











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkLos manzanos (Malus x domestica Borkh.) son uno de los cultivos templados más importantes en México (Ramírez-Legarreta et al., 2011). Los árboles de manzano son afectados por diversos factores bióticos y abióticos que limitan su producción, especialmente las enfermedades de raíz. En los agroecosistemas, el incremento en la diversidad de microorganismos patogénicos (Oomicetos, hongos, bacterias, nematodos, entre otros), pueden conducir a las enfermedades de pudrición de raíz. Las enfermedades de raíz causadas por hongos y Oomicetos en árboles de manzano, representan un importante problema económico alrededor del mundo. Estos microorganismos destruyen y colapsan el sistema radicular de portainjertos susceptibles, reduciendo la absorción de agua y nutrientes, bloqueando los haces vasculares y eventualmente, causando la muerte de los árboles de manzano (Rumberger et al., 2007; Samaniego-Gaxiola, 2007; Tewoldemedhin et al., 2011a, b). Los principales patógenos del manzano incluyen a Phytophthora spp., Pythium spp., Phymatotrichopsis omnivora, Fusarium spp., Rhizoctonia spp., entre otros (Latorre et al., 2001; Samaniego-Gaxiola, 2007). Sin embargo, en muchas áreas productoras de manzana, todavía hay patógenos que no han sido identificados como agentes de enfermedades para los manzanos. Las principales enfermedades en árboles de manzano, incluyen las enfermedades de pudrición de cuello y corona, resultado de un complejo de patógenos (Lamichhane y Venturi, 2015). La incidencia, agentes causales y patogenicidad de agentes causales de enfermedades de pudrición de cuello y raíz, han sido pobremente estudiados durante las últimas tres décadas en huertos de manzano del norte de México. Se requiere información en este sentido, en particular en ésta área productora de manzana, para desarrollar un mejor manejo integrado de enfermedades, con el fin de reducir el uso excesivo de plaguicidas químicos de amplio espectro para el control de estas enfermedades. Indudablemente, estas mejoras en el manejo integrado de enfermedades incluyen el uso de microorganismos con actividad antagonista, los cuales son colonizadores eficientes, producen metabolitos que inhiben el crecimiento de fitopatógenos y actúan como promotores del crecimiento vegetal (Ezziyyani et al., 2004). Por lo tanto, el objetivo del presente estudio fue estimar la incidencia de patógenos de la raíz en huertos de manzano de Chihuahua, México, identificando los hongos y Oomicetos probablemente involucrados en las enfermedades y evaluando su patogenicidad en portainjertos de manzano comerciales. También se determinó el efecto antagonista in vitro de algunos microorganismos contra algunos de los Oomicetos identificados.
MATERIALES Y MÉTODOS
Sitio experimental y estimación de incidencia de daño infeccioso
La incidencia de raíces con daño infeccioso se determinó una vez, en cinco huertos de las principales regiones productoras de manzana de Chihuahua, México (Cuahtémoc, Bachíniva, Namiquipa y Guerrero, Cuadro 1) durante junio y julio del 2013. Las condiciones climáticas y el nivel de tecnificación variaron entre las áreas evaluadas. Quinientos árboles, distribuidos en 10 hileras, se evaluaron en cada huerto (10,000 árboles en total) en un diseño completamente al azar (Cuadro 1). Los síntomas de daño infeccioso de raíz considerados fueron marchitez, pobre crecimiento de los brotes, follaje pálido o amarillo, follaje necrótico, defoliación prematura, muerte regresiva de hojas y brotes terminales, manchas negras o cafés en los tallos, formación de cánceres y crecimiento irregular.
Cuadro 1 Huertos de manzano muestreados, ubicación geográfica, portainjertos plantados, edad de los árboles e incidencia de árboles de manzano con posible enfermedad de raíz en cuatro localidades del estado de Chihuahua, México.
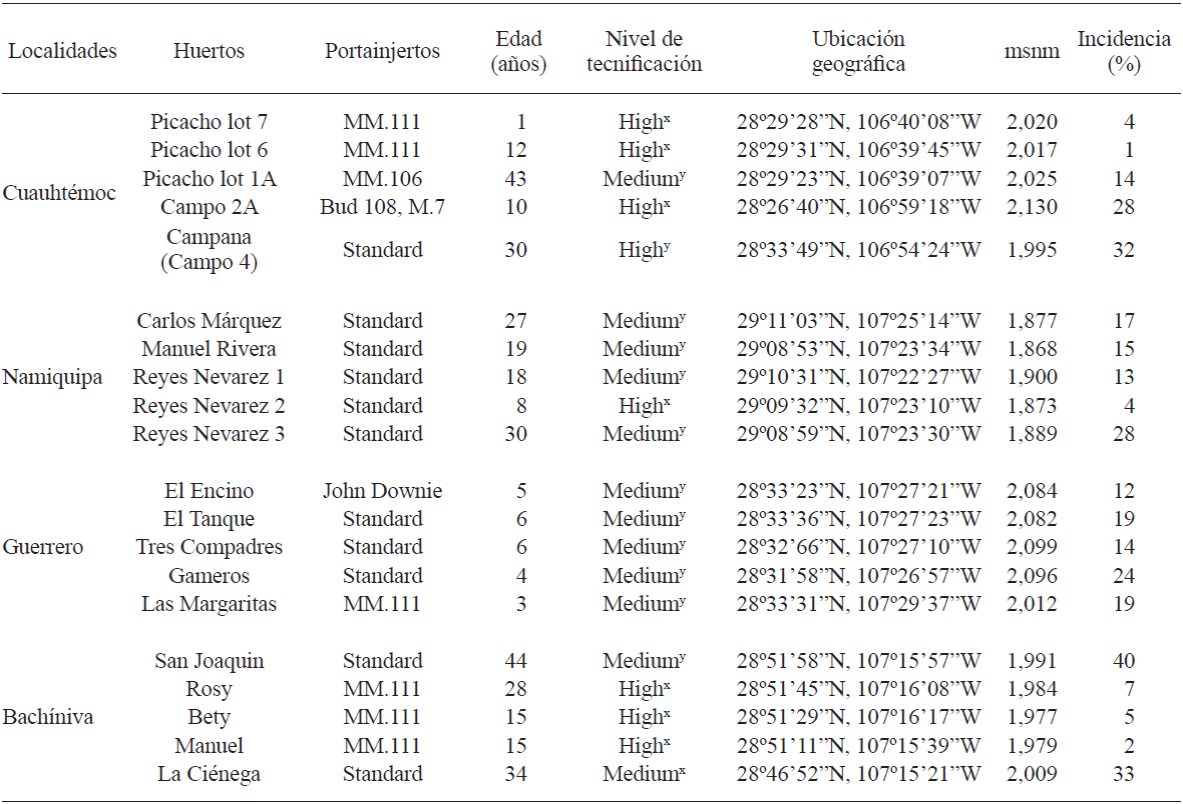
msnm: metros sobre el nivel del mar
x Alto: Irrigación por micro-aspersión, calefacción, malla antigranizo; y Medio: Riego rodado, calefacción, sin malla antigranizo, sin límite del uso de recursos y asistencia técnica ocasional (Ramírez-Legarreta et al., 2004)
Muestras de suelo y tejido
Se recolectó tejido enfermo (muestras de raíz y tallo de 10-20 cm de largo) y suelo cercano a la rizosfera (500-600 g) de 10 árboles con aparente daño infeccioso de raíces y de árboles libres de síntomas, de cada huerto para aislar los hongos y Oomicetos.
