











 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
En Colombia el banano es el tercer recurso agrícola de mayor exportación después del café y las flores, sin embargo estos niveles de exportación son afectados en gran medida por los problemas fitopatológicos (AUGURA, 2015). Entre los patógenos que afectan el banano, Mycosphaerella fijiensis Morelet, agente causal de la sigatoka negra es considerada la enfermedad más importante y con mayor relevancia a nivel mundial (FRAC, 2014). Y debido a la susceptibilidad de este cultivo a la enfermedad se requiere el uso de múltiples fungicidas a frecuencias relativamente altas con costos estimados en más del 30 % del total de la producción (Churchill, 2011).
En Colombia así como en los demás países exportadores de banano donde se ha reportado a M. fijiensis, su control se ha vuelto cada vez más costoso tanto económica como ambientalmente debido a que el número de tratamientos químicos necesarios ha aumentado con el tiempo a causa de la rápida aparición de resistencia a los diferentes fungicidas que han sido empleados (Pérez-Vicente, 2006; de Bellaire et al., 2010; FRAC, 2014).
A pesar que la resistencia a fungicidas es uno de los principales limitantes en el control de esta enfermedad se debe tener en cuenta que la resistencia es una causa directa de la misma aplicación de los fungicidas y por lo tanto inevitable (Grimmer et al., 2015), sin embargo su intensidad e impacto sobre los costos finales de producción sí pueden ser controlados y para este fin se ha considerado que la herramienta más útil es implementar un monitoreo continuo de la evolución de la resistencia, que permita la identificación de los individuos resistentes a la acción de un fungicida, los que según su número determinarán si la resistencia se está afianzando dentro de la población (Van Den Bosch et al., 2011).
En M. fijiensis el método de monitoreo más confiable para identificar la presencia de individuos resistentes consiste en la medición de la longitud del tubo germinativo de esporas que germinan en presencia del fungicida (FRAC, 2008). No obstante en una población de M. fijiensis se pueden encontrar diferentes niveles de inhibición de los tubos germinativos a una concentración del fungicida, por lo que algunos individuos resistentes pueden pasar desapercibidos (Pérez-Vicente, 2006).
Con esta investigación se busco complementar los métodos actuales para monitorear la resistencia a los fungicidas piraclostrobin, epoxiconazol y tridemorf, al aumentar las variables fenotípicas que pueden ser relacionadas con el nivel de sensibilidad, lo que daría mayor certeza al identificar los individuos resistentes y de esta forma poder definir con anterioridad a la aplicación de uno de estos ingredientes activos si la población patógena se encuentra conformada por un mayor o menor número de individuos tolerantes a estos ingredientes activos y así evitar una mayor presión de selección sobre estos individuos.
Materiales y Métodos
Diseño experimental
El diseño experimental se realizó a partir de las siguientes consideraciones:
1) Cada finca se tomó como una población patógena independiente cuyos individuos estuvieron representados por las múltiples esporas o individuos producidos en el ciclo infeccioso de la enfermedad. 2) Si se considera al fenotipo como la expresión del genotipo en función de un determinado ambiente y que en el ambiente de M. fijiensis la presión ejercida por los fungicidas favorece los genotipos que permiten tolerar en mayor medida la acción de estos fungicidas, se esperaría por lo tanto que si hay un genotipo favorecido este se expresaría a través de rasgos fenotípicos identificables entre individuos. 3) Por evaluaciones previas se ha observado que diferentes rasgos fenotípicos se repiten entre individuos, indicando que probablemente exista una relación entre estos rasgos. 4) Los ingredientes activos evaluados han sido aplicados comercialmente en campo, por lo que la población y los individuos patógenos de este estudio ya han sido sometidos a la presión de selección de estos fungicidas. En base a estas consideraciones se diseñaron experimentos in vitro dirigidos a comparar si la sensibilidad poblacional (finca) o la sensibilidad individual (monospórico) hacia piraclostrobin, epoxiconazol y tridemorf, tiene relación con el comportamiento fenotípico de cada individuo.
