











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkMéxico es el principal exportador de sandía en el mundo, aportando 23% de este fruto al comercio mundial (FAO, 2014), principalmente a Estados Unidos, Canadá y los Países Bajos. En los últimos diez años las exportaciones han crecido a una tasa promedio anual de 8% (SAGARPA, 2014). Sonora es el principal productor de sandía (Citrullus lanatus) en México. En 2016 se plantaron más de 9 000 ha en este estado (SIAP, 2017). No obstante, uno de los factores limitantes en la producción de sandía en este estado, son las enfermedades radiculares. En 2013, en ciertas plantaciones de sandía establecidas en los valles de Guaymas y la Costa de Hermosillo en Sonora, regiones que concentran alrededor del 96 % de la superficie estatal establecida con este cultivo, se observó que, previamente a su madurez fisiológica, ciertas plantas presentaron un amarillamiento y marchitamiento de hojas; días después las plantas murieron. El análisis visual demostró la presencia de lesiones y pudrición en la corteza de la base del tallo y la parte superior de la raíz principal, además de pudriciones en las raíces principales y secundarias, típicas de las enfermedades ocasionadas por hongos (Meza-Möller et al., 2014).
A la fecha no se tiene conocimiento de trabajos publicados acerca del complejo de hongos asociados a la pudrición de raíces en las plantas de sandía cultivadas en Sonora; generalmente los diagnósticos se basan solo en la sintomatología del cultivo y en el mejor de los casos en la morfología de las colonias y observación de estructuras reproductivas. Recurrentemente se menciona que las enfermedades radiculares de sandía cultivada en la región son debidas al ataque de: Fusarium oxysporum, Fusarium solani o Rhizoctonia solani.
Las especies pertenecientes al género Fusarium (Nectriaceae, Hypocreales, Sordariomycetes, Ascomycota), son ubicuas y de gran importancia económica en la agricultura, ya que muchas de ellas son patógenas para las plantas. Algunas de sus especies también producen toxinas nocivas para humanos y animales. Este grupo monofilético está conformado por 20 clados que incluyen más de 300 especies. Con algunas excepciones, las especies de Fusarium producen las características macroconidias multiseptadas y con forma de huso; pero además existen otras características morfológicas que permiten diferenciar entre especies (O’Donnell et al., 2013; Geiser et al., 2013; O’Donnell et al., 2015).
En el complejo Rhizoctonia, la morfología hifal y configuración del septo permiten diferenciar los géneros; mientras que las especies pueden ser distinguidas por el número de núcleos presentes en las células somáticas de hifas jóvenes y el grosor de las hifas guías, o por las características morfométricas de las estructuras reproductivas sexuales (Cedeño, 2008). El grupo de Rhizoctonia multinucleadas incluye a R. solani, R. zeae yR. oryzae.
R. solani [teleomorfo: Thanatephorus cucumeris] (Ceratobasidiaceae: Cantharellales: Agaricomycetes: Basidiomycota), agrupa una mezcla heterogénea de cepas causantes de pudrición radicular en muchos cultivos alrededor del mundo (González, 2013). De acuerdo al análisis de fusión hifal (anastomosis), estas cepas se separan en 14 grupos anastomósicos (AG), designados desde AG-1 hasta AG-13 más AG-BI (Carling et al., 2002). El grupo de Rhizoctonia binucleadas corresponde a los teleomorfos Ceratobasidium spp. y Tulasnella spp. De acuerdo a Sharon et al. (2008), Ceratobasidium consta de 21 grupos anastomósicos identificados como AG-A hasta AG-U, algunos de los cuales son altamente patogénicos en diferentes especies vegetales.
En el pasado la taxonomía de hongos se basaba en la morfología de sus estructuras reproductivas en el estado anamórfico. El concepto de especie morfológica aún prevalece como el método de diagnóstico más usual para diferenciar entre especies de hongos, debido a que los caracteres morfológicos de los individuos son fácilmente detectables y comparables. Sin embargo, no es un método capaz de detectar diferencias entre especies cercanas, subestimando la verdadera diversidad fúngica (Taylor et al., 2000).
Las técnicas moleculares basadas en análisis de ADN superan las desventajas de la identificación morfológica ya que son rápidas, precisas, objetivas y aplicables a un gran número de muestras. Permiten diferenciar entre genotipos y establecer índices de variabilidad genética existente dentro de una población (Narayanasamy, 2011). En años recientes se ha popularizado el concepto de especie filogenética entre los hongos filamentosos, basado en la concordancia de secuencias de múltiples genes de ADN; este enfoque filogenético permite definir mejor las especies (Taylor et al., 2000; Choi et al., 2013).
