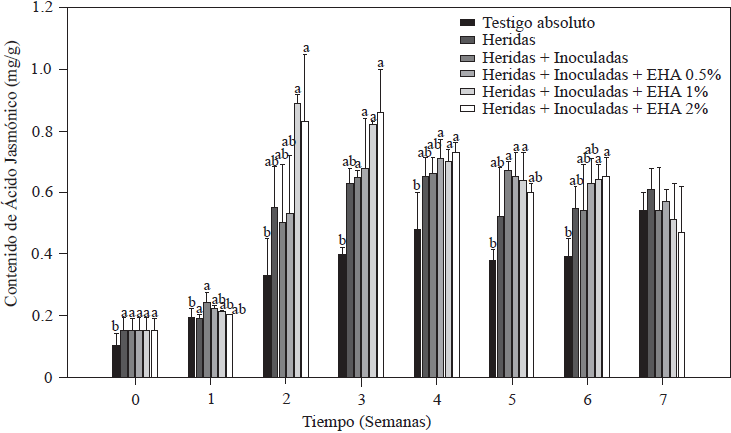
Figura 5 Concentración de ácido jasmónico en plantas de tomate inoculadas y no-inoculadas con Forl y tratadas con diferentes concentraciones de extracto de ajo. Los valores son la media de tres repeticiones de 10 plantas cada una. Las barras verticales representan la desviación estándar (n=3). Letras diferentes significa diferencia significativa entre tratamientos de acuerdo con Tukey-Kramer para p≤0.05. El experimento se repitió dos veces.
CONCLUSIONES
Con los resultados obtenidos en este estudio, se puede concluir que la aspersión de un EA al 1 y 2% a plantas de tomate le proporcionó la resistencia contra Fusarium oxysporum radicis-lycopersici (Forl), mediado por el incremento del contenido endógeno de ácido salicílico y ácido jasmónico.

