











 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
El litchi (Litchi chinensis Soon) es un fruto subtropical (Coates et al., 1994) nativo del sur de China y sureste de Asia; su pericarpio es delgado y áspero, y cuando madura es de color rojo y con coarteaduras debido a la presencia de braquiesclereidas, cuya función podría estar asociada con la protección contra daños mecánicos o estrés fisiológico (Nacif et al., 2001). El arilo es blanco y cubre una semilla marrón obscuro.
Los principales problemas en la producción del litchi son rendimientos bajos o irregulares debido a caída excesiva de frutos (Mitra, et al., 2005); oxidación y coarteadura del pericarpio (Huang et al., 2001; Xu et al., 2005); daños por plagas causados por C. gloeosporioides Penz (Campbell y Campbell, 2001), Peronophythora litchii Chen Ex Ko, Tessaratoma papillosa Drury y Dasineura sp., y costos altos en cosecha y empaque (Xu et al., 2005).
La producción de litchi en México es de 19,888.05 toneladas, en un área de 3,738.48 hectáreas (SIAP, 2013). En Oaxaca, México, los cultivares Brewster y Mauritius se siembran comercialmente en la región del Papaloapan y los Bajos Mixes, respectivamente. En ambos cultivares, una mancha obscura regular en el pericarpio representa uno de los principales problemas en condiciones de campo y postcosecha. Los síntomas en campo se caracterizan como lesiones semicirculares cafés y secas; el arilo y semilla no muestran daño. No se han encontrado registros de su causa o importancia en México hasta ahora; sin embargo, productores de Oaxaca reportan daños superiores a 50% en producción. Por la importancia socioeconómica de este problema, el objetivo de este estudio fue determinar la etiología del manchado café en frutos de litchi en Oaxaca, México.
Materiales y Métodos
El experimento se estableció en huertos comerciales de litchi en tres municipios de Oaxaca, México durante los ciclos de producción 2008-2010. Los cultivares Brewster y Mauritius se evaluaron en San José Chiltepec (17° 57' 57.92" N y 96° 10' 11.68" O), y en San Juan Cotzocón (17° 26' 07.74" N y 95° 19' 53.97" O) y Loma Bonita (17° 58' 53.1" N y 95° 50' 58.1" O), respectivamente.
Síntomas y caracterización de hongos. Los frutos sintomáticos (infección natural) y asintomáticos de los cultivares Brewster y Mauritius en madurez comercial, se colectaron en mayo de 2009 de una parcela comercial de litchi en San José Chiltepec y San Juan Cotzocón, respectivamente. Cien frutos de cada cultivar se colectaron de manera aleatoria a partir de 25 árboles, posteriormente se lavaron con jabón y agua en laboratorio, se secaron a temperatura ambiente y se seccionaron longitudinalmente. En el pericarpio, arilo, semilla y pedúnculo se evaluó la consistencia del síntoma, su forma y color, así como la presencia de halo clorótico y signos de microorganismos asociados.
Para la caracterización de hongos aislados, se cosecharon 123 frutos sintomáticos y 195 asintomáticos del cultivas Brewster en el ciclo 2008-2009, y 60 y 70 en 2009-2010. Además, se muestrearon 193 frutos sintomáticos y 72 asintomáticos del cultivar Mauritius en el ciclo 2008-2009, y 102 y 96 en 2009-2010.
Dos secciones de pericarpio y arilo de 1.5 cm2 se cortaron de cada fruto y se desinfestaron en hipoclorito de sodio por tres minutos, se enjuagaron tres veces en agua destilada por un minuto cada uno, y se secaron en papel estéril. Las porciones de tejido se incubaron en PDA (Bioxon(tm)) y en un medio semi-selectivo a Colletotrichum [19.5 g PDA, 7.5 g agar (Merck(tm)), 8 mg iprodiona, 20 mg triadimefón, 50 mg chloramphenicol (Merck(tm)), y 200 mg estreptomicina] (Davis, 2003) durante 5-7 días a 25º C bajo luz fluorescente. Los crecimientos de hongos se purificaron por punta de hifa y cultivos monoconidiales.