Desde que White et al. (1990) publicaron las secuencias de cebadores que permitieron la amplificación y secuenciación de secciones del operón de rDNA, surgió un marcado interés en la investigación filogenética, que ahora domina la taxonomía fúngica. La secuenciación de los fragmentos ITS de rDNA, sigue siendo el enfoque más ampliamente aceptado en la micología molecular para clasificar e identificar especímenes o cultivos de hongos desconocidos. Sin embargo, su resolución en relaciones taxonómicas de nivel superior es inferior a muchos otros genes. Numerosos estudios se han realizado para identificar loci con características de código de barras primarias adecuadas. El proyecto AFTOL (Assembling the fungal tree of life), completado en 2008, ha establecido una filogenia basada en la amplificación de los genes: RPB1, RPB2, nucLSU, nucSSU, mtSSU, TEF1α y mtATP6 (Stielow et al., 2015). El conocimiento de las especies causantes de una enfermedad es indispensable para su adecuado manejo y control.
En base a lo anterior, el objetivo del presente trabajo fue identificar las especies de hongos causantes de pudrición radicular en plantas de sandía en la Costa de Hermosillo y Valle de Guaymas en Sonora, en base a análisis morfológicos y de filogenia multilocus.
MATERIALES Y MÉTODOS
Muestreo: El estudio se realizó durante los ciclos primavera-verano 2013 y 2014, en cuatro campos comerciales, dos localizados en la Costa de Hermosillo (CH1, CH2) y dos en el Valle de Guaymas (VG1, VG2), en Sonora, México que representan el 10% de la superficie establecida con sandía en el estado. Las variedades cultivadas en estos campos fueron Sugar Red, SuperSeedless 7187HQ F1 y Precious Petit. En cada ciclo se colectaron aleatoriamente 40 plantas (10 plantas por campo) en las zonas donde se observaba marchitez y secazón de guías. Las muestras fueron colocadas en bolsas de polietileno etiquetadas y transportadas en contenedores con hielo al laboratorio para su procesamiento.
Aislamiento fúngico. Las raíces de plantas enfermas se lavaron con agua, se secaron con papel secante y se cortaron en pedazos de 1 cm. Se tomaron segmentos de la corona, raíz principal y raíces secundarias. Se desinfestaron sumergiéndolos por 2 min en una solución preparada con hipoclorito de sodio al 6%, alcohol etílico al 96% y agua destilada esterilizada en proporción 1:1:8, respectivamente. Posteriormente se enjuagaron dos veces con agua destilada estéril, se secaron en papel secante estéril y se colocaron en cajas con agar-agua al 2%. Las cajas se incubaron a 25 ± 0.1 °C hasta que el micelio emergido de los trozos vegetales permitió tomar una punta de hifa. Las puntas de hifas fueron cultivadas sucesivamente en Agar Dextrosa y Papa (PDA) suplementado con una solución de estreptomicina/neomicina e incubados a 25 ± 0.1 °C, hasta obtener un cultivo puro.
Caracterización morfológica y cultural. Todos los aislados fueron agrupados en base a las características de las colonias y al color desarrollado en el anverso y reverso de la caja de PDA. Un aislado representativo de cada grupo fue utilizado para la caracterización morfológica. En los aislados con características de Fusarium se determinó la presencia y morfología de microconidias, macroconidias y clamidosporas a partir de su crecimiento en agar hojas de clavel (CLA), después de siete días en oscuridad a 25 ± 0.1 °C (Leslie and Summerell, 2006). Los aislados con características del género Rhizoctonia se identificaron mediante la observación de características vegetativas como la coloración del micelio, septos, constricciones cerca de la ramificación, durante el crecimiento en PDA o agar extracto de malta (MEA). Para la determinación del número de núcleos, las hifas se tiñeron con azul de tripano en lactofenol. La velocidad de crecimiento se determinó en PDA, manteniendo los cultivos a 25 ± 0.1 °C y fotoperiodo de 14h/10h, de luz/oscuridad. Se midió el diámetro de las colonias cada 24 h, hasta que el micelio cubrió completamente la caja (Sneh et al., 1996). Todos los aislados fueron identificados inicialmente hasta nivel de género.
Extracción de ADN. El micelio proveniente de cultivos puros en PDA, se recogió con espátula microbiológica estéril y se colocó en un tubo del Kit Power Soil DNA Isolation (MoBIO Laboratories, California, EUA). La lisis celular se llevó a cabo en un homogeneizador Precellys Evolution (Bertin Technologies, Francia), agitando los tubos a 6500 rpm durante tres ciclos de 20 s con pausas de 20 s. La integridad del ADN se verificó en un gel de agarosa al 2%. El ADN extraído, se cuantificó en el NanoDrop 1000 (ThermoScientific). Sólo se amplificaron muestras con una relación de absorbancia 260/280 entre 1.8 y 2. El ADN se almacenó a -20 °C hasta su uso.
Amplificación y secuenciación de ADN. Se amplificó la región no codificante del espaciador transcrito interno (ITS) y una parte de la región que codifica para la subunidad mayor de la ARN polimerasa II (RPB2) de todos los aislados obtenidos. Adicionalmente, se amplificó la región que codifica para el factor de elongación de la transcripción 1a (TEF-1 α) para los aislados con características de Fusarium. La información sobre los primers empleados se muestra en la Cuadro 1.