Aislamiento y purificación de microorganismos
Se lavaron con agua corriente tres a cinco piezas (10-20 cm de largo) de tejido enfermo para eliminar el exceso de suelo y subsecuentemente se cortaron en piezas de 1 cm. Los pedazos se lavaron por 1 min con una solución de hipoclorito de sodio (NaOCl) 0.5%, se enjuagaron tres veces con agua destilada y se secaron en papel estraza estéril en una cabina de bioseguridad (Envirco Corporation, Albuquerque, New Mexico, EUA). Las submuestras de tejido (5 piezas de 1 cm de largo) se colocaron por triplicado en cámaras húmedas (cajas de Petri de 90-mm con algodón húmedo). Por otra parte, submuestras de tejido (cinco piezas de 3-5 mm de largo) se colocaron por triplicado en medios de cultivo semi-selectivos V8-agar [jugo V8, carbonato de calcio (CaCO3; Sigma-Aldrich) - agar (Sigma-Aldrich)] y papa-dextrosa-agar (PDA; BD Bioxon). Estos dos medios contenían antibióticos [pimaricina 0.1 g/L (Sigma-Aldrich, EUA), rifampicina 0.01 g/L (Sigma, EUA), oxytetraciclina 0.03 g/L (Sigma, Alemania)] (Tewoldemedhin et al., 2011a). Por lo tanto, se evaluaron 15 submuestras de 1 cm y 15 submuestras de 3-5 mm de cada árbol con y sin síntomas de daño infeccioso. Todos los ensayos se llevaron a cabo a 28 °C por 5 d en una cámara ambiental sin luz (Precision Scientific, Winchester, VA, EUA). Los hongos fueron purificados en medio PDA usando la técnica de cultivo monospórico, seguido por una incubación a 28 °C por 72-96 h. Para el aislamiento de Oomicetos, se usó el medio PARPH [17 g/L de medio de harina de maíz (Analytical Fluka, Sigma); 0.10 g/L pentacloronitrobenceno (PCNB), 0.27 g/L ampicilina (Sigma) y 0.01 g/L rifampicina (Sigma)]. Estos microorganismos fueron incubados bajo las mismas condiciones que los hongos.
Las especies de Trichoderma se aislaron por diluciones seriadas (1:10) de suelo tamizado (1 g) de árboles con y sin síntomas de daño infeccioso en tubos de ensayo con 9 mL de agua peptonada estéril (0.1% pepetona y 0.85% NaCl en agua destilada). Se sembraron por triplicado alícuotas (50 µL) de las diluciones (104, 105, and 106) por la técnica de difusión en cajas de Petri de 90-mm que contenían medio PDA adicionado con antibióticos indicados previamente (Schoenborn et al., 2004).
Identificación morfológica y molecular de microorganismos
Los hongos y Oomicetos fueron purificados en medio PDA sin antibióticos (Tewoldemedhin et al., 2011a) e identificados a nivel de género utilizando claves taxonómicas (Barnett y Hunter, 1972; Dugan, 2006; Watanabe, 2010), de acuerdo con sus caracteres morfológicos observados en un microscopio óptico (Carl Zeiss, Jena, Alemania). La identidad de estos microorganismos fue confirmada molecularmente.
El ADN genómico (ADNg) fue extraído y usado para llevar a cabo el análisis de identidad de los aislados fúngicos, basados en sus caracteres moleculares. Para este propósito, un explante del hongo purificado se colocó en una caja de Petri que contenía medio PDA, se cubrió con celofán estéril y se incubó a 28 °C por 7 d. Se recolectó el micelio y se colocó en un mortero de porcelana con un amortiguador (200 mM Tris-HCl (pH = 8), 250 mM NaCl, 25 mM EDTA, 0.5% SDS) a 70 °C. El micelio se maceró de acuerdo con Raeder y Broda (1985). El ADNg obtenido se visualizó por electroforesis en un gel de agarosa al 1% y subsecuentemente se usó para amplificar el espaciador transcrito interno (ITS4 y 5, por sus siglas en inglés) del ADNr, usando los cebadores universales ITS5 (5’-GGAAGTAAAAGTCGTAACAAGG-3’) e ITS4 (5’-TCCTCCGCTTATTGATATGC-3’) (White et al., 1990). Las condiciones de amplificación incluyeron una etapa inicial de desnaturalización a 94 °C por 5 min, 30 ciclos de desnaturalización a 94 °C por 30 s, alineamiento a 60 °C por 30 s, extensión a 72 °C por 45 s, y una etapa final de extensión a 72 °C por 10 min.