Variables fenotípicas evaluadas
Se realizó una caracterización fenotípica que permitió dividir los aislamientos en dos grupos, uno conformado por dos variables cuantitativas y el otro conformado por seis variables cualitativas (Figura 1). El primer grupo estuvo conformado por los rasgos fenotípicos calificados cuantitativamente: Longitud del tubo germinativo y Producción de biomasa (peso del micelio); el segundo grupo estuvo constituido por variables cualitativas: 1) Número de tubos germinativos (Un solo tubo germinativo, dos tubos germinativos de similar longitud o dos tubos germinativos desiguales en el cual uno mide menos de la mitad del otro. 2) Ramificación del tubo germinativo (Tubos germinativos con ramificaciones o sin ramificaciones, 3) Color del micelio (Color blanco, rosa o gris), 4) Morfología del micelio (división circular sobre el micelio, división paralela sobre el micelio, división de forma irregular y división de forma indefinida, 5) Presencia de exudado (exudado presente o ausente, 6) Forma del micelio (ovoide o indeterminada "amorfa").
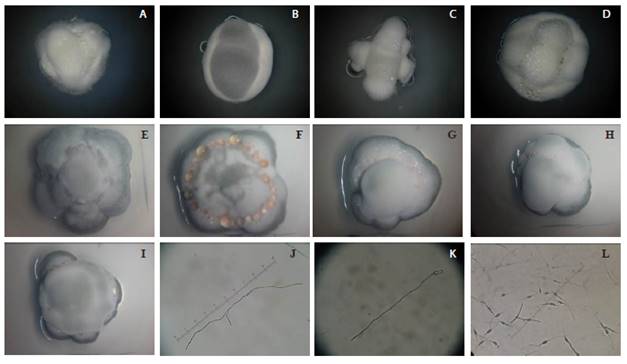
Figura 1 Variables fenotípicas cualitativas. De izquierda a derecha: A) Color blanco, morfología indefinida, con exudado, forma amorfa. B) color gris, morfología paralela, sin exudado, forma ovoide. C) color blanco, morfología indefinida, sin exudado, forma amorfa. D) color blanco, morfología paralela, con exudado, forma ovoide. E) color gris, morfología circular, sin exudado, forma ovoide. F) color rosa, morfología irregular, con exudado, forma ovoide. G) color rosa, morfología circular, con exudado, forma ovoide. H) color rosa, morfología paralela, con exudado, forma ovoide. I) color gris, morfología indefinida, con exudado, forma ovoide. J) dos tubos germinativos de similar longitud, con ramificaciones. K) un solo tubo germinativo, sin ramificaciones. L) dos tubos germinativos de similar longitud y en el que uno mide menos de la mitad del otro, sin ramificaciones.
Material vegetal
Se emplearon muestras vegetales con síntomas en estadio seis de la enfermedad (estadio final de la enfermedad caracterizado por la recombinación sexual y la producción de esporas), obtenidas durante los años 2013 y 2014, desde 12 fincas bananeras de Colombia, siete ubicadas en la región de Urabá y seis en la región de Santa Marta. De las 12 fincas, 10 fueron fincas comerciales y por lo tanto la población patógena allí ubicada ha sido constantemente sometida a la presión de los fungicidas y se espera que en mayor o menor medida hayan generado pérdida de sensibilidad, mientras las dos fincas restantes (nombradas como finca silvestre) no son fincas comerciales, por lo que la población patógena que allí se encuentra no ha sido sometida a la presión de los fungicidas y presenta altos niveles de sensibilidad hacia los diferentes ingredientes activos.
Determinación de la sensibilidad poblacional de M. fijiensis a los fungicidas
Inicialmente se cuantificó la sensibilidad en cada finca a partir del protocolo avalado por FRAC (Fungicide Resistance Action Commitee) (FRAC, 2008) para la evaluación de fungicidas sistémicos en M. fijiensis, denominado como "Ascospore germ tube elongation" basado en la medición de la longitud del tubo germinativo que permite valorar el nivel de sensibilidad poblacional a partir del promedio de mediciones individuales. Las concentraciones así como el número de individuos o esporas medidas se fijaron según las especificaciones FRAC. Para fungicidas QoI al que pertenece piraclostrobin se evaluaron por finca tres concentraciones (0-1-10 mg/L) y se midió un número de 600 esporas por finca. Para los dos ingredientes activos restantes se tomaron las especificaciones dadas para fungicidas tipo SBI's (Sterol Biosynthesis Inhibitors) al que pertenecen tanto epoxiconazol como tridemorf. De esta forma para el epoxiconazol se evaluaron por finca 210 esporas en cinco concentraciones (0-0.01-0.03-0.1 y 1 mg/L). Mientras para el tridemorf se evaluaron seis concentraciones (0-0.1-0.3-1-3 y 10 mg/L) y un número de 240 esporas por finca.