Pruebas de patogenicidad. Colletotrichum, Pestalotiopsis, Fusarium, y Lasiodiplodia se aislaron de todas las muestras de pericarpio sintomáticas de los cultivares Brewster y Mauritius y se patogenicidad fue evaluada en condiciones de laboratorio y campo.
Las pruebas de patogenicidad en laboratorio se avaluaron en frutos asintomáticos y en madurez comercial del cultivar Mauritius colectados en campo en mayo de 2010. Veinte frutos desinfestados fueron heridos con una aguja estéril y en cada herida se les colocó un disco de 0.5 cm de agar con Colletotrichum, Pestalotiopsis, Fusarium, o Lasiodiplodia (Johnson y Cooke, 2002). Los frutos de control (testigos) fueron inoculados con un disco de agar de 0.5 cm sin micelio. Todos los frutos inoculados se mantuvieron en condiciones de cámara húmeda a 28 °C ±2 por 7 días. Las observaciones se realizaron cada 24 h para detectar síntomas.
La patogenicidad en campo se evaluó en mayo y junio de 2010 en 20 frutos asintomáticos por árbol del cultivar Mauritius (5 frutos/punto cardinal). Dos etapas de desarrollo se evaluaron: frutos en el inicio de maduración, y en madurez comercial. Los frutos se desinfestaron con etanol 70 % y heridos con aguja estéril en la mitad del fruto y en la base del pedúnculo. Los frutos fueron inoculados individualmente por la noche (18:00 - 20:00 hora del centro) mediante deposición de 0.5 ml de una suspensión 1 x 106 conidios.ml-1 de Colletotrichum, Pestalotiopsis, Fusarium, Lasiodiplodia o agua destilada estéril (testigo) (Dinh et al., 2003). Los puntos de inoculación se cubrieron con algodón estéril húmedo por 24 h para crear condiciones de alta humedad para el crecimiento de los hongos. El desarrollo de los síntomas se registró por 15 días. Los reaislamientos de los frutos infectados en laboratorio y campo se realizaron para verificar los postulados de Koch.
Caracterización morfológica del patógeno. Cuatro de 256 aislamientos de Colletotrichum obtenidos de frutos sintomáticos fueron seleccionados con base a su crecimiento miceliar y producción de esporas, e identificados como M26, M28, M29, y M30. El género (Sutton, 1980; Hanlin, 1990; Barnett y Hunter, 2006) y especies (Sutton, 1980; Smith y Black, 1990; Photita et al., 2005) se determinó a través de colonias monoconidiales. La formación de apresorios se indujo en PDA depositado en un portaobjeto e inoculado individualmente con esporas del M26 y M28-M30 (Johnston y Jones, 1997). La forma y tamaño de los apresorios se caracterizó a partir de colonias de hongos de 7 días de edad crecidas en PDA.
Las tasas de crecimiento de M26 y M28-M30 se evaluaron por siete días en placas de PDA incubadas a 28 °C. Los datos se analizaron estadísticamente por ANOVA (α=0.05) y agrupadas por pruebas de Tukey (SAS, 1997).
Identificación molecular. M26 y M28-M30 fueron aislados mediante el método AP (Sambrook y Russel, 2001). La integridad del ADN se verificó por electroforesis en gel de agar 1 % a 80 volts por 40 min en Buffer TBE, y teñido con bromuro de etidio.
Las regiones ITS1 e ITS2 se amplificaron por PCR con los iniciadores ITS4 (5′-TCCTCCGCTTATTGATATGC-3′) e ITS5 (5′-GGAAGTAAAAGTCGTAACAAGG-3′) (White et al., 1990), los cuales amplificaron un fragmento de 580 pb. Para identificar y confirmar especies, el ADN se amplificó con ITS4 y el iniciador específico CgInt (5′-GGCCTCCCGCCTCCGGGCGG-3′) (Mills et al., 1992), los cuales amplificaron un fragmento de 450 pb.