Los productos de PCR se examinaron por electroforesis en un gel de agarosa al 1% y subsecuentemente purificados usando el kit DNA Clean & Concentrator (Zymo Research, California, EUA), de acuerdo con las instrucciones del fabricante. Estos productos fueron secuenciados en Macrogen (Rockville, MD, EUA). Las secuencias obtenidas fueron comparadas contra las bases de datos del NCBI usando el algoritmo de BLAST (Altschul et al., 1990) para verificar el porcentaje de identidad correspondiente a las especies identificadas. Las secuencias de nucleótidos obtenidas en el presente estudio no han sido depositadas en el NCBI.
Pruebas de patogenicidad en portainjertos de manzano
Los postulados de Koch se realizaron bajo condiciones de invernadero utilizando 15 patógenos (ocho hongos y siete Oomicetos) aislados de la rizosfera de árboles de manzano con y sin síntomas de daño infeccioso en raíz. Se adquirieron árboles de manzano de 1 año de edad de los 12 portainjertos más plantados en México (G.30, G.41, G.202, G.222, G.935, M.7, M.25, MM.106, MM.111, Bud 118, Bud 9 y Estándar), de dos viveros en Cuauhtémoc, Chihuahua (“Viveros Sacramento” y “Vivero Los Cienes”). Se inocularon cinco árboles de cada portainjerto con cada patógeno. Adicionalmente, se utilizaron cinco portainjertos como grupo testigo (sin inóculo).
Los árboles se plantaron en bolsas negras de polietileno (5 L) conteniendo sustrato previamente esterilizado a 121 °C y 15 psi. El sustrato estaba compuesto por suelo franco, vermiculita y peat moss (1: 1: 1). Dos meses después del trasplante, los árboles fueron inoculados con 10 mL de inóculo de hongos u Oomicetos sin cuantificar (Hantula et al., 2000). El inóculo tenía 3 d edad y fue crecido en caldo vegetal V8 [jugo V8 (Campbell´s ™) y carbonato de calcio (CaCO3; Sigma Aldrich)] a 28 °C en un agitador orbital a 140 rpm (Orbit 1900, Labnet International Inc.). Los portainjertos inoculados se mantuvieron por otros dos meses bajo condiciones de invernadero (Uthkede y Smith, 1991; Latorre et al., 2001). Durante este tiempo, se monitorearon semanalmente los síntomas característicos resultado de las infecciones de hongos y Oomicetos. Se consideraron como patogénicos a los aislados de hongos y Oomicetos que causaron la muerte de los árboles o que causaron síntomas severos de daño en las raíces, tallos, hojas y brotes después de 2 meses de la inoculación.
Actividad antagónica in vitro
Se confrontaron cuatro aislados de hongos y dos de bacterias antagonistas contra siete aislados de Oomicetos. Se incluyeron los antagonistas Trichoderma asperellum y dos especies de Bacillus previamente identificados (Rios-Velasco et al., 2016).
Se evaluó la actividad in vitro de los aislados de Trichoderma spp. y Bacillus contra los Oomicetos obtenidos de huertos de manzano. Para este propósito, se realizaron confrontaciones (Trichoderma spp. vs Oomicetos) usando la técnica de cultivo dual en cajas de Petri (90 mm) conteniendo Papa-Dextrosa-Agar (PDA). Se colocó un disco de papel filtro (6 mm) conteniendo el micelio y conidias sin cuantificar del patógeno en un lado de la caja y se colocó otro disco de papel filtro conteniendo el micelio y conidias sin cuantificar del antagonista, en el lado opuesto de la caja. Las confrontaciones de Bacillus spp. vs Oomicetos, se realizaron colocando un circulo de papel filtro de 6 mm de diámetro del patógeno (micelio y conidias sin cuantificar) en el centro de la caja Petri conteniendo PDA, mientras que la bacteria fue inoculada, el mismo día que el fitopatógeno, en los cuatro puntos cardinales de la caja de Petri usando círculos de papel filtro (6 mm de diámetro con inóculo de 48 h de edad) (Figura 1).
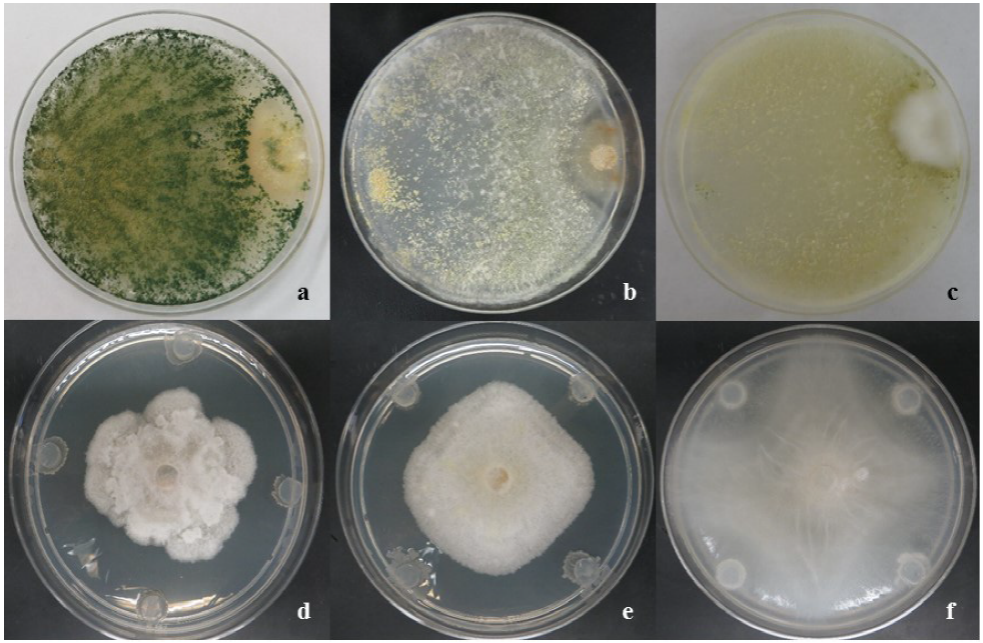
Figura 1 Micoparasitismo de especies de Trichoderma (a-c); a) T. asperellum vs Pythium ultimum; b) T. harzianum vs Pythium sp.; c) T. atroviride vs Phytophthora cactorum; y halos de inhibición de especies de Bacillus (d-f); d) B. amyloliquefaciens vs Phytophthora cactorum; e) B. amyloliquefaciens vs Phytophthora cactorum; f) B. methylotrophicus vs Pythium ultimum al sexto día post-inoculación in vitro.
