










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCuicuilco
versión impresa ISSN 0185-1659
Cuicuilco vol.22 no.64 México sep./dic. 2015
Diversas temáticas desde las disciplinas antropológicas
Evolución de la comunicación vocal y su papel en la estructuración del espacio social y bioacústico en prosimios: una aproximación biosemiótica
María G. Méndez-Cárdenas y Luis Alberto Vargas
Posdoctorante por la Coordinación de Humanidades. Instituto de Investigaciones Antropológicas. Universidad Nacional Autónoma de México.
Instituto de Investigaciones Antropológicas. Universidad Nacional Autónoma de México.
Recepción: 20 de febrero de 2014.
Aprobación: 10 de junio de 2014.
Resumen
El presente artículo es una revisión de los estudios de comunicación vocal llevados a cabo en prosimios; primates monógamos con sistemas sociales de tipo "solitario" considerados los más primitivos, es decir, sus capacidades sociales se aproximan a las condiciones ancestrales del grupo en cuestión. Trabajos ya publicados y recientes, con datos aquí presentados, sobre una de las especies, el lémur deportivo, servirán como introducción al tema de la evolución de la comunicación vocal de los primates y su papel como reguladora de las interacciones sociales que se presentan en diferentes espacios territoriales y nos permitirán establecer puentes entre disciplinas como la lingüística, la semiótica, el comportamiento social y la teoría evolutiva.
Presentaremos una breve revisión de las hipótesis sobre la evolución de la comunicación y la función de las vocalizaciones en un sistema social monógamo caracterizado por una alta territorialidad. Analizaremos si la estructura acústica se asocia con algún entorno o conducta, es decir, si es una señal referencial, qué tipo de información conlleva y cómo se relaciona esto con una función y significado contextual "representacional" utilizando dos modelos: el cooperativo y el competitivo, tanto para datos observacionales como experimentales.
Con base en estudios de paternidad, discutiremos si la teoría de selección de parentesco —la cual ha sido utilizada por Tecumseh Fitch para explicar cómo emergió un sistema de comunicación confiable— generó el desarrollo de una atención coordinada y una comunicación "honesta" (creíble o convincente) en prosimios, el cual pone de manifiesto la presión selectiva sobre las señales acústicas coordinadas. La forma como esto ocurrió es un tema central de las teorías del lenguaje y de su origen. Si las vocalizaciones sirven como reguladoras de la estructuración del espacio social y ecológico, éstas se asociarían a contextos específicos y territorios definidos por su uso.
Mostraremos cómo se efectúan los análisis sobre la estructura vocal de llamados durante la emisión y percepción, desde un enfoque biosemiótico. Examinaremos si las unidades mínimas de información o unidades vocálicas están concatenadas en un sistema protosintáctico como el reportado en otros primates. Finalmente analizaremos hasta qué punto la comunicación animal cumple con las funciones del modelo estructuralista del lenguaje de Jakobson y con las características lingüísticas propuestas por Hockett.
Palabras clave: comunicación vocal, primates, sintaxis, semiótica, evolución.
Abstract
This article provides a review of the studies carried out regarding the vocal communication conducted in prosimians; the monogamous primates with "solitary" social systems are considered the most primitive kind, that is to say, their social skills are closer to the ancestral conditions of the group in question. Data presented here, from previously published and recent works on one of the species: the sportive lemur, serves as an introduction to the topic of the evolution of vocal communication in primates, along with its role as a regulator of social interactions that occur in different territorial areas and allow us to build bridges between disciplines such as linguistics, semiotics, social behavior and evolutionary theory.
The paper presents a brief review of the hypotheses of the evolution of communication and the role of vocalizations in a monogamous social system characterized by high territoriality. We also consider whether the acoustic structure is associated with a specific environment with behavior, that is to say, it means, if it is a reference signal, what type of information is involved and how does this relate to a function and how it is "representational" regarding the contextual meaning through the use of two models: the cooperative and the competitive, regarding both observational and experimental data.
Based on studies of paternity, we discuss whether the theory of kin selection —which was used by Tecumseh Fitch to explain how a trustworthy communication system emerged— generated the development of a coordinated care and "honest" (credible or convincing) communication in prosimians, thus highlighting the selective pressure on the coordinated acoustic signals. How this happened is a central theme regarding the theories of language and origin. If vocalizations serve as regulators of social and ecological structuring of space, then they would also be associated with specific contexts and the territories defined by their use.
We show how the analysis of the vocal structure of calls is carried out —during the broadcast and perception— using a biosemiotic approach. We examine whether the smallest units of information, or vowel units, are concatenated into a proto-syntactic system as reported in other primates. Finally, we discuss how animal communication fulfills the functions of Jacobson's structuralist model of language, along with the linguistic characteristics proposed by Hockett.
Keywords: voice communication, primates, syntax, semiotics, evolution.
That which distinguishes man from the lower animals is not the understanding of articulate sounds, for, as every one knows, dogs understand many words and sentences... It is not the mere articulation which is our distinguishing character, for parrots and other birds possess this power. Nor is it the mere capacity of connecting definite sounds with definite ideas [...] The lower animals differ from man solely in his almost infinitely larger power of associating together the most diversified sounds and ideas; and this obviously depends on the high development of his mental powers.
Charles Darwin, 1871, The Descent of Man1
Introducción
Comprender las reglas que vinculan la comunicación y el comportamiento social que, a su vez, está influido por los patrones espaciales de distribución de los recursos alimenticios y por la amenaza de los depredadores [Crook y Gartlan 1966; Crook 1970; Scheumann et al. 2007], es un prerrequisito esencial para discernir cómo pudo evolucionar un sistema de comunicación entre los primates [McComb y Semple 2005]. Dentro de los factores sociales, ecológicos y de curso vital que contribuyen de manera importante en la variación de los sistemas sociales [Kappeler 1997; Fuentes 2002], la comunicación vocal juega un papel primordial como reguladora de las interacciones sociales y sus mensajes contienen información sobre el medio externo e interno del individuo. Especialmente en primates, el interés en los sistemas de parejas ha sido un aspecto central en estudios de la evolución de la organización social [Schaik y Dunbar 1990; Jolly 1998; Palombit 1999; Schaik y Kappeler 2003]. Un atractivo de los estudios sobre parejas como unidad social se debe al alto índice de especies de primates que viven en pares, así como a su semejanza con la típica organización social en humanos de países industrializados del occidente [Lovejoy 1981].