La amplificación por PCR se realizó en un termociclador (Applied BiosystemsTM 2720) en una reacción de 25 µl con 1.0 µl de cada iniciador (10 nM), 1.0 µl de dNTP (10 nM), 5 µl de Buffer PCR 10X, 2.5 µl de MgCl2 (50 nM), 0.5 µl de Taq Polimerasa, 0.5 µl de ADN y 13.5 µl de agua libre de nucleasas. Las reacciones PCR sin templado se utilizaron como control negativo.
La PCR consistió de una desnaturalización inicial de 5 min a 95 °C seguido de 35 ciclos de 1 min de desnaturalización a 95 °C, 1 min de alineamiento a 57 °C, y 1 min de extensión a 72 °C, con una extensión final de 10 min a 72 °C. Los productos PCR fueron electroforesados en un gel de agarosa 1 %, purificado con el kit QUIAGEN DNeasy Plant y secuenciado en un secuenciador automático 3730XL. Las secuencias fueron alineadas contra secuencias en el banco de genes y su homología se determinó en el Centro Nacional para Información Biológica (BLAST, 2011).
Resultados
Caracterización de síntomas. El exocarpo de Brewster mostró una lesión café obscura sin halo clorótico (Figura 1A). En el endocarpo, la lesión fue una mancha café obscura circular, hundida, semi-húmeda y sin halo clorótico (Figura 1B). En el exocarpo de Mauritius se desarrolló una lesión café obscura bien definida con halo clorótico (Figura 1D); y en el endocarpo se observó una mancha café obscura no húmeda, no hundida (Figura 1E). En ninguno de los frutos de Brewster y Mauritius se observó daños en el arilo (Figura 1C y 1F ), semilla, o pedúnculo, y no se observaron signos de microorganismos.

Figura 1 Frutos de litchi (Litchi chinensis Soon) con síntomas de antracnosis. Cultivar Brewster: A) Síntoma externo en pericarpio, B) Arilo asintomático, C) Síntoma interno de pericarpio. Cultivar Mauritius: D) Síntoma externo en pericarpio, E) Arilo asintomático F) Síntoma interno de pericarpio. Patogenicidad de C. gloeosporioides en frutos de litchi en laboratorio: G) Frutos de control, H) Síntoma externo de pericarpio, I) Síntoma interno de pericarpio. Patogenicidad en frutos de litchi Mauritius en campo: J) Frutos control, K) Síntoma externo de pericarpio, L) Síntoma interno de pericarpio. Bar = 1 cm.
Aislamiento de hongos. Colletotrichum sp., Pestalotiopsis sp., Fusarium sp., y Lasiodiplodia sp., se aislaron de pericarpios sintomáticos de Brewster y Mauritius en los ciclos de producción 2008-2009 y 2009-2010. Sin embargo, Colletotrichum sp. (256/640 aislamientos) y Pestalotiopsis sp. (71/640 aislamientos) fueron aislados más frecuentemente que los otros (Cuadro 1 y 2 ). Colletotrichum sp., Pestalotiopsis sp., y Lasiodiplodia sp., fueron también aislados de pericarpios asintomáticos de Brewster (Cuadro 1); y Fusarium de muestras asintomáticas de Mauritius (Cuadro 2). Colletotrichum sp., creció más en los fragmentos de tejido de pericarpio incubados en medio semi-selectivo que los incubados en PDA (Cuadro 1 y 2 ). Los aislamientos de Colletotrichum sp., produjeron conidios en medio semi-selectivo y PDA de 4 y 8-12 días de edad, respectivamente. Ningún microorganismo fue aislado de fragmentos de tejidos de arilo incubados en ambos medios de cultivo (Cuadro 1 y 2 ). En 13 de 15 fragmentos de pericarpio incubados en cámara húmeda existió producción de conidios al quinto día; las masas de conidios de color salmón se identificaron morfológicamente como Colletotrichum sp.