Cuadro 1 Oligonucleótidos empleados en este estudio.
| Locus | Primer | Oligonucleótidos (5'-3') | (pb) | Referencia |
| ITS | ITS1 ITS4 | TCCGTAGGTGAACCTGCGG TCCTCCGCTTATTGATATGC | = 550 | Whiteet al.(1990) |
| TEF-1a | EF1 EF2 | ATGGGTAAGGARGACAACAC GGARGTACCAGTSATCAT | = 700 | O'Donnell et al. (1998) |
| RPB2x | RPB2-5F2 fRPB2-7cR | GGGGWGAYCAGAAGAAGGC CCCATRGCTTGYTTRCCCAT | = 900 | O'Donnell et al. (2013) |
| RPB2z | RPB2-980F fRPB2-7cR | TGYCCIGCIGARACICCHGARGG CCCATRGCTTGYTTRCCCAT | = 674 | Gonzálezet al.(2016) |
xEspecíficos para el género Fusarium
zEspecíficos para el género Rhizoctonia
La reacción en cadena de la polimerasa se realizó mezclando 12.5 ml de GoTaq® Green Master Mix (Promega), 1 ml de cada primer forward y reverse (IDT Technologies) a una concentración 10 mM, 1 μl de ADN a una concentración de 1 ng/ml y agua grado biología molecular hasta obtener un volumen final de 25 μl. Los productos de PCR se separaron mediante electroforesis en gel de agarosa al 2%, mezclados previamente con GelRed (Biotium Inc) en 5X Green GoTaq reaction buffer (Promega) 15 μl:1ml. Se visualizaron en luz UV (DigiDoc-It™, UVP) observando el tamaño del amplicón y su pureza. La purificación se realizó con ExoSAP-IT PCR Product Cleanup (Affymetrix) o mediante el corte de bandas del tamaño esperado con el kit Wizard® SV Gel and PCR Clean-Up System (Promega). Los amplicones purificados se secuenciaron en ambas direcciones con el equipo ABI 3730xl DNA Analyzer (Applied Biosystems) en GENEWIZ. Cada secuencia se revisó manualmente y los nucleótidos en posiciones ambiguas se corrigieron con las secuencias complementarias obtenidas con ambos primers, usando el software ChromasPro v2.1.6. Las secuencias de las regiones ITS, RPB2 y TEF de Fusarium se compararon mediante alineamiento, con las contenidas en las bases de datos: Fusarium MLST (http://www.cbs.knaw.nl/Fusarium), Fusarium ID (http://isolate.fusariumdb.org) y National Center for Biotechnology Information (NCBI), donde también se compararon las secuencias de las regiones ITS y RPB2 de Rhizoctonia spp., mediante la herramienta “Basic local alignment search tool” (BLAST) (Altschul et al., 1990).
Análisis filogenético. La determinación de las especies de Fusarium y de los grupos anastomósicos de Rhizoctonia y Ceratobasidium, se realizó por separado en dos matrices de datos. En cada caso, se realizaron múltiples alineamientos usando el software Clustal Omega (Sievers et al., 2011). La concatenación y edición de las secuencias se realizó con el software UltraEdit32. El análisis filogenético de cada matriz de datos se realizó por separado bajo el criterio de Máxima Verosimilitud (MV) con el software PAUP 4.0a152 (Swofford, 2002). Se estableció el mejor modelo de sustitución de nucleótidos con ModelTest (Posada y Crandall, 1998).
Los árboles se visualizaron y modificaron en FigTree y se exportaron a editores gráficos. Los árboles consenso para Rhizoctonia spp. y Fusarium spp., se enraizaron con Botryobasidium simile (aislado GEL2348) y Neofusicoccum parvum (cepa CCF216), respectivamente.
Pruebas de patogenicidad. En cumplimiento con los postulados de Koch, para comprobar que los aislados obtenidos son los agentes causales de la pudrición de raíz en plantas de sandía, se seleccionó aleatoriamente un cultivo monospórico de cada una de las especies identificadas en el análisis molecular. Los tratamientos fueron: 1) Testigo sin inocular, 2) F. falciforme, 3) F. oxysporum, 4) F. brachygibbosum, 5) R. solani, 6) Ceratobasidium sp, 7) F. brachygibbosum + F. solani, 8) F. brachygibbosum + F. oxysporum, 9) R. solani + Ceratobasidium sp. 10) Ceratobasidium sp. + F. solani y 11) Ceratobasidium sp. + F. oxysporum. Se utilizaron plantas sanas de las variedades SuperSeedless 7187HQ F1 y Precious Petit, de 21 días de edad, establecidas en macetas con una mezcla de suelo:perlita (1:3). Se inocularon seis plantas por cada tratamiento. Cada planta se trató con micelio de cinco días de crecimiento en PDA. Se colocaron cuatro discos de 8 mm de diámetro alrededor de la raíz. Plantas sanas tratadas con discos de PDA estéril se utilizaron como testigo. Las macetas se colocaron en una cámara de ambiente controlado a 25 ± 0.1 °C, con un fotoperiodo de 14h/10h día/noche, hasta la aparición de los síntomas. El riego se realizó en base a los requerimientos hídricos y cada semana se aplicó solución nutritiva de Hoagland. Se observó el número de plantas enfermas. El porcentaje de raíces infectadas se determinó utilizando diez segmentos de raíz de cada planta. Estos fragmentos de tejido se desinfectaron por separado y se colocaron en PDA. La evaluación se realizó observando el desarrollo de micelio después de 7 días de incubación.