El lazo de pareja es considerado la unidad basal en modelos evolutivos de muchas sociedades de primates [Fuentes 2002] que evolucionó en varias ocasiones de forma independiente. Su análisis contribuyó a la búsqueda de orígenes evolutivos y de presiones selectivas similares que llevaron a los machos a copular con una sola hembra y cómo esta unión influyó en la adecuación individual [Rasmussen 1981]. En la literatura primatológica, los vínculos entre adultos son descritos como relaciones predecibles estimadas por la tasa de interacción afiliativa, los grados de proximidad y de reciprocidad entre dos individuos [Hinde 1983; Krebs y Davies 1997]. Uno de los despliegues vocales más interesantes son los duetos que se dan sobre todo en aves y primates, en su mayoría con sistemas monogámicos. En este artículo se analizarán las diferentes funciones de dichos duetos.
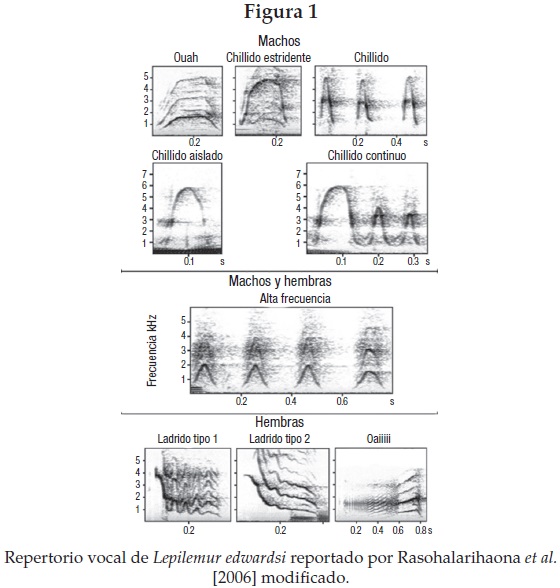
El repertorio vocal específico para cada sexo (véase la figura 1) y la individualidad vocal reportada en Lepilemur edwardsi [Rasoloharijaona et al. 2006] permitió comprobar la estructura y probar experimentalmente la función de los duetos. Con los experimentos de playback1 aquí presentados y el estudio descriptivo sobre la conducta y las interacciones vocales [Méndez y Zimmermann 2009] por primera vez en un prosimio nocturno se pusieron a prueba dos de las hipótesis más controversiales del funcionamiento del dueto: la defensa conjunta de territorio y la atracción y defensa de pareja. Ambas hipótesis se propusieron para explicar el origen y la evolución de los duetos en aves territoriales y primates con vida en pareja.
El paradigma del playback se utiliza para determinar si los individuos discriminan variantes de llamados en una forma significativa funcional [Snowdon 1997] respondiendo conductualmente al estímulo. Estudios sobre la percepción de los llamados indican cómo la variación acústica es clasificada dentro de categorías funcionales con significado para el receptor. Esto nos provee algunas pistas sobre el contenido informativo de las señales [Hauser 1998]. Existen pocos experimentos sobre comunicación vocal en primates en condiciones naturales que hayan examinado cómo la información típica de la especie está codificada en la estructura de dichos signos vocales de los primates [Robinson 1979; Ghazanfar et al. 2001]. Es necesaria información sobre cómo los primates perciben estos llamados y los clasifican, nuestro estudio contribuyó a llenar este vacío.
A los duetos los podemos considerar como sistemas cohesivos de comunicación (consorcio), resultado de la conducta adaptacional plástica para resolver y evitar conflictos conespecíficos y heterospecíficos. Consorcio define a un grupo de organismos conectados vía relación de signos [Kull 2010]. A la estructura de relación espacio-temporal de un consorcio se le llama cronotopo [Bakhatin 1986]. Así, un grupo de individuos es un consorcio si se mantienen juntos mediante lazos comunicativos, es decir, a través de signos: semiosis [Malavasi et al. 2013]. La semiosis es cualquier forma de actividad, conducta o proceso que involucre signos, incluyendo la creación de un significado y una propiedad emergente de nuestro universo. La biosemiótica es un intento por integrar los descubrimientos de la biología y la semiótica, que estudia la producción, acción e interpretación de los signos en el reino físico y biológico [Emmeche 2003], es decir, las bases biológicas de todos los signos y su interpretación.
Antecedentes
Sistema de comunicación en despliegues coordinados
Muchas aves y primates monógamos realizan despliegues vocales conjuntos denominados duetos [Thorpe 1972; Farabaugh 1982; Haimoff 1986; Malacarne et al. 1991; Hall 2004]. Los duetos representan una forma de comunicación acústica interactiva en los cuales un compañero sexual coordina sus vocalizaciones en el tiempo con las de su pareja [Wickler 1980; Wickler y Seibt 1982]. Este llamado interactivo de parejas sexuales ocurre de forma repetida y predecible en el tiempo [Langmore 2002]. Se han identificado duetos en más de 200 especies de aves [Thorpe 1972; Kunkel 1974; Farabaugh 1982]. Además, la mayoría de los primates monógamos, como indris, társidos, gibones, siamangs, monos titi y langures Mentawi despliegan duetos [Marshall y Marshall 1976; Tilson y Tenaza 1976; Haimoff 1986; Pollock 1986; Robinson, et al. 1987; Geissmann 1999; Nietsch 1999; Müller y Anzenberger 2002].