Aislamientos monospóricos
Luego de establecer la sensibilidad poblacional y a partir de este mismo montaje se seleccionaron al azar esporas germinadas en ausencia del fungicida (0 mg/L), a las que previamente se les cuantificó la longitud, la existencia de ramificaciones y el número de tubos germinativos. Estas esporas fueron aisladas por medio de un microscopio de luz y un capilar de vidrio y llevadas a medio nutritivo PDA donde crecieron a temperatura ambiente (26±1 °C) por un periodo de 40 días hasta obtener un cultivo monospórico. Transcurrido este tiempo se determinó el color, la morfología, la presencia de exudado y la forma del micelio, luego y aprovechando el crecimiento compacto no extendido de M. fijiensis bajo condiciones in vitro, el total de micelio fue pesado en balanza analítica y se pudo cuantificar la biomasa producida. Se obtuvo un total de 123 cultivos monospóricos sobre los que se realizaron las comparaciones.
Determinación de la sensibilidad individual de M. fijiensis a los fungicidas
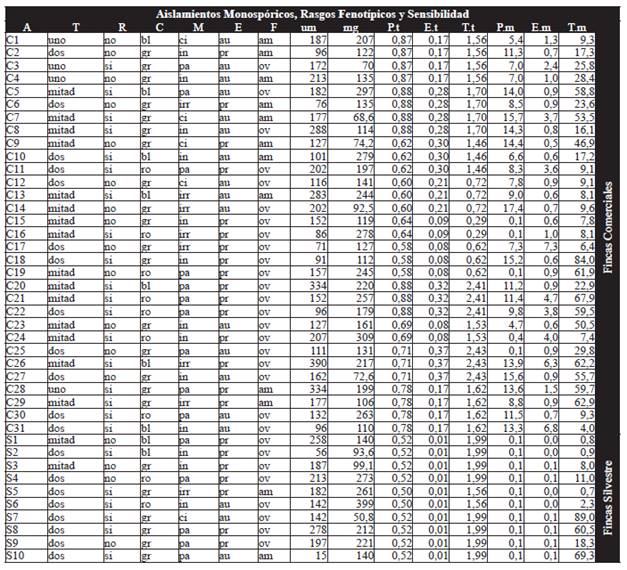
A partir de los 123 cultivos monospóricos, se seleccionaron los de mayores diferencias en la longitud del tubo germinativo y en el peso del micelio (debido a que sobre estos rasgos se basó la cuantificación de sensibilidad) y se obtuvo un total de 41 aislamientos monospóricos a los que se les cuantificó la sensibilidad individual a los tres ingredientes activos empleando microtécnica espectrofotométrica (Cuadro 1). Esta técnica es también avalada por FRAC pero no para M. fijiensis, debido a que la ausencia de esporulación sexual de este patógeno bajo condiciones in vitro implica que esta evaluación deba realizarse sobre cultivos monospóricos o sobre un limitado número de individuos que no representarían la población, de esta forma esta técnica no sería útil para la determinación de la sensibilidad de una población pero si para determinar la sensibilidad de un individuo de esa población. Esta técnica se basó en preparar a partir del micelio de cada uno de los 41 aislamientos monospóricos, una suspensión a una concentración de 104 fragmentos de micelio/ml, que fue puesta en incubación durante ocho días en platos ELISA de 96 pozos. Para cada ingrediente activo se evaluaron cuatro concentraciones: piraclostrobin (0-0.1-1-20 mg/L); epoxiconazol (0-0.1-1 y 10 mg/L) y tridemorf (0-1-10 y 100 mg/L). Estas concentraciones se fijaron a partir de evaluaciones previas en las que se estableció el rango de inhibición para cada uno de los ingredientes activos cuando se empleó esta técnica.