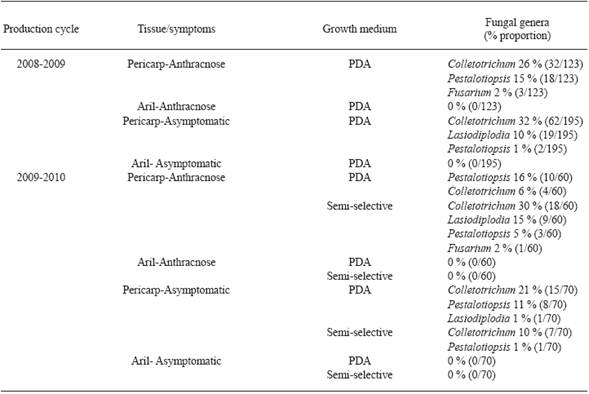
Cuadro 1 Géneros de hongos aislados de frutos de litchi sintomáticos y asintomáticos (cultivar Brewster), en San José Chiltepec, Oaxaca.
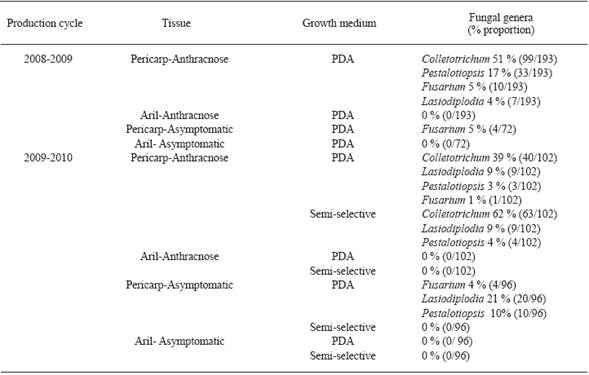
Cuadro 2 Géneros de hongos aislados de frutos de litchi sintomáticos y asintomáticos (cultivar Mauritius), en San Juan Cotzocón, Oaxaca.
Pruebas de patogenicidad. Los resultados en pruebas de laboratorio mostraron que 85 % (17 de 20) de los frutos inoculados con Colletotrichum sp., desarrollaron síntomas en el pericarpio en forma similar a los observados en campo. Estos síntomas aparecieron 72 h después de la inoculación como lesiones pequeñas circulares de color café brillante (Figura 1G). En el endocarpo se desarrolló una lesión café obscura circular y semi-húmeda (Figura 1H). Los frutos testigo (Figura 1I) y frutos inoculados con Fusarium sp., Pestalotiopsis sp., o Lasiodiplodia sp., no mostraron ningún síntoma.
En campo, los síntomas se desarrollaron en 95% (19 de 20) de los frutos de Mauritius en madurez comercial inoculados con Colletotrichum sp. en la parte media del pericarpio en mayo, y 90% en junio. Los síntomas se observaron ocho días después de la inoculación como lesiones café obscuras en el pericarpio (Figura 1J). Los síntomas en el endocarpo fueron lesiones café obscuras, circulares y hundidas (Figura 1K).
Los frutos testigo (Figura 1L) y frutos inoculados con Fusarium sp., Pestalotiopsis sp., o Lasiodiplodia sp., no mostraron ningún síntoma.
El aislamiento de Colletotrichum que indujo el síntoma de antracnosis en condiciones de laboratorio y campo fue reaislado e identificado como C. gloeosporioides (Sutton, 1980).
Caracterización e identificación del patógeno. La mayoría (193) de los 256 aislamientos de C. gloeosporioides formaron micelio blanco-gris claro, ligeramente algodonoso, con abundantes masas de conidios naranja brillante en círculos concéntricos (Figura 2A, 2D y 2G ). Algunos (78) de ellos produjeron un micelio blanco con pocas masas conidiales próximas al punto de transferencia (Figura 2J). Cuatro de estos 256 aislamientos fueron designados como M26, M28, M29 y M30.