La evolución de los duetos en grupos filogenéticamente independientes sugiere que la selección para formar duetos es intensa [Helversen 1980]. Tanto en aves como en primates se sugiere que los duetos fortalecen los lazos de pareja. Sin embargo, se conoce muy poco sobre la presencia y función de los duetos en mamíferos. En társidos se comprobó que los duetos juegan un papel importante para la comunicación intraespecífica [Nietsch 1999] y probablemente para la atracción de la pareja y su defensa [Nietsch 2003]. Diversas hipótesis funcionales se han puesto a prueba para explicar la evolución de los duetos. El modelo de reunión de pareja reproductiva, para coordinar y sincronizar actividades reproductivas, se predice ocurriría sólo durante el periodo de apareamiento en ranas [Tobías et al. 1998], en sapos [Bush y Bell 1997] y en aves [Stokes y Williams 1968].
El modelo de defensa de la pareja en aves predice que los machos que protegen y retienen a las hembras son acústicamente más responsivos hacia los llamados de ella y hacen duetos cuando un macho intruso está presente [Sonnenschein y Reyer 1983]. El modelo de reunión de pareja se explica en los primates con sistemas cohesivos de pareja quienes invierten más tiempo dueteando por las mañanas, cuando las familias se juntan para dormir, y menos por las tardes, cuando comienzan el forrajeo [MacKinnon y MacKinnon 1980; Nietsch 2003].
Estudios del modelo de defensa coordinada del territorio en aves [Topp y Mennil 2007] muestran que después de la reproducción, las parejas mantienen la reciprocidad comunicativa, lo cual apoya la hipótesis de que los duetos son un despliegue cooperativo que permite defender los recursos ante otros rivales, coordinando la defensa entre parejas [Seibt y Wickler 1977; Hall 2000; Mulder et al. 2003; Rogers et al. 2006]. En primates con sistemas cohesivos y de pareja se estima que los duetos sirven para expulsar a vecinos o desconocidos de un territorio ocupado [Haimoff y Gittins 1985], o cuando un grupo se aproxima o cruza la frontera de otro grupo [Pollock 1975], lo cual indica que puede funcionar como una declaración territorial, vinculada a sitios prominentes del medio ambiente [Wickler 1976].
¿Qué es un sistema social en primates?
Un sistema social incluye tres factores interconectados: la organización social relacionada con el tamaño del grupo, la composición sexual o proporción entre hembras y machos, y la cohesión social (qué tan cercanos espacialmente se encuentran los miembros de la unidad social) [Müller y Thalmann 2000; Kappeler y van Schaik 2002]. La estructura social se relaciona con las alianzas y el patrón de las interacciones sociales, mientras que el sistema reproductivo o de apareamiento tiene correspondencia con el comportamiento sexual y reproductivo [Emlen y Oring 1977] y un componente genético, que describe el éxito reproductivo [Clutton-Brock 1989; Kappeler y Van Schaik 2002]. Como sistemas sociales, existen los monogámicos (pareja), poliándrico (unihembra), poligínico (unimacho), polígamos (múltiples machos y hembras) y pueden ser de tipo cohesivo, fisión-fusión, alianzas macho-hembra, asociaciones patrilineales o matrilineales y sin uniones, es decir, dispersos y solitarios [Clutton-Brock y Harvey 1977; Müller y Thalmann 2000].
La diversidad de los sistemas sociales en primates es enorme y ésta también se presenta dentro de una misma especie, lo cual demuestra no sólo una inercia filogenética (homologías) o "causas últimas", sino también un componente ecológico adaptativo (analogías) o "causas próximas". Estudios científicos apoyan la idea de que el tipo y la intensidad de la competencia por alimentos tiene implicaciones importantes para delimitar el tamaño del grupo, así como el comportamiento social, las relaciones de dominación y los patrones de dispersión en primates [Koenig 2000; Stevenson y Castellanos 2000]. Según Lehman [2006] las restricciones cognitivas pueden limitar el tamaño del grupo.
En cuanto al sistema de apareamiento debemos aclarar que la teoría de la selección sexual asume que ésta actúa de manera diferente para cada sexo. Es decir, si la "mayor" inversión energética para la reproducción la hace la hembra, es ella quien escoge al macho e intenta que éste invierta en el cuidado parental pero, a su vez, el macho tratará de escaparse y seguir copulando [Trivers 1972] sentando así las bases para el conflicto de intereses entre machos y hembras. Darwin [1871] consideraba que la selección sexual tenía dos componentes: la competencia por cópulas y la selección de pareja; centró sus explicaciones en la competencia intrasexual.
Los modelos sociobiológicos sugieren que la monogamia ocurre sólo si los beneficios en la adecuación (supervivencia de los adultos y éxito reproductivo: supervivencia de las crías) pueden compensar a los machos por los costos de perder cópulas adicionales [Trivers 1985; Maynard 1991]. Sin embargo, estudios genéticos de paternidad en sistemas monogámicos de aves y algunos mamíferos señalan un alto porcentaje de cópulas extrapareja en el caso de las hembras, y una baja certeza en la paternidad de los machos, lo cual pone en evidencia cierta promiscuidad de las hembras. Lo anterior sugiere explicaciones alternas como mecanismos conductuales que funcionen como contra estrategias al infanticidio, en el que los machos no puedan discernir entre sus crías y las ajenas. Estas acotaciones darían mayor énfasis al conflicto intersexual, que sigue jugando un papel poco relevante en la teoría de selección sexual [Stumpf et al. 2011].
¿Cómo han evolucionado los sistemas de comunicación?
La variación en la comunicación vocal puede explicarse por diversos factores como el medio ambiente acústico [Wiley y Richards 1978], las propiedades físicas de la transmisión del sonido y sus implicaciones en la evolución del diseño del signo [Wiley 1991; Tubaro et al. 1993], los mecanismos perceptuales y las estrategias de respuesta [Naguib y Wiley 2001]. Entre los primates, los llamados pueden contener información sobre especies y poblaciones, pero también del estatus del individuo y los referentes externos [Cheney y Seyfarth 1980; Zimmermann et al. 1995; Hauser 1996]. Además, la variación morfológica del órgano fonatorio, por ejemplo, el hueso hioideo [Schön 1971], la tráquea [Fitch 1999], los sacos aéreos y laríngeos [Hewitt et al. 2002], determina las características del sonido, así como la capacidad de aprender nuevos repertorios vocales [Catchpole y Slater 1995; Janik y Slater 2000]. Sin embargo, numerosos estudios en mamíferos sugieren que los rasgos genéticos juegan un papel predominante para determinar la estructura acústica de los llamados [Jürgens 1979; Gautier y Gautier 1977].