Cuadro 1 Rasgos fenotípicos para los 41 aislamientos monospóricos (31 provenientes de fincas comerciales y 10 provenientes de fincas silvestres) a los que se les midió la sensibilidad tanto por medición del tubo germinativo como por microtécnica. (A) Aislamiento. (T) Número de tubos germinativos. (R) Ramificación. (C) Color (bl: blanco. ro: rosa. gr: gris). (M) Morfología (ci: circular. pa: paralela. in: indefinida. irr: irregular). (E) Exudado (au: ausente. pr: presente). (F) Forma (am: amorfa. ov: ovoide). (um) Longitud tubo germinativo. (mg) Peso biomasa. (P.t) IC50 piraclostrobin tubo germinativo. (E.t) IC50 epoxiconazol tubo germinativo. (T.t) IC50 tridemorf tubo germinativo. (P.m) IC50 piraclostrobin microtécnica. (E.m) IC50 epoxiconazol microtécnica. (T.m) IC50 tridemorf microtécnica.
Cuantificación de la sensibilidad
El parámetro empleado para cuantificar la sensibilidad tanto a nivel poblacional como individual fue la IC50 "half maximal inhibitory concentration", calculada a partir de la regresión lineal de dos puntos (Y = mX + b), donde los puntos graficados fueron las concentraciones por encima y por debajo entre las que se encontraría el 50 % de inhibición y la curva resultante para cada fungicida se representó en un gráfico de Excel junto con la IC50 estimada. La IC50 estimada fue la dosis o concentración (eje X) que produjo el 50 % de inhibición (eje Y) en la curva (Chinn, 1997).
Análisis estadístico
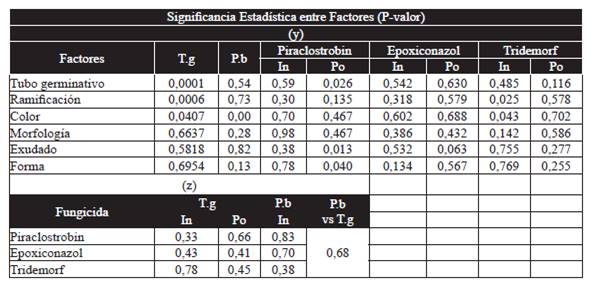
Para establecer si existieron diferencias significativas entre la expresión de los diferentes rasgos fenotípicos, además entre la expresión de estos rasgos con la sensibilidad individual o poblacional a los fungicidas evaluados, se realizaron análisis de varianza con un 95 % de confianza para comparar entre los factores que representan las variables cuantitativas: a) Longitud del tubo germinativo, b) Producción de biomasa, c) Sensibilidad poblacional o individual a cada ingrediente activo (IC50). Y los factores que representan las variables cualitativas: a) Número de tubos germinativos, b) Ramificación del tubo germinativo, c) Coloración del micelio, d) Morfología del micelio, e) Presencia de exudado, f) Forma del micelio. Aceptando la significancia estadística siempre que el P-valor fuera menor que 0.05 (Cuadro 2). Se realizaron además pruebas de comparaciones múltiples dentro de los factores empleando el método de la mínima diferencia significativa de Fisher (LSD) para definir si existieron diferencias significativas entre las medias obtenidas para cada variable cuantitativa cuando se comparó con la presencia de cada rasgo que representa las variables cualitativas (Cuadro 3).
Cuadro 2 P-valor obtenidos a partir de análisis de varianza que comparó entre variables las cuantitativas (T.g: longitud tubo germinativo; P.b: producción biomasa; In: sensibilidad individual hacia el fungicida; Po: sensibilidad poblacional hacia el fungicida) y las variables cualitativas (y). Y cuando se comparó solo entre las variables cuantitativas (z). Desde que el P-valor fuera inferior a 0,05, existió significancia estadística.