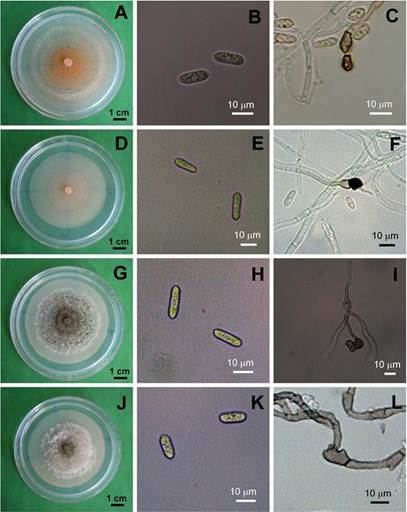
Figura 2 Morfología colonial (A, D, G, J), conidios (B, E H, K) y apresorios miceliales (C, F, I, L) de cuatro aislamientos representativos M26 (A, B, C), 28 (D, E, F), 29 (G, H, I), 30 (J, K, L) de Colletotrichum gloeosporioides de frutos de litchi, después de cinco días en PDA.
Los conidios de M26 y M28-M30 fueron cilíndricos (Cuadro 3 y Figura 2B, 2E , 2H y 2K ). Los conidios de M26 fueron estadísticamente más largos que los de M28, los cuales tuvieron los mayores diámetros (Cuadro 3). Los cuatro aislamientos formaron apresorios clavados (Figura 2C, 2F , 2I y 2L ). No existieron diferencias entre las tasas de crecimiento miceliar de los cuatro aislamientos incubados a 28 °C en PDA (α= 0.05) (Cuadro 3).
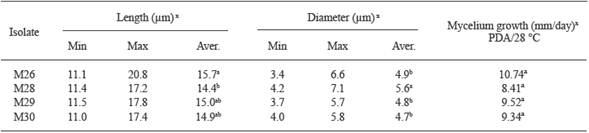
xValores con las misma letra en cada columna son estadísticamente iguales, Tukey (P≤0.05).
Cuadro 3 Tamaño de conidio y tasas de crecimiento miceliar de los aislamientos M26 y M28-M30 de C. gloeosporioides obtenidos de frutos de litchi con síntomas de antracnosis en San José Chiltepec y Loma Bonita, Oaxaca (ciclo de producción 2009-2010).
Identificación molecular. Los iniciadores ITS4-ITS5 e ITS4-CgInt amplificaron un fragmento de 580 y 450 pb, respectivamente, lo cual indicó que los aislamientos M26 y M28-M30 corresponden a C. gloeosporioides (Figura 3).
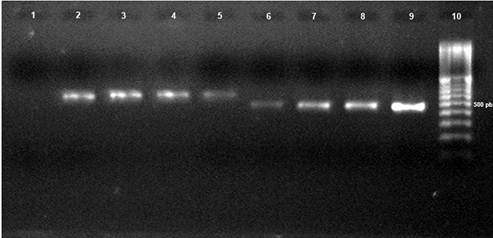
Figura 3 Producto de PCR de cuatro aislamientos característicos (M26, M28, M29, y M30) de Colletotrichum gloeosporioides. Lineas: 1) Control (H2O PCR); 2) M26- ITS4/ITS5; 3) M28- ITS4/ITS5; 4) M29- ITS4/ITS5; 5) M30- ITS4/ITS5; 6) M26- ITS4-CgInt; 7) M28- ITS4-CgInt; 8) M29- ITS4-CgInt; 9) M30- ITS4-CgInt; 10) marcador de 100 pares de bases (bp) (Invitrogen).