RESULTADOS
Daños observados durante el muestreo. La Figura 1A muestra los daños observados en el cultivo. Las plantas sintomáticas presentaron diferentes tipos de pudrición en la corona, tallo, raíces y raicillas. Un tipo de lesiones fueron pequeñas, hundidas y no hundidas, de color marrón (Figura 1B) y de aspecto húmedo, típicas de Rhizoctonia sp. En ciertos casos este tipo de lesiones se observaron con pequeñas pústulas. En otras plantas las lesiones fueron de color café claro, con decoloración y pudrición de haces vasculares, típicas de síntomas de daños por Fusarium spp. (Figura 1C).

Figura 1 Daños en plantas cultivadas de sandía. A) Daño en campo, B) Raíces de plantas enfermas, C) Daño vascular.
Aislamiento y morfología de las colonias y estructuras reproductivas de los hongos. En total, se obtuvieron 45 aislados fúngicos provenientes de los cuatro sitios de muestreo y en cada sitio se aisló al menos una especie de Rhizoctonia y Fusarium. La distribución de especies por sitio se muestra en la Figura 2.
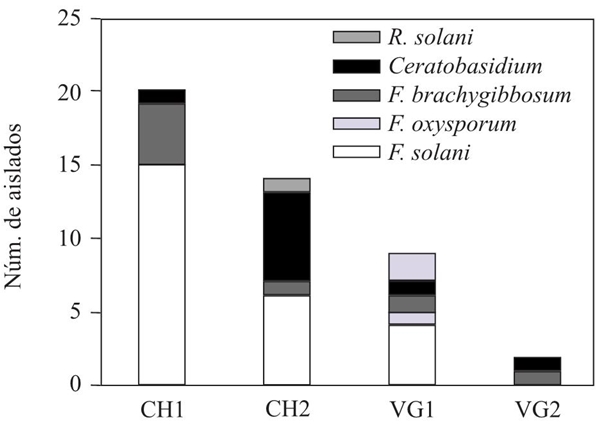
Figura 2 Distribución de especies de hongos patogénicos, por sitios de muestreo. CH1= Costa de Hermosillo 1. CH2=Costa de Hermosillo 2. VG1=Valle de Guaymas 1. VG2=Valle de Guaymas 2.
El análisis morfológico permitió formar dos grupos de aislados. Un grupo formado de 13 aislados que presentaron colonias con abundantes hifas aéreas, de color marfil al inicio, tornándose café claro o marrón después de siete días. En estos aislados las hifas fueron robustas con ramificaciones en ángulo recto, constricción de la ramificación y formación de un septo cercano al punto de origen, sin presencia de esporas, características típicas de Rhizoctonia sp. (Sneh et al., 1996). La velocidad de crecimiento promedio fue de 1.25 mm h-1. La tinción de núcleos reveló la presencia tanto de aislados con células binucleadas, como multinucleadas (Figura 3).
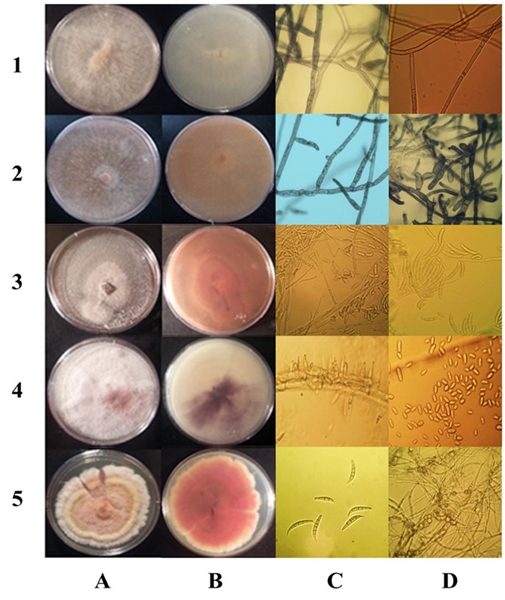
Figura 3 Morfología de las colonias y coloración del medio en PDA de Rhizoctonia spp., Ceratobasidium sp., F. falciforme, F. oxysporum, F. brachygibbosum, (1A-5A y 1B-5B) respectivamente. (1C) Hifas polinucleadas. (1D) Septos. (2C) Hifas binucleadas. (2D) Células monilioides. (3C) Monofialides y microconidias. (3D) Macroconidias. (4C) Microconidia in situ en CLA. (4D) Microconidia. (5C) Macroconidia. (5D) Clamidosporas.