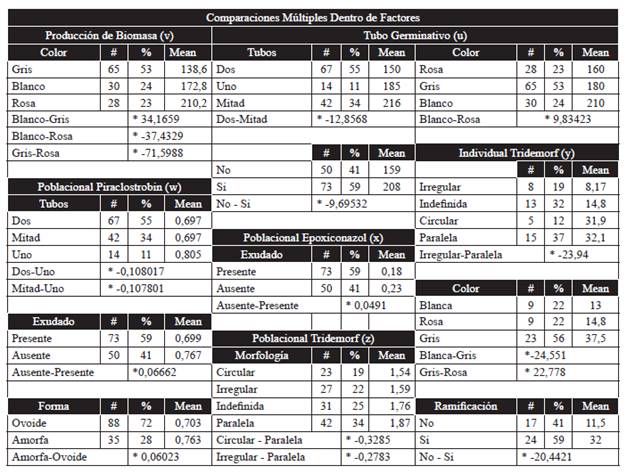
Cuadro 3 Se muestra los rasgos evaluados, el número de individuos con ese rasgo, su porcentaje poblacional, la media estadística de la longitud del tubo germinativo en micrómetros (u), la media estadística del peso de la biomasa en miligramos (v), la media estadística de las IC50 para los rasgos fenotípicos asociados a la sensibilidad poblacional a piraclostrobin (w), epoxiconazol (x), tridemorf (z), y la sensibilidad individual al tridemorf (y). (*) Indica diferencia significativa al comparar factores.
Ingredientes activos
Los ingredientes activos evaluados, piraclostrobin; epoxiconazol y tridemorf, fueron tomados desde productos pertenecientes a la empresa BASF Química S.A. y empleados en la industria bananera bajo los nombres comerciales de COMET (Piraclostrobin 250 g/L); OPUS (Epoxiconazol 75 g/L) y CALIXIN (Tridemorf 750 g/L).
Resultados
Rasgos fenotípicos asociados a la longitud del tubo germinativo
El análisis de varianza mostró diferencias entre factores para el número de tubos germinativos (Cuadro 2y). Es así como dentro de los factores al comparar las medias se observó que las esporas que generaron dos tubos germinativos, pero en el que uno de ellos alcanzó menos de la mitad del otro, presentaron una relación significativa con tubos germinativos de mayor longitud (X=216 um). Sin embargo cuando se presentaron también dos tubos pero ambos tuvieron tamaños similares se relacionaron con una menor longitud (X=150 um). Esporas que solo generaron un tubo germinativo se relacionaron a longitudes intermedias y no presentaron diferencias significativas con aquellas esporas de dos tubos germinativos (X=185 um). Los individuos que presentaron dos tubos germinativos de tamaño similar y por lo tanto la menor longitud fueron mayoría dentro de la población y representaron el 55 % de esta, mientras los de mayor longitud representaron un 34 % de la población (Cuadro 3u). La existencia de ramificaciones fue otro factor que presentó diferencias significativas entre factores en relación a la longitud del tubo germinativo (Cuadro 2y). Dentro de los factores y al comparar las medias la aparición de estas ramificaciones estuvo significativamente relacionada a tubos germinativos de mayor longitud (X=208 um) que a de menor longitud (X=159 um). Los individuos que presentaron ramificaciones y una mayor elongación fueron mayoría dentro de la población y representaron el 59 % (Cuadro 3u). La longitud del tubo germinativo también presentó diferencias significativas entre los factores con el color del micelio (Cuadro 2y), dentro de los factores y al comparar las medias se observó que una mayor longitud dio lugar a micelio color blanco (X=210 um), mientras una menor longitud generó micelio color rosa (X=160 um). No existieron diferencias significativas cuando se obtuvo micelio color gris (X=180 um). El micelio de color gris relacionado con esporas de tubo germinativo de tamaño intermedio fue mayoría dentro de la población y representó el 53 %. Mientras el micelio de color blanco relacionado a una mayor longitud representó un 24 % de la población, porcentaje similar al 23 % del micelio color rosa relacionado a una menor longitud (Cuadro 3u).