Las amplificaciones de M26 y M28-M30 fueron secuenciadas y alineadas con las del GenBank, y dieron una homología del 96, 93, 99 y 99% con C. gloeosporioides, respectivamente (gb/HM016794.1; gb/JF487788.1; gb/JF487788.1; gb/FJ755268.1) (BLAST, 2011).
Discusión
Los resultados de este estudio indican que C. gloeosporioides es el agente causal de la antracnosis en pre-cosecha de frutos de litchi de los cultivares Brewster y Mauritius en Oaxaca, México. Los hongos fueron aislados de frutos infectados de manera natural y artificial de ambos cultivares, y formaron un crecimiento miceliar algodonosos blanco a gris claro con masas conidiales de color salmón en PDA. Los síntomas en frutos inoculados fueron muy similares a los observados en campo con infecciones naturales.
En esta investigación, Colletotrichum fue también aislado de frutos asintomáticos de Brewster; resultados similares fueron reportados por Ann et al., (2004), quienes aislaron C. gloeosporioides de flores asintomáticas y frutos inmaduros de litchi cultivar Black leaf. Una vez que los frutos se cosecharon y almacenaron, desarrollaron síntomas de antracnosis, lo que sugirió que este patógeno ataca al litchi a través de infecciones latentes. Nosotros consideramos la probable naturaleza endofítica de C. gloeosporioides en campo.
Como se mencionó antes, la antracnosis de litchi ha sido consignada principalmente como una enfermedad postcosecha (Coates et al., 1994, Liu et al., 2003, y Liu et al., 2006) que resulta de infecciones quiescentes (Li et al., 2005); sin embargo, este hongo se ha reportado como agente causal de la pudrición del pedúnculo y del síntoma de pepper spotting en frutos de litchi en condiciones de campo en Australia (Cooke et al., 2009). Otras causas del obscurecimiento del pericarpio en postcosecha son la pérdida de humedad debido a la disecación del pericarpio (Chen y Hong, 1992; Underhill y Critchley, 1993), degradación de antocianinas y oxidación de sustratos fenólicos por las enzimas oxidasa polifenol (Underhill y Critchley, 1993) y/o peroxidasa (Zhang y Quantick, 1997).
Las especies de Colletotrichum no se pueden reconocer por el rango de hospederos (Sutton, 1980, 1992), por ello la identificación y caracterización de los aislamientos obtenidos en este estudio se realizó basado en características morfológicas (Smith y Black, 1990; Gunnell y Gubler, 1992; Sutton, 1992). De acuerdo a estos criterios cuatro de los 256 aislamientos de Colletotrichum spp., fueron seleccionados (M26, M28-M30) e identificados como C. gloeosporioides Penz; su patrón de crecimiento miceliar concuerda con los descrito por Alvarez (1949); sin embargo, de acuerdo a Smith y Black (1990) y Gunnel y Gluber (1992), este crecimiento es similar a C. fragariae incubado en PDA, sin embargo las colonias de C. fragariae en placas de harina de maíz, harina de avena, o frijol-lima-agar son obscuras con círculos concéntricos y masas conidiales de color rosa (Brooks, 1931). Además, el tamaño promedio de los conidios de M26 y M28-M30 fueron más cortos y más anchos que los reportados en C. fragariae (Smith y Black, 1990; Gunnel y Gubler, 1992).
Basado en las amplificaciones de PCR a través de iniciadores específicos, las especies M26 y M28-M30 se corroboraron como C. gloeosporioides con una similaridad del 95-100% con secuencias reportadas para la misma especie en el banco de genes (Freeman et al., 2000).
Conclusiones
Los síntomas de antracnosis observados en frutos de litchi en Oaxaca fueron causados por Colletotrichum gloeosporioides Penz. La enfermedad se caracterizó por lesiones cafés obscuras y circulares en el exocarpo y endocarpo, pero no en el arilo ni semilla. Este es el primer reporte Colletotrichum gloeosporioides como agente causal de antracnosis de frutos de litchi (Litchi chinensis) en Oaxaca, México.