El segundo grupo de 32 aislados mostró la formación de macroconidias, microconidias, clamidosporas y monofiálides en CLA, típicas de Fusarium sp., según las descripciones de Leslie y Summerell (2006). 25 aislados se ajustaron a la descripción de F. solani: colonias color crema con pigmentos rojo a gris oscuro en el anverso, microconidias ovales sin septos, monofiálides largas, clamidosporas solas o en pares, abundantes macroconidias rectas con 3 a 5 septos. Otros 2 presentaron características propias de F. oxysporum: micelio algodonoso, escaso blanco a violeta pálido y morado en el agar; monofiálides cortas. El resto de los aislados produjo micelio blanco, el cual se tornaba rosado, con esporodoquios de color amarillo; microconidias ovales unicelulares, producidas en monofiálides, macroconidias curveadas de 3 a 5 septos, con células centrales anchas, ápice ligeramente agudo, clamidosporas solas o en cadena (Figura 3). No se detectaron aislados con morfología diferente a Rhizoctonia o Fusarium.
Identificación molecular y análisis filogenético de los hongos. Un primer análisis BLAST a partir de la región comprendida entre el espaciador transcrito interno I y II (ITS1-ITS2) de todos los aislados, permitió determinar que 25 secuencias tenían un 99-100% de similitud a F. solani; 5 a F. brachygibbosum, 2 a F. oxysporum, 10 a Ceratobasidium sp. y 3 secuencias a Rhizoctonia solani también con 99-100% de similitud con secuencias homólogas de NCBI.
El análisis filogenético bajo el criterio de MV para el género Fusarium se realizó con la matriz concatenada de las secuencias de los genes ITS, RPB2 y TEF1 de los 32 aislados de este estudio y de 25 cepas de referencia depositadas en colecciones de cultivos (Al-Hatmi et al., 2016). El mejor modelo de sustitución de nucleótidos fue TIM2+I+G. Se consideraron 1816 nucleótidos en el conjunto de datos. El análisis multilocus permitió definir la identidad correcta de los aislados inicialmente propuestos como F. solani, ya que éstos forman un clado separado con 100 % de similitud con los aislados tipo de F. falciforme. Un integrante del complejo de especies de Fusarium solani (FSSC). La identidad de F. brachygibbosum y F. oxysporum fue corroborada (Figura 4).
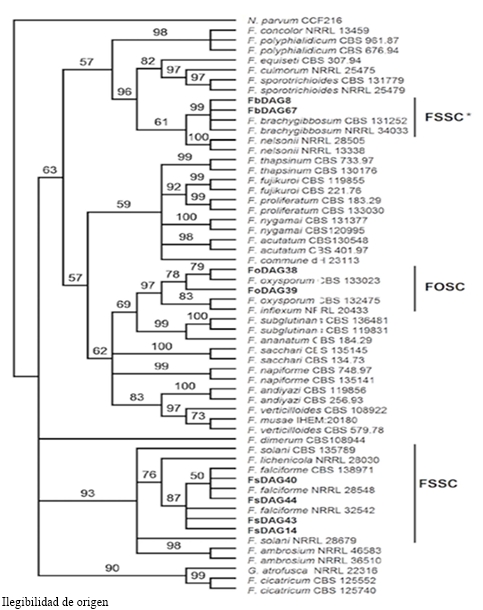
Figura 4 Filogenia por MV de las secuencias concatenadas de los genes ITS, EF y RPB2 de 8 aislados de Fusarium spp. (en negritas) representativos de 29 obtenidos de plantas de sandía con marchitez y pudrición de raíz. Bootstrap de 1000 réplicas; grupo externo: Neofusicoccum parvum .
Un análisis similar, se realizó con las secuencias de los aislados de Rhizoctonia y Ceratobasidium sp. Se procesó una matriz integrada por las regiones ITS y RPB2 de 49 aislados, incluyendo los del presente estudio y 36 cepas de referencia (Cantharellales), obtenidas de colecciones de cultivos (González et al., 2016). El mejor modelo de sustitución de nucleótidos fue GTR+I+G. Se consideraron 1355 nucleótidos de cada aislado en el conjunto de datos. Este análisis estableció la identidad de dos grupos anastomósicos para el género, AG-4 y AG-6 y dos para el género Ceratobasidium, AG-A y AG-F. (Figura 5). El Cuadro 2 muestra la identidad y los números de accesión de las secuencias obtenidas.