A pesar de la importancia que pueden tener los factores genéticos en la comunicación vocal, otro aspecto fundamental son los estudios que exploran las reglas que vinculan la comunicación vocal con el comportamiento social [McComb y Semple, 2005], que han propuesto que las vocalizaciones o llamados tienen distintas funciones como agrupar a los compañeros, aumentar la cooperación, mantener alejados a los rivales y reducir los conflictos [Marler 1968]. Dunbar [2003] propuso que el lenguaje surgió como una forma de mantener los lazos sociales en grupos cada vez mayores. Aunado a ello, el nivel de sociabilidad puede determinar el tipo de intercambios vocales y su complejidad [Freeberg 2006].
En contraste Libermann y Corballis [Christiansen y Kirby 2003] sostienen que el advenimiento del bipedalismo fue el primer paso hacia la evolución del lenguaje, debido a las adaptaciones cerebrales que se originaron y dieron lugar a la capacidad de aprender secuencias de movimientos más complejos y de mayor procesamiento. Hauser y Fitch [2003] proponen que los mecanismos que subyacen a la producción y percepción del habla en humanos también las compartimos con otros mamíferos y que no evolucionaron para las funciones actuales del lenguaje sino para otras aplicaciones cognitivas y sociales, como la orientación espacial y el forrajeo [Hauser et al. 2002] en un antepasado común entre el humano y el chimpancé. Esto sugiere que quizás la recursividad pudo surgir de otros dominios como el de la orientación espacial, llevando así esta característica de un dominio específico del lenguaje a uno más general de tipo cognitivo, y plantea que la recursividad que dio origen al lenguaje debió ser la única del pasado humano o bien un producto secundario de otros tipos de reorganización neuronal. Los autores hacen énfasis en la necesidad de realizar estudios comparativos para probar ambas hipótesis.
La biosemiótica en el estudio de la comunicación
La biosemiótica pretende usar conceptos semióticos para contestar preguntas sobre la emergencia del significado biológica y evolutivamente, sobre la intencionalidad y el mundo psíquico; concibe la evolución de la vida y de los sistemas semióticos como dos aspectos de un mismo proceso [Vidales 2009]. Su relevancia para la comunicación es investigar los orígenes del fenómeno semiótico y establecer un puente entre las humanidades y las ciencias naturales para superar el falso dualismo entre cultura y naturaleza a través del entendimiento de la relación entre la "naturaleza interna y externa" [Hoffmeyer apud Kull 1999].
En el estudio de la comunicación, la semiótica se encarga de comprender el intercambio simbólico, y como el símbolo es equívoco, el estudio de la semiótica debe ser de carácter interdisciplinario. Carles Riba [1990] se da a la tarea de integrar teoría y método de la etología en el estudio de la psicología animal, utilizando como herramienta la semiótica de Charles Sanders Peirce como una lógica de la acción. Según la teoría de Peirce [1998] el signo es una pragmática formada por conceptos de índole formal y, en cierta medida, vacíos de contenido, aun cuando es una de sus ventajas. El mismo Peirce afirmó que sus términos no podían permanecer vacíos y que era necesario proporcionar a los conceptos un contenido conductual: "psicologizar la pragmática" [1998].
Morris, en su libro Fundamentos de la teoría de los signos [1938], planteaba que si la conducta es un símbolo, entonces su significado serán las acciones que el animal haga en función del ambiente donde se desarrolle (contexto). Cuando el hecho pertenece al pasado, el significado de la conducta es referencial, si es futuro, el significado es funcional. Con ambas dimensiones se obtiene un repertorio de significados o repertorio vocal y el etograma para la especie no sólo en cuanto a su forma y estructura, sino en lo que respecta a la función de los signos. En el caso de los coros o vocalizaciones sincronizadas, éstas tienen una estructura similar a las de las biofonías descritas por Krause [1987], en las cuales las especies vocalizan al mismo tiempo pero cada vocalización es claramente distinguible, porque los individuos utilizan diferentes intervalos de frecuencias o tiempos. La estructura biofónica aprovecha un importante código en las comunidades acústicas: el sobrelapamiento de vocalizaciones pudiera tener un significado agonístico en el cual se reduce la emisión del sonido [Dabelsteen et al. 1997]. Los individuos quebrantan el código cuando señalizan con un patrón tipo antagonista o cuando forman un nuevo código complementario. En contraste, los individuos forman el código mientras alternan el tiempo de sus vocalizaciones de tal forma que no se crean cacofonías.
Características lingüísticas que comparten primates humanos y no humanos
A diferencia del lenguaje en primates no humanos, el lenguaje humano tiene una doble articulación [André Martinet 1960], es decir, un enunciado se puede descomponer en unidades significativas que a su vez pueden fragmentarse en otras menores no significativas. Hockett [1958] denomina a esta característica "dualidad de patrón", y considera que es uno de los universales lingüísticos. Un mensaje de una lengua natural se construye por medio de segmentos mínimos en dos niveles: la primera articulación del lenguaje sirve para formar enunciados constituidos por "unidades significativas" mínimas o monemas. La segunda está constituida a partir de "unidades distintivas sin significación" o fonemas.