Rasgos fenotípicos asociados a la producción de biomasa
Existieron diferencias significativas entre los factores para la cantidad de biomasa producida por cada individuo y el color del micelio (Cuadro 2y). Dentro de los factores y al comparar las medias los individuos de menor peso correspondieron a los de color gris (X=139 mg), los de color blanco se relacionaron a un peso mayor que el gris (X=173 mg) pero menor que el rosa (X=210 mg), que fue el color asociado a una mayor producción de biomasa. La distribución proporcional indicó que un 53 % de los individuos presentaron micelio color gris y por lo tanto un menor peso, mientras un 24 % presentó micelio color blanco y el 23 % restante presentó micelio color rosa y un mayor peso (Cuadro 3v).
Rasgos fenotípicos asociados a la sensibilidad a fungicidas
Se estableció el nivel de sensibilidad hacia los fungicidas evaluados a través de una IC50 hallada por microtécnica espectrofotométrica desde cultivos monospóricos que permitieron conocer una respuesta individual y a través de una IC50 hallada a partir del promedio de las longitudes alcanzadas por los tubos germinativos de un número determinado de esporas que permitieron conocer una respuesta poblacional.
Piraclostrobin
Respuesta individual. Entre los factores no se encontraron diferencias significativas entre la sensibilidad de un individuo hacia piraclostrobin y las características fenotípicas evaluadas (Cuadro 2y).
Respuesta poblacional: Entre los factores existieron diferencias significativas para el nivel de sensibilidad poblacional hacia piraclostrobin y el número de tubos germinativos (Cuadro 2y). Dentro de los factores y al comparar las medias se encontró que una espora con un solo tubo germinativo estuvo significativamente más relacionada a una población con una menor sensibilidad a piraclostrobin (IC50=0,805) que una espora con dos tubos germinativos, estas últimas relacionadas a una población de mayor sensibilidad sin importar si uno de los tubos mide menos de la mitad del otro (IC50=0,697). La distribución proporcional indicó que el 89 % de la población presentó dos tubos germinativos y por lo tanto una menor IC50 poblacional, mientras el 11 % restante presentó un solo tubo germinativo y una mayor IC50 poblacional (Cuadro 3w). También se halló entre los factores diferencias significativas con la producción de exudado (Cuadro 2y) y dentro de los factores se encontró que al comparar las medias las esporas aisladas de una población con una menor IC50 (IC50= 0,699) y por lo tanto una mayor sensibilidad a piraclostrobin estuvieron significativamente más relacionadas con la producción de exudado en su micelio, mientras que el aumento en las IC50 (IC50= 0,767) se relacionó con la ausencia en la producción de exudado. La distribución proporcional indicó que el 59 % de la población presentó exudado, mientras el 41 % restante careció de él (Cuadro 3w). Entre los factores la forma del micelio del cultivo monospórico, también exhibió diferencias significativas con la sensibilidad de la población a piraclostrobin (Cuadro 2y). Y dentro de los factores se encontró al comparar las medias que esporas provenientes de una población con una menor IC50 (IC50= 0,703) estuvieron significativamente relacionadas con una forma ovoide en su micelio, mientras que si la población presentaba una mayor IC50 (IC50= 0,763) el micelio resultante estuvo más relacionado a no tener una forma determinada (amorfa). La distribución proporcional indicó que el 72 % de los individuos presentaron forma ovoide y por lo tanto estuvieron relacionados con una mayor sensibilidad a piraclostrobin, mientras el 28 % restante no presentó una forma determinada (Cuadro 3w).
Epoxiconazol
Comportamiento individual. Entre los factores no se encontraron diferencias significativas entre las características fenotípicas evaluadas y la sensibilidad de los individuos monospóricos hacia epoxiconazol (Cuadro 2y).
Comportamiento poblacional. Entre los factores no se encontraron diferencias significativas (Cuadro 2y), sin embargo dentro de los factores y al comparar entre medias se observó que la sensibilidad poblacional a epoxiconazol presentó diferencias significativas para el rasgo producción de exudado, debido a que aislamientos provenientes de una población más sensible a este ingrediente activo (IC50=0,18) presentaron exudado mientras aislamientos menos sensibles no lo presentaron (IC50=0,23). La distribución proporcional indicó que el 59 % de la población presentó exudado y por lo tanto una mayor sensibilidad al epoxiconazol, mientras el 41 % restante no presentó exudado (Cuadro 3x).