Tanto los testigos de patógenos y antagonistas se colocaron individualmente en el centro de la caja de Petri y se realizaron por triplicado con 10 unidades experimentales cada una. Las cajas se incubaron a 28 °C en una incubadora Precision Scientific (Modelo 31534) y se realizaron mediciones sistemáticas cada 24 h por ocho días, del crecimiento radial de las colonias de hongos confrontadas con los hongos y bacterias antagonistas y las cajas testigo, respectivamente.
Se evaluó la posible actividad antagonista de Trichoderma spp. y Bacillus spp. en términos de crecimiento radial del patógeno (CRP), crecimiento radial del antagonista (CRA) y el porcentaje de inhibición del crecimiento radial (PICR). Se evaluó el PICR de acuerdo con Ezziyyani et al. (2004) usando la fórmula PICR = (R1 - R2) / R1 × 100, donde R1 es el crecimiento radial del patógeno testigo y R2 es el crecimiento radial del patógeno en la confrontación, mientras que el tipo de antagonismo fue medido de acuerdo con la escala de Bell (Bell et al., 1982).
Análisis estadístico
Ambos ensayos se realizaron por triplicado, con diez cajas de Petri por repetición para cada evaluación in vitro de bacteria-hongo, usando un diseño completamente al azar con los siguientes seis tratamientos: cuatro aislados de hongos antagonistas y dos aislados de bacterias antagonistas confrontadas contra siete aislados de Oomicetos, con un testigo para cada microorganismo, mientras que cada tratamiento constó de un microorganismo antagonista con 30 cajas de Petri. Los datos del PICR y el halo de inhibición se analizaron usando el Sistema de Análisis Estadístico versión 9.0 (SAS, 2002) para un análisis de varianza (ANVA), y las medias fueron separadas usando la prueba de Tukey (p = 0.05).
RESULTADOS Y DISCUSIÓN
Incidencia de daño infeccioso en raíces
La incidencia total de enfermedades de raíz en los huertos muestreados fue de 17%. Dicha incidencia osciló de 1 a 32% y de 4 a 28% en huertos de Cuauhtémoc y Namiquipa, respectivamente, con una incidencia promedio de 16% para ambas áreas. En Guerrero y Bachíniva, la incidencia fluctuó de 12 a 24% y de 2 a 40%, respectivamente, con una incidencia promedio de 17% para ambas áreas (Cuadro 1). La incidencia más alta se observó en los huertos “San Joaquín” y “La Ciénega”, localizados en Bachíniva, probablemente como consecuencia de la avanzada edad de los árboles (44 y 34 años, respectivamente) y el nivel medio de tecnificación, haciéndolos más susceptibles al ataque de los fitopatógenos. Las condiciones climáticas de ésta región, las cuales son mejores para el crecimiento de los microorganismos, también pueden estar involucradas en esta alta incidencia de daño infeccioso de raíces.
Los valores de incidencia probablemente dependen del nivel de tecnificación (principalmente el método de riego), las condiciones climáticas, el tipo de suelo, las interacciones sin reconocer entre patógenos, el portainjerto, las diferencias en las prácticas hortícolas y la edad de los árboles en cada huerto. Además, es importante considerar que en algunos huertos, los agricultores aplican materia orgánica al suelo, lo que puede favorecer el establecimiento de microorganismos antagonistas, mejorando su desarrollo y, por lo tanto, evitando el crecimiento de fitopatógenos (Kamal et al., 2010). La variabilidad de los valores de incidencia para cada huerto y localidad, pueden ser consecuencia de la patogenicidad y virulencia de los microorganismos (Wilcox, 1993). Manici et al. (2013) también observaron que la incidencia de enfermedades de raíces varió considerablemente entre regiones y la severidad de las enfermedades se atribuyó a la composición del complejo de hongos.
Las enfermedades de pudrición de corona y raíz en huertos de manzano son problemas importantes alrededor del mundo. Manici et al. (2013) observaron una alta incidencia de infecciones causadas por Pythium spp. y las infecciones causadas por especies de Fusarium fueron altamente asociadas a enfermedades de replante. Este problema también se observó en el presente estudio para los portainjertos de manzano de 1 año de edad (MM.111) en “El Picacho Lot 7” huerto de Cuauhtémoc, con un alto nivel de tecnificación. De acuerdo con Ramírez-Legarreta et al. (2004), el nivel de tecnificación (medio y alto) en huertos de manzano, afectan directamente la incidencia de enfermedades, principalmente mediante el tipo de micro aspersión o riego rodado, pero también del tipo de calefacción, uso de malla anti-granizo, uso de insumos y asistencia técnica continua u ocasional, también están involucradas, entre otros factores. Sin embargo, la mayoría de los huertos muestreados en el presente estudio, con un nivel medio de tecnificación, mostraron la presencia de P. cactorum y Pythium spp. y particularmente el hongo F. oxysporum, lo cual, en algunos casos, se puede atribuir al tipo de manejo del huerto, siendo un factor determinante potencial en la incidencia de los patógenos, incluyendo los agentes causales de enfermedades de cuello y raíz. Es importante destacar que los huertos con un alto nivel de tecnificación, cuentan con asistencia técnica permanente, la cual ayuda en la detección temprana de enfermedades para su prevención y manejo.