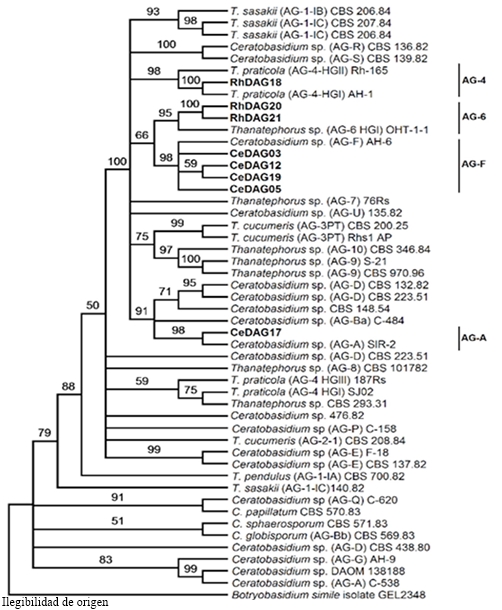
Figura 5 Filogenia por MV de las secuencias concatenadas de los genes ITS y RPB2 de 8 aislados de Rhizoctonia spp. (en negritas) representativos de 16 obtenidos de plantas de sandía con marchitez y pudrición de raíz. Bootstrap de 1000 réplicas; grupo externo: Botryobasidium simile . AG=Grupo anastomósico.
Cuadro 2 Identidad, localización y números de accesión de secuencias obtenidas en el presente estudio.
| Identidad | Nombre del aislado | Sitio | Número de accesión en el GenBank | ||
| ITS | EF | RPB2 | |||
| F. falciforme(FSSC) | FsDAG10 | CH2 | KX583230 | KY514171 | MF939148 |
| F. falciforme(FSSC) | FsDAG11 | CH1 | KX583231 | KY514185 | MF939149 |
| F. falciforme(FSSC) | FsDAG14* | CH2 | KX583232 | KY514172 | KY637066 |
| F. falciforme(FSSC) | FsDAG36 | CH2 | KX583233 | KY514173 | MF939150 |
| F. falciforme(FSSC) | FsDAG43* | CH1 | KX583234 | MF939167 | KY637067 |
| F. falciforme(FSSC) | FsDAG44* | CH1 | KX583235 | KY514176 | KY637068 |
| F. falciforme(FSSC) | FsDAG45 | CH1 | KX583236 | KY514177 | MF939151 |
| F. falciforme(FSSC) | FsDAG46 | CH1 | KX583237 | KY514187 | MF939152 |
| F. falciforme(FSSC) | FsDAG48 | CH1 | KX583238 | KY514178 | MF939153 |
| F. falciforme(FSSC) | FsDAG49 | CH1 | KX583239 | KY514179 | MF939154 |
| F. falciforme(FSSC) | FsDAG50 | CH1 | KX583240 | KY514180 | MF939155 |
| F. falciforme(FSSC) | FsDAG51 | CH1 | KX583241 | KY514181 | MF939156 |
| F. falciforme(FSSC) | FsDAG52 | CH1 | KX583242 | KY514182 | MF939157 |
| F. falciforme(FSSC) | FsDAG53 | CH1 | KX583243 | KY514183 | MF939158 |
| F. falciforme(FSSC) | FsDAG54 | CH1 | KX583244 | KY514184 | MF939159 |
| F. falciforme(FSSC) | FsDAG30 | CH1 | KX583245 | KY514186 | MF939160 |
| F. falciforme(FSSC) | FsDAG37 | CH2 | KX583246 | KY514174 | MF939161 |
| F. falciforme(FSSC) | FsDAG29 | CH1 | KX583247 | MF939168 | MF939162 |
| F. falciforme(FSSC) | FsDAG40* | VG1 | KX583248 | KY514175 | KY637065 |
| F. brachyigibbosum (FSSC)a | FbDAG41 | VG1 | KX583249 | MF939166 | MF939163 |
| F. brachyigibbosum(FSSC)a | FbDAG67* | CH1 | KX583250 | KY514170 | KY637062 |
| F. brachyigibbosum (FSSC)a | FbDAG8* | CH1 | KX583251 | KY514169 | KY637061 |
| F. brachyigibbosum(FSSC)a | FbDAG9 | CH2 | KX583252 | MF939165 | MF939164 |
| F. oxysporum f. sp. niveum (FOSC) | FoDAG38* | VG2 | KX583253 | KY514188 | KY637063 |
| F. oxysporum f. sp. niveum (FOSC) | FoDAG39* | VG1 | KX583254 | KY514189 | KY637064 |
| Ceratobasisium sp.AG-F | CeDAG3* | CH2 | MF804912 | KY637071 | |
| Ceratobasisium sp.AG-F | CeDAG5* | VG2 | KX583256 | - | KY637074 |
| Ceratobasisium sp.AG-F | CeDAG12* | CH2 | KX583257 | - | KY637072 |
| Ceratobasisium sp.AG-F | CeDAG19* | VG2 | KX583258 | - | KY637073 |
| Ceratobasisium sp. | CeDAG17 | CH1 | MF804910 | - | MF804913 |
| Ceratobasisium sp. | CeDAG22 | CH2 | KX583259 | - | - |
| Ceratobasisium sp. | CeDAG23 | CH2 | KX583260 | - | - |
| T. praticola=R. solaniAG-4 | RhDAG18* | CH2 | MF804911 | - | MF804914 |
| Thanatephorus sp. | RhDAG20* | VG1 | KX583262 | - | KY637069 |
| Thanatephorus sp. | RhDAG21* | VG1 | KX583263 | - | KY637070 |
*= Aislados considerados en el análisis filogenético. FSSC= Fusarium solani Species Complex. FSSCa= Fusarium sambucinum Species Complex. FOSC= Fusarium oxysporum Species Complex. CH1= Costa de Hermosillo1, CH2= Costa de Hermosillo2, VG1= Valle de Guaymas1, VG2= Valle de Guaymas. AG=Grupo anastomósico. - = sin dato
Pruebas de patogenicidad: La aparición de síntomas en las plantas ocurrió después de 14 días de su inoculación, en forma de lesiones en la raíz y base del tallo. Todos los aislados y sus combinaciones causaron la muerte de las plantas después de 21 días. El Cuadro 3 muestra el porcentaje de raíces infectadas en cada tratamiento. Las plantas testigo no presentaron síntomas.