Tridemorf
Comportamiento individual. Entre los factores se encontraron diferencias significativas para la existencia de ramificaciones en el tubo(s) germinativo (Cuadro 2y). Y dentro de los factores al comparar las medias se observó que la presencia de ramificaciones estuvo significativamente relacionada a una menor sensibilidad a tridemorf (IC50=32), mientras la ausencia de ramificaciones se relacionó con una mayor sensibilidad (IC50=11,5). Al analizar la distribución proporcional se encontró que los individuos sin ramificaciones fueron minoría dentro de la población (41 %), mientras los individuos con ramificaciones y una menor sensibilidad fueron mayoría (59 %) (Cuadro 3y). Entre los factores también se observaron diferencias significativas para el color del micelio (Cuadro 2y) y dentro de los factores al comparar las medias se observó que una menor sensibilidad del cultivo monospórico a tridemorf (IC50=37,5) estuvo significativamente relacionada con un micelio color gris, mientras si el cultivo presentaba un color blanco (IC50=13) o rosa (IC50=14,8) se relacionó con una sensibilidad mayor. Cuando se evaluó la distribución proporcional se encontró que los aislamientos de color gris asociados con una menor sensibilidad a tridemorf fueron mayoría en la población y representaron el 56 %, mientras los aislamientos de color blanco y rosa se encontraron a una proporción igual del 22 % (Cuadro 3y). Dentro de los factores al comparar las medias para el del rasgo morfología, se observó que si un cultivo monospórico presentaba un micelio de morfología irregular este individuo estaría significativamente más relacionado con una sensibilidad mayor (IC50=8,17), ya que requeriría bajo condiciones in vitro de una menor concentración de tridemorf para lograr el mismo nivel de inhibición que un cultivo monospórico de morfología indefinida (IC50=14,8), circular (IC50=31,9) y paralela (IC50=32,1), esta última la de mayor IC50. La distribución proporcional mostró que la morfología paralela asociada a una menor sensibilidad a tridemorf estaba en un mayor porcentaje de individuos (37 %), seguida de la morfología indefinida (32 %) asociada a una sensibilidad mayor pero intermedia. La morfología irregular relacionada con la mayor sensibilidad a tridemorf se encontró a una menor proporción (19 %), mientras un menor porcentaje de individuos (12 %) presentaron morfología circular, que se caracterizó por tener los segundos niveles de sensibilidad más bajos, muy cercanos a los observados para la morfología paralela (Cuadro 3y).
Comportamiento poblacional. Entre los factores no se observaron diferencias significativas (Cuadro 2y), sin embargo dentro los factores al comparar las medias se observaron diferencias significativas para la morfología del micelio. Una población con mayor sensibilidad a tridemorf se relacionó con individuos de una morfología circular (IC50=1,54) o irregular (IC50=1,59), mientras una sensibilidad menor al tridemorf se relacionó con individuos de una morfología indefinida (IC50=1,76) o paralela (IC50=1,87). En este caso la distribución proporcional mostró que la morfología paralela asociada con una menor sensibilidad se encontró en una mayor proporción dentro de la población (34 %), seguida de la morfología indefinida (25 %), la irregular (22 %) y la circular (19 %) (Cuadro 3z).
Longitud del tubo germinativo y producción de biomasa en relación a la sensibilidad a fungicidas
La longitud alcanzada por el tubo germinativo y la cantidad de biomasa producida bajo condiciones in vitro y en ausencia del fungicida no presentaron diferencias significativas con el grado de sensibilidad adquirido o inherente que tuviera un individuo o la población a la que este individuo pertenecía, hacia piraclostrobin, epoxiconazol o tridemorf (Cuadro 2z). Es decir una mayor o menor IC50 no tuvo relación con una mayor o menor longitud del tubo o producción de biomasa (Cuadro 1).
Longitud del tubo germinativo en relación a la producción de biomasa
Cuando en ausencia del fungicida se comparó la longitud del tubo germinativo y la cantidad de biomasa, no se encontraron diferencias significativas entre estos dos factores (Cuadro 2z), es decir una mayor o menor elongación del tubo germinativo no tuvo relación con producir más o menos micelio (Cuadro 1).