Identificación morfológica y molecular de microorganismos
Los microorganismos asociados con los síntomas de enfermedades de raíz en huertos de manzano fueron los Oomicetos Phytophthora spp. y Pythium spp. y los hongos Fusarium spp, Gibberella spp. (teleomórfo de Fusarium spp.), Bionectria ochroleuca, Clonostachys sp., Alternaria spp., y Phymatotrichopsis sp. Estos microorganismos fueron identificados de acuerdo con sus caracteres morfológicos y secuencias genómicas. El género Fusarium spp., fue el más frecuentemente observado (67.3%, 202 aislados), con 32.3% (97 aislados) de este género, pertenecientes a F. oxysporum. Las especies restantes de éste género fueron F. solani, F. subglutinans, F. sacchari, F. succisae, F. tricinctum, F. fujikuroi, F. proliferatum y F. equiseti. Ésta distribución de especies del género Fusarium es similar a lo encontrado en otras regiones geográficas alrededor del mundo (Serdani et al., 2002; Tewoldemedhin et al., 2011a). Leslie et al. (2007) encontraron que F. succisae está filogenéticamente relacionado con F. proliferatum, la cual ha sido asociada con plántulas de manzano en China (Ju et al., 2014). Tanto F. fujikuroi y F. sacchari, fueron asociadas con arroz, caña de azúcar y parcelas de maíz de la Península de Malasia (Ju et al., 2014), mientras que F. equiseti ha sido asociada con la rizosfera de árboles de manzano en Chihuahua, México (Pérez-Corral et al., 2015). Además, Carreri et al. (2013) aislaron F. tricinctum de raíces y bulbos de cebolla en el sur de Italia. Sin embargo, no se ha reportado una asociación entre las especies de Fusarium y la rizosfera de árboles de manzano. Por lo tanto, se considera que es la primera vez que se reportan a F. fujikuroi, F. sacchari, F. tricinctum and F. subglutinans como patogénicos para las raíces de manzano.
El segundo género más frecuentemente observado fue el teleomórfo de Fusarium (Gibberella spp.), con 15% (45 aislados pertenecientes a G. moniliformis anamórfo: Fusarium verticillioides y G. intermedia anamórfo: Fusarium proliferatum), seguidos por Bionectria ochroleuca (12%, 36 aislados), Alternaria spp. (17%, 5 aislados) que comprenden principalmente A. brassicae y A. alternata. Clonostachys sp. (1.3%, 4 aislados), Phytophthora cactorum (1.3%, 4 aislados), Pythium spp. (0.7%, 3 aislados), Phymatotrichopsis sp. (0.3%, 1 aislado) (Cuadro 2). Scherm et al. (2013) demostraron que el teleomórfo de las especies de Fusarium (Gibberella spp.), puede ser encontrado en otros cultivos. Además, los Oomicetos P. cactorum y Pythium spp. encontrados en este estudio, fueron distribuidos diferencialmente entre los huertos. Estas especies han sido reportadas como los principales agentes causales de pudriciones de cuello, corona y raíz en manzanos en otros países (Roiger y Jeffers, 1991; Yao et al., 2006). Otras especies tales como B. ochroleuca, A. alternata y A. brassicae, también han sido asociadas con pudriciones en árboles de manzano en Sudáfrica (Serdani et al., 2002). Por otra parte, P. omnivora, el agente causal de la pudrición texana de la raíz, ha sido reportado como patógeno que causa pérdidas económicas importantes en diferentes cultivos en el norte de México y el sur de Estados Unidos de América, particularmente en algodón y nogales, reflejando su adaptabilidad a una amplia gama de condiciones climáticas (Samaniego-Gaxiola, 2007), y siendo un potencial agente causal de enfermedades de raíz en otros árboles frutales. El presente estudio reporta por primera vez el hongo Phymatotrichopsis sp., como agente patógeno de raíces en huertos de manzano de México. Además se obtuvieron cuatro especies de Trichoderma de muestras de suelo, siendo Trichoderma gamsii el antagonista más frecuentemente observado con 72.5%, seguido por T. hamatum, T. harzianum y T. atroviride con 15, 7.5 y 5%, respectivamente (Cuadro 2).
Cuadro 2 Aislados de hongos y Oomicetos y posibles hongos antagonistas, identificados molecularmente, asociados a árboles de manzano con aparente enfermedad de raíz y libres de síntomas de cuatro localidades en el estado de Chihuahua, México, y su ocurrencia (%).