Cuadro 3 Porcentaje de raíces infectadas en las pruebas de patogenicidad.
| Tratamiento | SS-7187 HQ F1 | Precious Petit |
| Testigo | 0 | 0 |
| F. solani | 23 | 66 |
| F. oxysporum | 71 | 75 |
| F. brachygibbosum | 87 | 100 |
| Rhizoctonia solani | 54 | 84 |
| Ceratobasidium | 48 | 100 |
| F. solani + F. brachygibbosum | 55 | 100 |
| F. oxysporum + F. brachygibbosum | 55 | 100 |
| R. solani + Ceratobasidium | 71 | 100 |
| F. solani + Ceratobasidium | 40 | 80 |
| F. oxysporum + Ceratobasidium | 100 | 100 |
DISCUSIÓN
Aun cuando el estado de Sonora es el principal productor de sandía en México, no se tienen reportes formales acerca de los hongos asociados a la pudrición de raíz. Los datos patológicos en esta investigación demuestran que individualmente o en conjunto, al menos cinco especies diferentes de hongos causan pudrición radicular y eventualmente la muerte de plantas de sandía antes de alcanzar su madurez fisiológica, al momento de la formación y desarrollo de frutos. Las características de las colonias, la morfología, la tinción de núcleos y el análisis filogenético de secuencias permitió identificar dos especies de la familia Ceratobasidiaceae: Rhizoctonia solani y Ceratobasidium sp. y tres del género Fusarium: F. falciforme, F. oxysporum y F. brachygibbosum, lo que demuestra que hay una comunidad diversa de hongos causando pudrición de raíz en sandía cultivada en la costa de Hermosillo y el Valle de Guaymas, Sonora.
La especie predominantemente encontrada fue F. falciforme, con 25 aislados provenientes de tres campos de los dos sitios muestreados. Inicialmente, este hongo fue identificado como la morfoespecie politípica F. solani. Con base en el análisis filogenético concatenado de las regiones ITS, TEF-1α y RPB2 usando MV se forma un clado separado dentro del Fusarium solani Species Complex (FSSC). Este complejo agrupa por lo menos a 60 diferentes especies que, por similitud en la morfología de sus conidias, son llamadas especies crípticas. Cuentan con un amplio rango de hospederas y han sido subdvididas en formae speciale, dependiendo de la especificidad al hospedero (O’Donnell et al., 2015). Análisis filogenéticos recientes han revelado que cada formae speciale corresponde a una especie, biológica y filogenéticamente distinta (Coleman, 2016; O’Donnell et al., 1998). F. falciforme ha sido reportado como agente causal de marchitamiento y pudrición de raíz en frijol lima y garbanzo en Brasil (Sousa et al., 2017, Cabral et al., 2016).
Se identificaron cinco aislados de F. brachygibbosum perteneciente a Fusarium sambucinum Species Complex (FSSC) en tres sitios de muestreo. Las características de las colonias coinciden con la primera descripción publicada (Padwick, 1945), presenta micelio aéreo abundante de blanco a rosa, esclerocios de blanco a ámbar de hasta 2.0 mm de diámetro, conidias ovoides a fusiformes, macroconidias hiperbólicamente curveadas, clamidosporas terminales o intercaladas, sencillas o en cadenas, generalmente de una célula. Como producto preliminar de esta investigación, F. brachygibbosum fue reportado por primera vez como agente patógeno, causante de marchitez en plantas de sandía (Rentería-Martínez et al., 2015). Recientemente se ha registrado como agente causal de podredumbre del tallo del maíz (Shan et al., 2017), marchitez y muerte regresiva en Euphorbia larica y olivo (Al-Mahmooli et al., 2013; Trabelsi et al., 2017) y de podredumbre y cancros en almendro y nogal (Stack et al., 2017; Marek et al., 2013).
F. oxysporum f. sp. niveum perteneciente al Fusarium oxysporum Species Complex (FOSC), se aisló solamente de los campos ubicados en el Valle de Guaymas. Los miembros de FOSC causan marchitez vascular y pudrición radicular en más de 100 diferentes especies vegetales, y en base a su especificidad con el hospedero se han reportado más de 80 formae speciale. En ese sentido un diagnóstico certero, aún antes de que se presenten sus síntomas, es crucial para el manejo de los cultivos (López-Mondéjar et al., 2012).