Discusión
Cuando los resultados fueron analizados en relación a que rasgos fenotípicos predominaron en las poblaciones de M. fijiensis en Colombia, se encontró que el color del micelio fue un rasgo que a pesar de aparentemente no estar relacionado con el fitness si estuvo relacionado con rasgos representativos como la longitud del tubo germinativo y la producción de biomasa. El color gris fue el rasgo que estuvo presente en una mayor cantidad de individuos es decir que este sería un rasgo favorecido por la selección, sin embargo los individuos de mayor producción de biomasa o mayor tubo germinativo no presentaron este color, es decir los individuos de color gris habrían aumentado su número por encima de individuos de mejor fitness. Por lo tanto existirían presiones externas que estarían seleccionando los individuos de menor fitness y estas presiones estarían relacionadas a la acción de los fungicidas.
La selección ejercida por los fungicidas sería diferencial según el ingrediente activo y para el piraclostrobin estaría provocando que los individuos con rasgos asociados a una mayor sensibilidad fueran mayoría (Cuadro 3w), indicando que los individuos con mayores oportunidades de propagación serían sensibles a piraclostrobin. Alteraciones en la eficacia biológica asociadas a la resistencia a fungicidas QoI´s ya han sido reportadas para otros hongos fitopatógenos (Veloukas et al., 2014; Fernández-Ortuño et al., 2010; Ma, B. y Tredway 2013). Un monitoreo en el tiempo de estos rasgos asociados a una mayor o menor sensibilidad ayudaría a establecer si el piraclostrobin afecta el fitness de M. fijiensis y de comprobarse indicaría que este fungicida es de bajo riesgo de generar resistencia y no de alto riesgo como se cataloga actualmente (FRAC, 2014).
Se observó que tanto para piraclostrobin como para epoxiconazol los individuos sensibles presentaron exudado mientras los menos sensibles no, indicando una posible relación entre los mecanismos que generan resistencia a estos dos ingredientes activos. La comprobación de estos resultados permitiría establecer si en M. fijiensis al igual que en otros hongos fitopatógenos existe resistencia múltiple "Multidrug" (MDR) entre fungicidas QoI y DMI (Villani y Cox 2014; Kretschmer et al., 2009). Además se estaría advirtiendo a cerca de la inconveniencia de un programa de fumigación basado en la mezcla de epoxiconazol y piraclostrobin debido a que se estaría aumentando la presión de selección sobre individuos tolerantes a ambos ingredientes activos.
Para tridemorf los rasgos asociados a una menor sensibilidad se encontraron en un mayor porcentaje de la población, sin embargo esto no significó que existiera pérdida de sensibilidad, debido a que individuos provenientes de poblaciones sometidas a la presión de fungicidas (fincas comerciales) no presentaron menor sensibilidad que aquellos provenientes de poblaciones no sometidas a la presión de fungicidas (fincas silvestre) (Cuadro 1). La explicación a estos resultados puede estar relacionada a lo observado con el rasgo morfología, donde los individuos asociados a una mayor sensibilidad sumaron un porcentaje poblacional similar al de las morfologías asociadas a una menor sensibilidad (Cuadro 3y; z). Estos resultados indicarían que dentro de una misma población existirían diferentes grupos de individuos con variables niveles de tolerancia, en el que ningún grupo tendría el dominio poblacional. Esta heterogeneidad de sensibilidades implicaría la no existencia de pérdida de sensibilidad pero también implicaría una presencia constante tanto de individuos sensibles como tolerantes.
Conclusión
Se identificaron individuos que expresaron rasgos fenotípicos relacionados con el nivel de tolerancia hacia un ingrediente activo, además este nivel de tolerancia determinaría si los individuos que poseen una serie de rasgos específicos son mayoría o no en la población. Por lo que una caracterización fenotípica a modo de monitoreo ayudaría a determinar que momento es el más propicio para la aplicación de un ingrediente activo según exista en ese momento un mayor o menor número de individuos con rasgos que expresen tolerancia hacia ese ingrediente activo.
Se encontró que algunos rasgos fenotípicos estaban relacionados o por el contrario se expresaban diferencialmente según el ingrediente activo, indicando que la caracterización fenotípica podría ayudar a determinar que mezclas de fungicidas aún pertenecientes a diferentes familias no conducirían a resistencia cruzada.