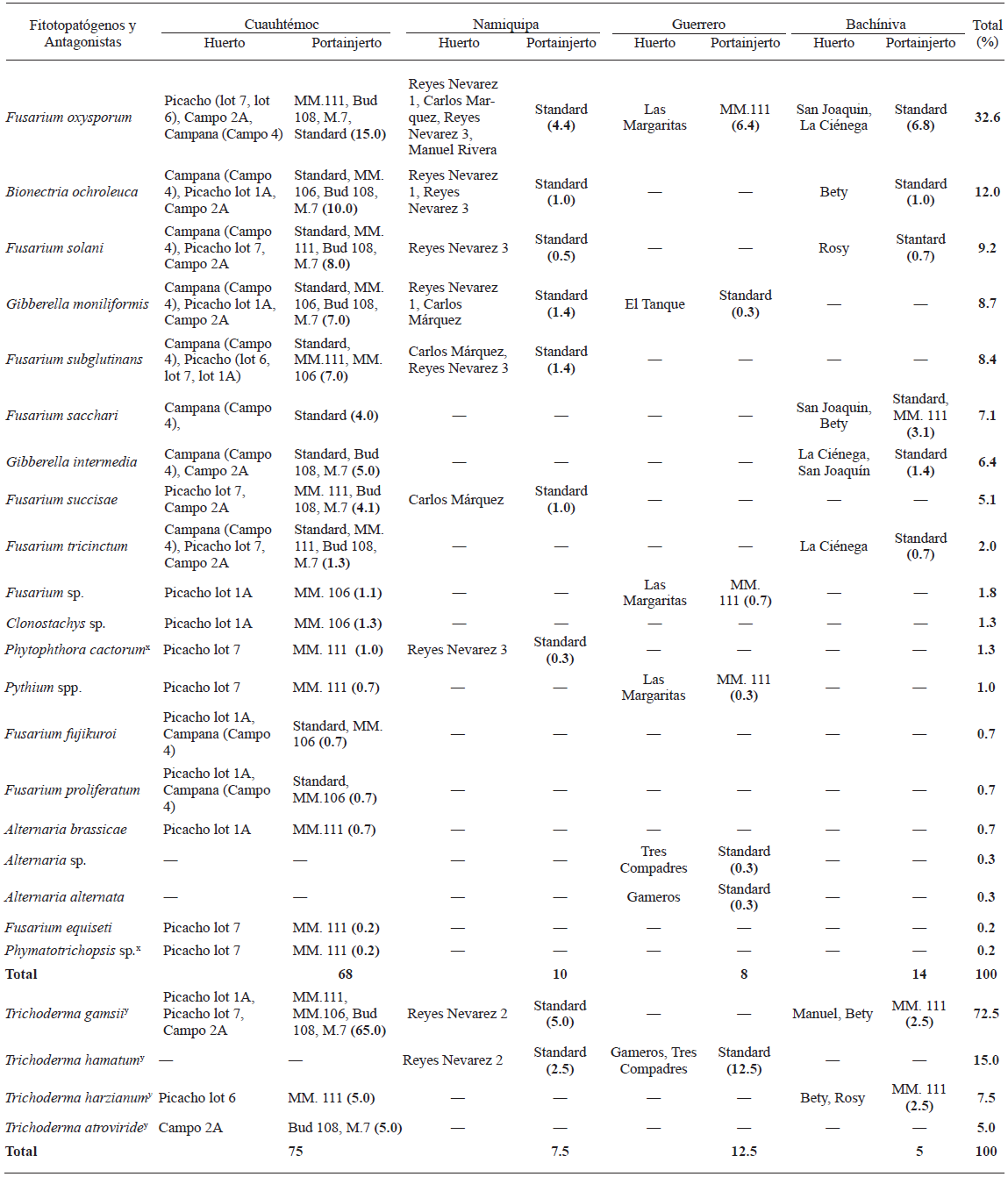
x Aislados de Oomicetos y hongos identificados de acuerdo con sus caracteres morfológicos
y Posibles hongos antagonistas
Pruebas de patogenicidad
En el Cuadro 3, se muestra la susceptibilidad de los portainjertos a hongos y Oomicetos. Todos los portainjertos fueron susceptibles al menos a dos de los aislados, excepto G.935, el cual fue resistente a los aislados de hongos y Oomicetos probados. Los portainjertos G.30, G.202 y G.41 fueron los más susceptibles a 13, 10 y 9 de los patógenos evaluados, respectivamente. Pythium ultimum fue el más patogénico, causando enfermedades en el 100% de los árboles probados (Cuadro 3). Aunque los portainjertos G.30 y MM.111 fueron altamente susceptibles (más de 50-75%), son ampliamente plantados en las áreas productoras de manzana de Chihuahua, México. Estos portainjertos mostraron síntomas de infección a cinco aislados de Oomicetos, siendo P. cactoum C1 y Pythium ultimum los aislados más patogénicos. Estos portainjertos junto con MM.106 han sido reportados como los más susceptibles a estos patógenos (Roiger y Jeffers, 1991; Uthkede y Smith, 1991; Latorre et al., 2001). Resultados similares fueron encontrados por Latorre et al. (2001) después de la inoculación de aislados de Phytophthora, siendo P. cactorum el más virulento y patogénico a los portainjertos M.9, M.25, MM.106 and MM.111. El portainjerto Estándar únicamente fue susceptible a dos especies de Pythium, P. irregulare y P. ultimum con 80 y 60%, respectivamente (Cuadro 3). Éste hallazgo es contrario a lo que se observó en los huertos evaluados, donde la más alta incidencia de daño infeccioso en raíces fue encontrado en portainjertos Estándar. Sin embargo, esto puede estar relacionado a la edad de los árboles (30-40 años), haciéndolos más susceptibles al ataque de fitopatógenos. La mayoría de los portainjertos probados fueron susceptibles a aislados de P. cactorum y Pythium spp., como fue demostrado previamente por otros (Yao et al., 2006). Roiger y Jeffers (1991) encontraron que P. cactorum fue altamente responsable de la pudrición de corona y raíz en árboles de manzano alrededor del mundo. Sin embargo, Tewoldemedhin et al. (2011a-b) reportaron que el agente etiológico de las enfermedades de raíz es un complejo múltiple de especies de hongos y Oomicetos pertenecientes a los géneros Phytophthora, Pythium y Rhizoctonia (Manici et al., 2013). En el presente estudio, P. cactorum, P. ultimum, F. sacchari y F. oxysporum fueron las especies más patogénicas. Estas especies han sido previamente reportadas en otros estudios como agentes causales de enfermedades de pudrición de raíz en árboles de manzano de mayor importancia alrededor del mundo (Uthkede y Smith, 1991; Latorre et al., 2001).
El aislado más patogénico de Fusarium fue F. sacchari, el cual infectó 100% de los árboles M.25, Bud.9 y G.30, a pesar de que este aislado tuvo una baja consistencia (7.1%) en comparación con el resto de los microorganismos identificados. Este efecto de F. sacchari puede ser consecuencia de su baja tasa de crecimiento o a la presencia de algunos aislados antagonistas coexistiendo en el mismo microambiente (Cuadro 3). En contraste, F. tricinctum y F. solani, resultó ser no-patogénico en siete portainjertos mientras el resto de los aislados de Fusarium mostró una baja patogenicidad. Manici et al. (2003) también observaron aislados no-patogénicos en árboles de manzano.
Cuadro 3 Porcentaje de patogenicidad de aislados de Oomicetos y hongos al ser evaluados bajo condiciones de invernadero contra doce portainjertos de 1 año de edad predominantemente plantados en el estado de Chihuahua, México, después de dos meses de ser inoculados.