El signo por excelencia es el signo lingüístico y sus características son las siguientes: está dotado de contenido semántico (significado) y de expresión fónica (significante). Une un concepto y una imagen acústica [Saussure 1986]. Los dos aspectos son concomitantes, lo cual se pone de manifiesto en que no hay uno sin el otro. La relación entre significante y significado es arbitraria y necesaria al mismo tiempo. El signo y el enunciado lingüístico se desarrollan en el tiempo, es decir, dos unidades no pueden encontrarse en el mismo punto de la cadena oral, la posición de las unidades es distintiva. Esta característica permite distinguir sistemas como la lengua, cuyo dominio es el tiempo, de sistemas visuales, donde los signos se organizan en el espacio. Hockett [1960] aísla 13 particularidades que singularizan al lenguaje y lo distinguen de otros sistemas de comunicación, e identifica nueve, incluida la semanticidad como compartida con otros organismos y cuatro como únicas del ser humano: 1) la doble articulación o dualidad de patrón antes descrita; 2) el desplazamiento: significados sobre el pasado y futuro o referentes distantes que pueden ser comprendidos; 3) la productividad: nuevas manifestaciones pueden ser rápidamente acuñadas y comprendidas y, 4) la transmisión tradicional: los lenguajes se aprenden y no están genéticamente codificados.
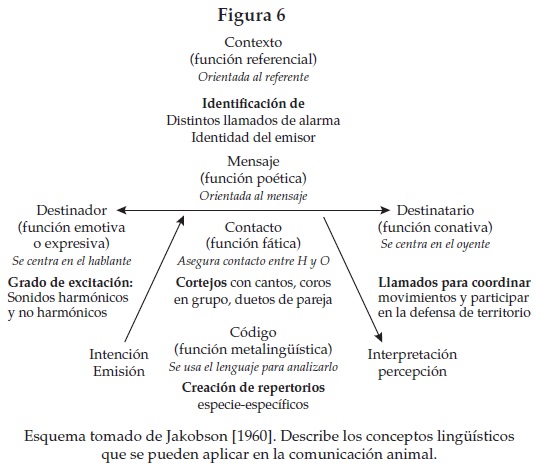
En 1963, Hockett incluyó tres más: la prevaricación, la reflexividad y la habilidad para aprender otras lenguas. Jakobson [1960] determinó las funciones del lenguaje como: 1) denotativa, cognitiva o referencial: orientada al referente, evita confusión entre signo y objeto; 2) expresiva o emotiva: centrada en el hablante, es la manifestación espontánea de emociones de carácter o sociales; 3) conativa o apelativa: centrada en el oyente, se dirige a la inteligencia que organiza la acción en común o la afectividad del receptor, movilizando la participación; el mensaje es un condicionamiento a través de estímulos asociados; 4) fática: asegura el contacto entre el hablante y el oyente, son las formas ritualizadas para prolongar la conversación, hacer presencia y acentuar la adhesión al grupo; el referente es la comunicación; 5) metalingüística: se usa el lenguaje para analizarlo y definir el sentido de los signos remitiéndolo al código del cual extrae su significación y, 6) poética: orientada al mensaje, el cual se convierte en su objeto o referente.
Son varios los trabajos que han tratado de documentar las indudables dotes simbólicas de especies no humanas [Herman et al. 1984; Savage-Rumbaugh et al. 1993; Arnold y Zuberbühler 2006] y más escasos, en cambio, los que han intentado documentar cualidades de tipo combinatorio que pudieran ser razonablemente interpretadas como precursoras de las sintaxis lingüística. Un estudio donde se analizan vocalizaciones concatenadas en monos Campbell llevada a cabo por Ouattara et al. [2009] demuestra que la evolución de una morfología acústica compleja inició en los primates mucho antes de la emergencia en los homínidos, y precede a la evolución de la comunicación intencional. En ese estudio los autores encuentran que las unidades vocales, codificadas en secuencias, están relacionadas con seis contextos específicos, es decir, los individuos siguen un número de reglas cuando concatenan secuencias, y las probabilidades de transición entre tipos vocales no son aleatorias, ni lo es la adición de llamados específicos a secuencias preexistentes para formar una secuencia diferente, o la combinación de secuencias para formar una tercera.
Los autores concluyen que estos primates superaron las restricciones de control vocal a través de la organización combinatoria de secuencias, que el orden de los elementos es importante para reconocer el mensaje, ya que secuencias puestas en sentido inverso o unidades embrocadas disminuyen la reacción, no sólo conductual sino la respuesta neuronal. La respuesta débil a estímulos con orden invertido demuestra que la sensibilidad neuronal a características temporales se extiende más allá de los límites de las sílabas.
Métodos
Registro conductual, vocal y experimentos de playback de un prosimio
A lo largo de un año de trabajo en el noroeste de Madagascar, y con el objetivo de registrar el uso de territorio (ámbito hogareño) y sitios para dormir y comer de 17 individuos de L. edwardsi, a un total de seis parejas adultas se les colocaron radio-collares TW-3 button-cell tags (Biotrack, Dorset, Reino Unido), y dos juveniles y tres crías fueron monitoreadas. Utilizamos radio-telemetría (radio-collares con frecuencias específicas y antenas), un receptor TR-4 y una antena RA-14K (Telonics, Inc., Impala) y a través del método de triangulación obtuvimos un total de 60 puntos de geolocalización por individuo para cada periodo reproductivo (apareamiento) y no reproductivo (gestación y lactancia) y los ubicamos en mapas con el programa Arc View GIS 3.3 (ESRI) [véase Méndez y Zimmermann 2009]. Muestras de tejido cartilaginoso de la oreja fueron obtenidas durante ocho años de un total de 126 individuos, 63 hembras y 63 machos, y de los 17 individuos de este estudio. Las muestras se usaron para extraer el ADN con el método de PCR y llevar a cabo los análisis de paternidad en tres diferentes programas y utilizando nueve microsatélites detectados en un secuenciador megabase 1000 (Amersham Pharmacia) [Leinweber 2008].