En lo que respecta a la familia Ceratobasidiae, Rhizoctonia binucleada (teleomorfo: Ceratobasidium sp.) fue la especie de más amplia distribución, ya que se aisló de todos los campos muestreados, y más frecuentemente en el campo No. 2 de la Costa de Hermosillo. Los síntomas observados en raíces y tallos de plantas infectadas fueron lesiones localizadas café-rojizas y ligeramente hundidas en la base del tallo y de 0.2 a 2.0 cm de largo. En algunos casos, la decoloración presentada por las plantas enfermas afectó casi el 90 % de sistema radicular (Meza-Möller et al., 2014).
El análisis filogenético mediante MV mostró que los aislados obtenidos de Ceratobasidium sp., corresponden a dos grupos anastomósicos distintos: AG-A y AG-F, previamente registrados en plantas enfermas de Ipomoea batatas y Arachis hypogaea, respectivamente. Se ha demostrado que las relaciones filogenéticas de hongos de Rhizoctonia demuestran consistencia entre los clados formados y los grupos anastomósicos a los que pertenecen (González et al., 2016).
Es importante señalar que la mayoría de los aislados de Ceratobasidium sp. encontrados en esta investigación pertenecen al grupo AG-F, lo que coincide con trabajos previos sobre pudrición radicular en sandía realizados en Arizona, EUA (Nischwitz et al., 2013) e Italia (Aiello et al., 2012). Ceratobasidium AG-F también es el agente causal de pudrición radicular en fresa (Sharon et al., 2007), Tagetes erecta (Saroj et al., 2013) y pistache (Alaei et al., 2017).
No existe información previa acerca de la patogenicidad del grupo anastomósico AG-A en sandía, pero sí en remolacha azucarera y manzano (Wang y Wu, 2012).
Los aislados de menor frecuencia fueron los pertenecientes al género Rhizoctonia, que por la alta variabilidad en su distribución geográfica, morfología, especificidad de hospederas, y patogenicidad se ha propuesto también como un complejo de especies (González et al., 2006). El aislado RhDAG18 obtenido del sitio CH2 pertenece al AG-4 y ha sido delimitado morfológicamente como Thanatephorus praticola (Mordue et al., 1989), lo que fue corroborado posteriormente por el análisis filogenético multilocus (González et al., 2016). T. praticola se ha presentado en asociación con Ceratobasidium sp. AG-F como parte de un complejo de hongos que causan pudrición de raíz y declive de plantaciones de sandía en etapa de producción en Italia (Aiello et al., 2012).
Las pruebas de patogenicidad demostraron que los hongos aislados son agentes causales de pudrición radicular en sandía. Los resultados del porcentaje de raíces infectadas evidenciaron que las dos variedades de sandía evaluadas son susceptibles a los cinco patógenos identificados, inoculados por separado o en combinaciones. A este respecto, Aiello et al. (2012) detectaron porcentajes de incidencia de la enfermedad superiores a 81 %, al evaluar 6 diferentes aislados de Ceratobasidium del grupo anastomósico AG-F.
Debido a las pérdidas económicas derivadas de enfermedades radiculares en las regiones estudiadas, la siembra directa en suelo ha sido sustituida casi en su totalidad por sandía injertada sobre patrones resistentes a pudrición de raíz. Los portainjertos comúnmente empleados en la región son híbridos de Cucurbita maxima x C. moschata, sin embargo, estos híbridos ya han mostrado susceptibilidad a pudrición de raíz (López-Elías et al., 2010). Algunos portainjertos han mostrado susceptibilidad a Fusarium en España, principal productor de cucurbitáceas en Europa (Armengol et al., 2000). Por otro lado, la aparente resistencia a marchitez por Fusarium de algunas variedades comerciales de sandía, es dependiente del tipo y concentración inicial del inóculo (Martyn y McLaughlin, 1983); en ese sentido, la oportuna identificación del patógeno permitirá una mejor elección del portainjerto. Adicionalmente, una vez determinada con precisión la especie patogénica presente, la cuantificación de su concentración, permitiría un mejor manejo de la enfermedad. Mediante la técnica de PCR cuantitativa (qPCR), es posible detectar hasta 1 pg de ADN fúngico en plantas asintomáticas (Haegi et al., 2013).
CONCLUSIONES
En el presente estudio se reporta por primera vez a Fusarium falciforme y Thanatephorus praticola como causantes de pudrición radicular en plantas de sandía en México. El análisis multilocus permitió corroborar también a Fusarium brachygibbosum y Ceratobasidium sp., previamente reportados. Esta investigación, representa un primer intento para generar información que conduzca a estudios más amplios, que permitan establecer estrategias adecuadas para el manejo de la enfermedad en plantaciones comerciales de sandía.