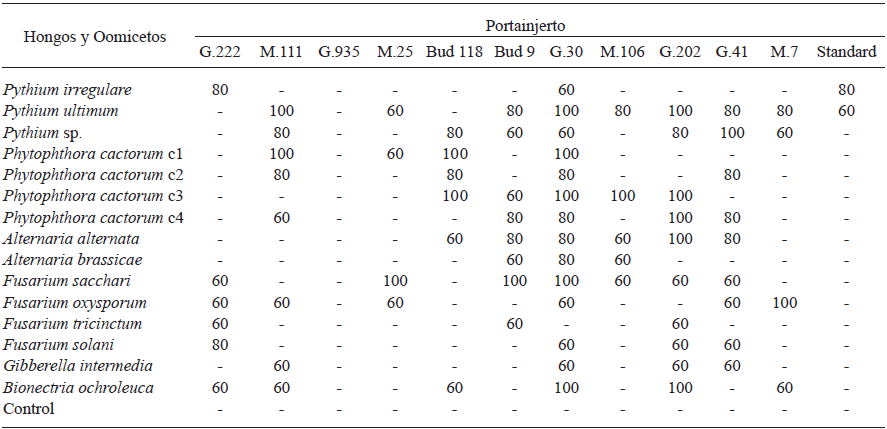
Solo se consideraron patogénicos a los aislados de Oomicetos y hongos que causaron síntomas en ≥ 60% de cada uno de los portainjertos, después de dos meses de haber sido inoculados
(-) Sin síntomas
Alternaria alternata fue el más patogénico en portainjertos G.202. Los portainjertos testigo se mantuvieron libres de enfermedades durante el experimento. Los principales síntomas observados fueron malformaciones, marchitez gradual en los portainjertos enfermos, defoliación, senescencia, tejido necrótico, clorosis, hojas rígidas, hojas con manchas redondas café oscuro con anillos concéntricos amarillos, brotes jóvenes marchitos, pudrición de cuello y raíz, tallos colapsados con manchas ovales negras, cánceres, decoloración de marrón a marrón rojizo y exudados gomosos.
Actividad antagónica in vitro de Trichoderma spp. vs Oomicetos
Las cuatro especies de Trichoderma mostraron un comportamiento antagónico similar cuando fueron confrontados contra aislados de Pythium spp. y P. cactorum. Sin embargo, el PICR fue diferente para aislados de Pythium spp. (1.6-31.4%, p=0.05) y P. cactorum (8.1-92.1%, p=0.05). Se observó sobrecrecimiento (invasión total sobre los patógenos) de las cuatro especies de Trichoderma para todos los aislados de ambos Oomicetos después de ocho días post-confrontación (Cuadro 4). Gajera y Vakharia (2010) también observaron la más alta inhibición de crecimiento de los hongos fitopatógenos con algunas especies de Trichoderma. Gajera y Vakharia (2010) demostraron que dichas especies compiten contra hongos fitopatógenos y producen enzimas extracelulares (ej. quitinasa, β-1,3 glucanasa, y proteasas), además de ser antifúngicos, promotores del crecimiento, e inducen la resistencia de las plantas a patógenos. Entre las especies de Trichoderma probadas, T. asperellum mostró un bajo PICR (15.5-31.4%, p=0.05) en las tres especies de Pythium, con la inhibición más alta observada con P. irregulare al quinto día post-confrontación. En las confrontaciones con aislados de P. cactorum el PICR fue >84% (p=0.05) al octavo día post-confrontación (Cuadro 4; Figura 1a). De manera similar, Rios-Velasco et al. (2016) también observaron inhibiciones del crecimiento radial significativas en confrontaciones de T. asperellum contra Aspergillus nidulans y Penicillium crustosum, mostrando el potencial de estas cepas para ser utilizadas como agentes de control biológico de diferentes tipos de fitopatógenos. En nuestro estudio, T. harzianum mostró la misma tendencia en PICR al ser comparada con T. asperellum; cuando fue confrontada contra Pythium spp., la incidencia fluctuó entre 1.6 y 7.9% (p=0.05) pero en confrontaciones con aislados de P. cactorum, la incidencia osciló de 86.6 a 92.1% (p=0.05) (Cuadro 4; Figura 1b). Esta diferencia se puede atribuir a la habilidad de T. harzianum para producir el antibiótico 6-pentil-α-pirona y regular los genes involucrados en la biosíntesis de tricotecenos y micotoxinas con actividades antimicrobianas de amplio espectro (Cooney et al., 2001). Adicionalmente, este antagonista llenó completamente la caja de Petri después de tres días.
Trichoderma gamsii mostró valores de PICR de 15.5 a 29.7% (p=0.05) en aislados de Pythium spp. y >81% en aislados de P. cactorum. Estos resultados fueron similares a los obtenidos con T. asperellum (Cuadro 4). Por otra parte, T. atroviride mostró la misma tendencia con valores de PICR de 11.8 a 26.3% (p=0.05) en aislados de Pythium spp. y >89% (p=0.05) en aislados de P. cactorum al quinto y sexto día post-confrontación, respectivamente (Cuadro 4; Figura 1c). En este estudio, T. atroviride fue el mejor agente de control para la inhibición del crecimiento de P. cactorum en medio PDA. Gajera y Vakharia (2010) también observaron que T. atroviride fue efectivo en reducir la incidencia de pudrición de cuello, reduciendo la virulencia del patógeno. Roiger y Jeffers (1991) sugirieron que las especies de Trichoderma son promisorios agentes de control biológico a P. cactorum, porque su uso es compatible con prácticas de manejo y otras enfermedades de raíz, incluyendo el uso de plaguicidas. Las especies de Trichoderma también mostraron actividades de promoción de crecimiento vegetal, sugiriendo otro beneficio potencial para su uso. De acuerdo con la escala de Bell, se mostró un antagonismo Tipo 1 para las cuatro especies de Trichoderma, donde el antagonista (Trichoderma) sobrecreció completamente la colonia del patógeno y cubrió completamente el medio, después de seis días post-inoculación contra aislados de Phytophthora.