Los datos de conducta y las interacciones vocales fueron registrados con una grabadora análoga (Sony Professional WMD6C y un micrófono direccional con filtro Sennheiser ME67/K6). Las grabaciones se registraron en casetes. Las observaciones se realizaron de forma focal y continua durante un mínimo de siete horas y un máximo de 12. Se observó a cada individuo en determinado periodo. Las vocalizaciones fueron digitalizadas utilizando el software Bat Sound Pro Sound analysis 3.31 [Elektronik 2001] y examinadas en Avisoft AviSoft SAS Lab Pro 4.39 [Specht 2003] para medir distintos parámetros como frecuencias, duraciones y armónicos. Con el propósito de probar las hipótesis de defensa cooperativa de territorio y defensa competitiva de pareja, aplicamos experimentos de playback donde emitimos tres categorías de llamados "solos" de hembras, "solos" de machos y duetos y se analizaron en dos grupos: durante los periodos reproductivo y no reproductivo. Los estímulos se emitieron a través de un CD player D-EJ250 Sony situado a dos o cinco metros del sitio donde dormían y conectado a un altavoz (Easy Port FP1pro/CD) que se colocaba de 10 a 15 metros ocultándolo en la vegetación para que no fuera visible para el primate y a una altura de metro y medio a dos metros del suelo. Las respuestas de cada individuo se grababan cuando salían o regresaban al agujero donde dormían, utilizando una cámara nocturna (Sony digital CM 16K, 120x digital zoom). Estas grabaciones se digitalizaron en el programa 8 Pinnacle studio quick start (Pinnacle Systems Inc., 2003) para analizarlas cuadro por cuadro con el programa Interact 8.0 (Mangold Int, 2006).
Análisis de datos
El ámbito hogareño se analizó utilizando Trackasc (software A. Ganzhorn, 1996, unpub.) y el programa Ranges 6 v1.8 (Institute of Terrestrial Ecology, Wareham, Reino Unido) [Kenward et al. 2002] aplicando el método de polígonos mínimos convexos [Méndez y Zimmermann 2009]. Para estudiar si los llamados se asocian a algún contexto, la actividad conductual vinculada a un llamado se clasificó en ocho distintas categorías: descanso, locomoción, alimentación, atención visual, autoacicalamiento, afiliación (acicalamiento a otros), agresión hacia extraños y cuidado de la cría. Para explorar cómo los duetos tienen la función de cooperación y coordinación de las actividades en las parejas, registramos sus actividades durante un minuto antes y después de que vocalizaran y establecimos cuatro categorías asociadas al llamado cuando la pareja muestra: 1) diferentes actividades antes y después de vocalizar (DaDd); 2) similares actividades antes y después de vocalizar (MaMd); 3) las mismas actividades antes de vocalizar y distintas luego de vocalizar (MaDd) y, 4) disímiles actividades antes de vocalizar y las mismas actividades después de vocalizar (DaMd, coordinación).
Todos los eventos de vocalización estuvieron vinculados a una de las cuatro categorías. Para saber si existían diferencias entre éstas, se usó la Anova de Friedman,2 y la prueba de Wilcoxon matched pair.3 Para explorar si la actividad de los llamados en dueto es diferente según el espacio ecológico, se hicieron registros vocales en los sitios de dormir y comer, se calculó la frecuencia de los duetos por individuo en cada lugar y se aplicó la prueba de Mann Whitney U.4 Para comprobar si las unidades vocales están concatenadas de forma no aleatoria, analizamos la organización de nueve tipos de vocalización (figura 1), que combinados se asocian a 10 contextos específicos: llegada al sitio de dormir, forrajeo, encuentro afiliativo macho-hembra (M-H), apareamiento, encuentro agonístico H-H, encuentro agonístico M-M, encuentro agonístico M-H, movimiento H, movimiento M y salto conjunto M-H.
Finalmente para analizar las respuestas de los playbacks, se clasificaron las respuestas en cuatro categorías: no interrupción de la actividad, interrupción (reacción), repuesta no orientada y respuesta orientada (hacia el altavoz). Con el programa Interact se colocaron marcas en los videos para calcular la latencia de respuesta (intervalo entre el comienzo del playback hasta que el sujeto interrumpe la actividad que realizaba) y su duración. Obtuvimos la duración promedio de las respuestas orientadas de cada individuo hacia duetos y solos y lo examinamos para los dos periodos. Para determinar si las respuestas fueron diferentes entre sexos aplicamos una prueba de Mann-Whitney-U para los dos parámetros, se midió duración y latencia de respuesta. La Anova de Friedman nos permitió analizar el efecto de los estímulos del playback sobre la latencia y la duración de respuesta de los individuos. Pruebas de Wilcoxon se utilizaron para explorar si los individuos discriminan entre "solos" y duetos, su respuesta fue distinta a los estímulos, y si existen divergencias según en el periodo reproductivo en el cual se encuentran y entre sexos con una prueba de MannWithney-U.
Resultados
Los resultados obtenidos nos indican que Lepilemur edwardsi tiene un comportamiento social más cohesivo que disperso, donde las parejas muestran un alto traslape de sus territorios corroborando su monogamia social [Méndez y Zimmermann 2009]. Referente a los aspectos genéticos de la paternidad encontramos que, en la población, el mayor porcentaje de madres sociales fueron las madres biológicas y entre los padres existe mayor incertidumbre de paternidad. En las parejas que seguimos durante un año se corroboró que la totalidad de las madres sociales fueron biológicas y sólo uno de los padres sociales no fue el padre biológico [Méndez et al. en proceso].