Cuadro 4 Inhibición de crecimiento de Oomicetos, cuando se confrontaron in vitro contra especies de Trichoderma a los 8 d post-inoculación y por especies de Bacillus al sexto día post-inoculación.
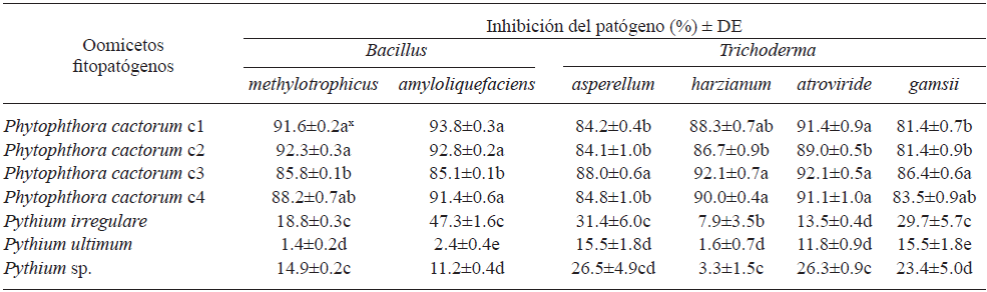
x Las letras en la misma columna indican diferencias significativas en los tratamientos donde las medias seguidas por la misma letra no son diferentes significativamente (ANVA) de acuerdo con la prueba de Tukey (p=0.05)
Actividad antagonista in vitro de Bacillus spp. vs Oomicetos
Las bacterias B. amyloliquefaciens y B. methylotrophicus inhibieron el crecimiento de aislados de P. cactorum en más del 85% (p=0.05) al ser evaluadas in vitro, y ambos antagonistas mostraron valores de PICR menores al 50 y 20% (p=0.05), respectivamente, cuando fueron confrontados contra Pythium spp. Bacillus amyloliquefaciens mostró un PICR fluctuando de 85.1 a 93.8% cuando fue confrontado contra P. cactorum y de 2.4 a 47.6% (p=0.05) contra Pythium spp. al sexto día post-confrontación. Bacillus methylotrophicus mostró la misma tendencia, con un PICR de 1.4 a 18.8% (p=0.05) cuando fue confrontado contra Pythium spp. y un valor de PICR superior al 85% (p=0.05) en aislados de P. cactorum (Cuadro 4; Figura 1d-f). Ambas bacterias mostraron altos PICR en confrontaciones con aislados de P. cactorum, reflejando probablemente la producción de antibióticos, enzimas que atacan los componentes de las células de patógenos o compuestos volátiles que inhiben el crecimiento del patógeno. Sin embargo, se observó un comportamiento diferente cuando fueron confrontados con especies de Pythium exhibiendo rápido crecimiento, y los rangos de PICR fueron bajos comparados con los aislados de P. cactorum cuando fueron confrontados in vitro contra Bacillus spp. (Cuadro 4). Jeyaseelan et al. (2012) evaluaron el efecto antagonista de algunas especies de Bacillus contra Pythium aphanidermatum y también observaron un bajo efecto de inhibición por algunas especies antagonistas como las evaluadas en el presente estudio, reflejando la incapacidad de las especies de Bacillus para producir compuestos antimicrobianos altamente efectivos contra algunas especies de Pythium. De acuerdo con Guillén-Cruz et al. (2006), el género Bacillus mostró efectos antagonistas in vitro contra Phytophthora spp., siendo B. amyloliquefaciens el más eficiente y mostrando un efecto de promoción de crecimiento vegetal, probablemente contribuyendo a la fijación de los macronutrientes y la solubilización de fosfatos y otros micronutrientes.
Notablemente, las especies de Bacillus mostraron un halo de inhibición >29 mm cuando fueron confrontadas contra P. cactorum a los 6 d post-confrontación y no hubo día de contacto (Cuadro 5). Algunos de los mecanismos más importantes de las bacterias antagonistas como Bacillus son la antibiosis, consistiendo en la producción de compuestos antibióticos y la inhibición de otros microbios (Intana et al., 2008). Souto et al. (2004) demostraron que B. amyloliquefaciens ejerce propiedades antifúngicas y que puede ser considerado como un buen agente de control biológico. Dev-Sharma et al. (2013) obtuvieron >90% de inhibición en hongos fitopatógenos cuando fueron confrontados in vitro contra B. methylotrophicus. Silo-Suh et al. (1994) reportaron que algunas especies de Bacillus tienen la habilidad de producir efectivos antibióticos de amplio espectro, tales como péptidos, lipopéptidos, aminoglicósidos y aminopolióles. Por otra parte, Wheatley (2002) se refirió a los compuestos orgánicos volátiles producidos por diferentes bacterias y hongos como ideales en las interacciones microbianas, reflejando su eficacia sobre un amplio rango de escalas espaciales, inhibiendo el crecimiento de muchos hongos patógenos, ya que estos compuestos tienen efectos directos o indirectos en la actividad de enzimas fúngicas específicas.
Cuadro 5 Halo de inhibición mostrado cuando se confrontaron in vitro especies de Bacillus contra Oomicetos a los seis días después de la inoculación.

x Las letras en la misma columna indicaron diferencias significativas en los tratamientos donde las medias seguidas por la misma letra no son diferentes significativamente (ANVA) de acuerdo con la prueba de Tukey (p=0.05)
CONCLUSIONES
Los hongos y Oomicetos fueron los más abundantes en los aislados recolectados, siendo Fusarium oxysporum la especie más representativa. Los portainjertos de 1 año de edad G.935, Estándar y M.25 fueron los más resistentes a los aislados probados, lo cual se puede atribuir a la edad y genotipo de los portainjertos. Nuestros resultados demostraron que ambos grupos antagonistas pueden ser considerados como candidatos para ser usados como agentes de control biológico para P. cactorum, el cual es un importante agente causal de enfermedades de raíz en portainjertos de manzano. Se requieren estudios adicionales in vitro e in vivo para confirmar la efectividad de aislados de Trichoderma y Bacillus contra varios agentes causales de enfermedades de raíz en huertos de manzano en Chihuahua.