Datos descriptivos de la conducta vocal y experimentos vocales
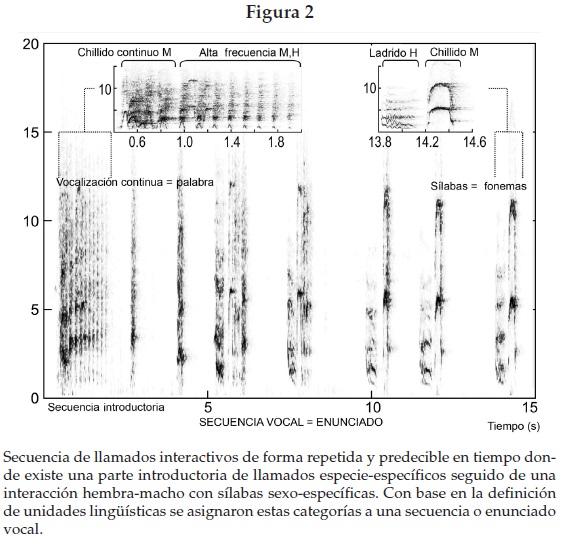
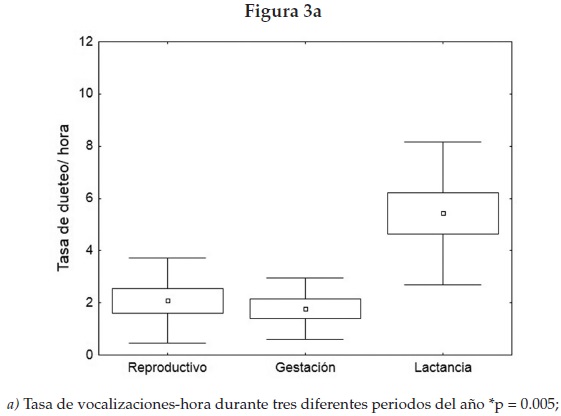
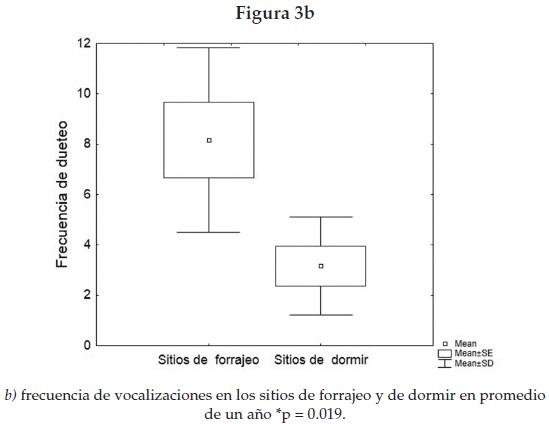
En cuanto a las vocalizaciones, la mayoría de las que se presentaron durante el periodo de cuidado de la cría estaban estructuradas en duetos y se presentaron de forma intensa, repetida y predecible en el tiempo (figura 2) con una mayor ocurrencia durante la etapa de cuidado de la cría (figura 3a) y con mayor frecuencia en los sitios de forrajeo que en los espacios para dormir (Mann Whitney U, Nsd = 6, Nsf = 6, p = 0.019) (figura 3b). De 68 duetos, para los cuales tenemos registrada la ubicación, en un periodo de observación y contacto visual de 103 horas, 72% estuvo relacionado con los sitios de forrajeo.
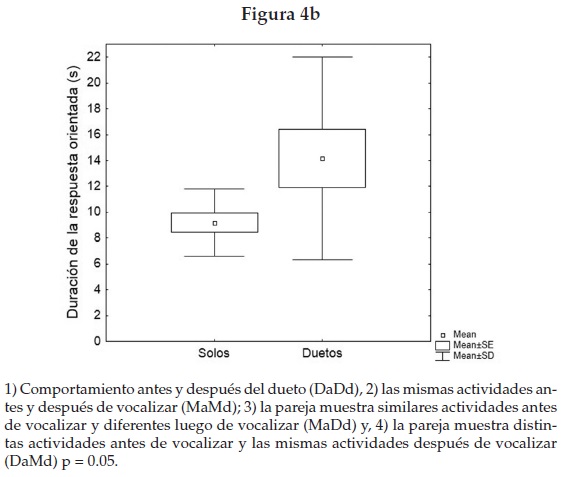
Durante el periodo de cuidado parental (lactancia), encontramos que los duetos se relacionaron con una mayor coordinación de las actividades entre macho y hembra. Esto se analizó considerando el contexto anterior y posterior del macho y la hembra; según la tríada de Pierce, examinamos las categorías posibles con respecto a la emisión del dueto. Sólo en una de éstas la ocurrencia de duetos fue significativamente mayor y se asoció a la coordinación de actividades entre macho y hembra (figura 4a).

Los estímulos de playback emitidos tanto en el periodo reproductivo como no reproductivo, al analizar todos los individuos, no tuvieron efectos significativos sobre la latencia de respuesta (LR) (F-Anova N = 10, df = 3; p = 0.92), tampoco se encontraron diferencias entre machos y hembras (MWU-Test, Nh = 6, Nm = 6; p = 0.81). Sin embargo, sí percibimos efectos considerables en la duración de las respuestas orientadas (F-Anova N = 10, df = 3; p = 0.0087). Además hallamos que los duetos simulados artificialmente en el periodo no reproductivo estimularon respuestas muy distintas y de mayor duración en comparación con los "solos" (Wilcoxon matched pairs test N = 12, p = 0.05) (figura 4b), incluida la respuesta vocal coordinada en pareja. De las seis parejas que mostraron respuestas orientadas hacia los duetos, tres respondieron con duetos.
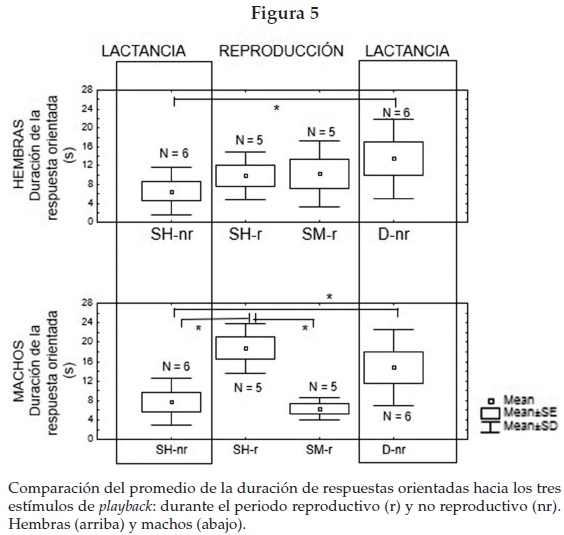
En todos los casos fueron las hembras las que iniciaron la respuesta vocal y los machos se unieron para formar un dueto y las siguieron físicamente aproximándose al altavoz. Solamente una de las seis parejas durante el periodo no reproductivo desplegó un dueto cuando se emitió un solo de hembra; el macho se aproximó al altavoz, comenzó con un llamado de alta frecuencia, high pitch, y la hembra se unió después al dueto. Al analizar las respuestas de machos y hembras por separado para la etapa no reproductiva, encontramos que la duración de respuesta orientada se mantiene significativamente diferente entre playbacks de solos y de duetos (Wilcoxon matched pairs Test en machos (Nm = 6, p = 0.028) en hembras (Nh = 6, p = 0.028) (figura 5). En contraste, el tiempo de respuesta hacia los solos de hembra durante el periodo reproductivo y no reproductivo muestra que las réplicas entre machos y hembras fueron distintas. Los machos despliegan respuestas orientadas más cortas a solos de hembras durante la etapa no reproductiva y más largas en el periodo reproductivo (Wilcoxon matched pairs Test in males (Nm = 5, p = 0.04); mientras que en las hembras no existieron divergencias en las repuestas orientadas a los solos de otras hembras en ambos periodos (Nf = 5, p = 0.22) (véase la figura 5). La duración de respuestas orientadas de machos y hembras también reveló que en estado reproductivo sólo los machos muestran réplicas significativamente cortas hacia otros machos que hacia las hembras (Wilcoxon matched pair Test (Nm = 5, p = 0.043), mientras que las hembras no difieren en el tiempo de respuestas hacia solos de machos o de hembras (Nf = 5, p = 0.68) (véase la figura 5).
Análisis teórico de la función de los duetos
Utilizando el modelo propuesto por Jakobson [1960], a continuación mostramos una comparación entre las funciones propuestas por este autor y las analizadas en este estudio (figura 6). Jakobson propone que de la comunicación afectiva se pasa a la colaboración práctica, que es una coordinación y sincronización del trabajo en común, la cual postula una codificación y socialización del mensaje a expensas de su contenido informacional. En el comportamiento de los primates podemos corroborar la presencia de una atención (proceso cognitivo) y una comunicación (de tipo afectiva), así como una colaboración (práctica) o defensa territorial. Según los análisis que se han llevado a cabo en este trabajo y en otros estudios de la comunicación entre primates, de las seis funciones, sólo de las dos últimas no se tienen pruebas ni empíricas ni experimentales para incluirlas en el lenguaje de los primates. Las pruebas que sustentan dicha propuesta se muestran en la siguiente figura.
Análisis semántico de los duetos
En el Lepilemur deportivo encontramos que los tipos de vocalizaciones se concatenan en secuencias interactivas que podemos dividir en los siguientes contextos: afiliativos, agonísticos, de locomoción y de alimentación, asociados a secuencias específicas de llamados, sexo-específicos para cada contexto, descritos en el siguiente cuadro.
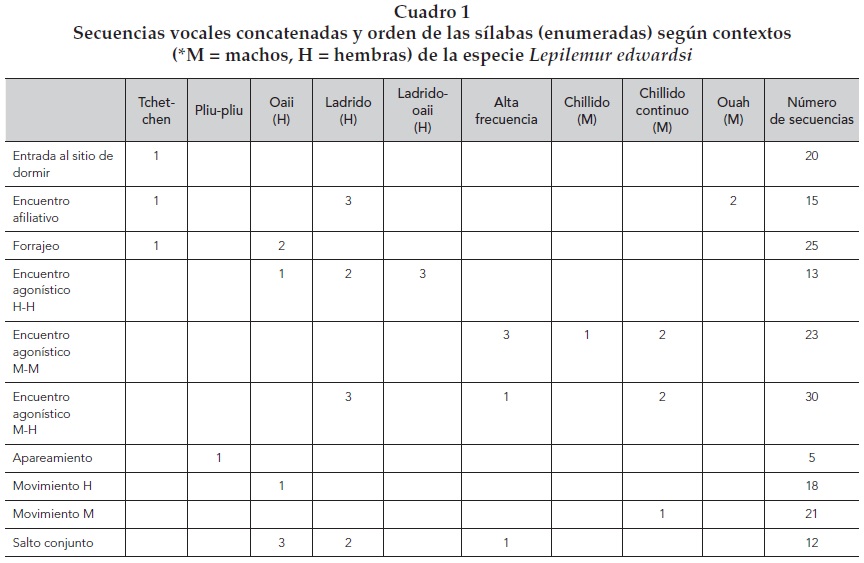
Lo que observamos es que en encuentros afiliativos, de descanso y forrajeo no se presentan ni el chillido ni el chillido continuo ni el de alta frecuencia, pero para los encuentros agonísticos intrasexuales existen claras diferencias entre machos y hembras, las primeras van a utilizar oaii, ladridos y la mezcla de ambos al final, mientras que los machos comienzan con un chillido, el cual se vuelve continuo y termina con una mayor intensidad de alta frecuencia. Los encuentros agonísticos intersexuales inician en su mayoría con un llamado de alta frecuencia, seguido por un chillido continuo y ladridos de baja intensidad.
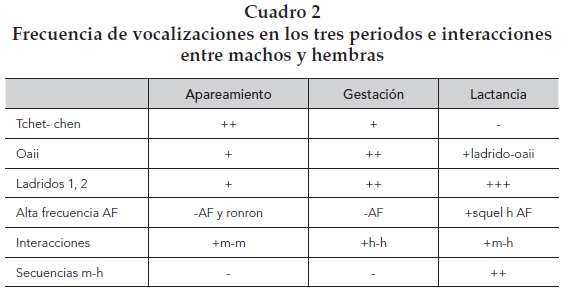
Otro hallazgo interesante, considerando el tipo de vocalizaciones que mostraban mayor ocurrencia, fue que al comparar su ocurrencia, a lo largo de los tres periodos reproductivos, encontramos que durante la etapa de apareamiento prevaleció la vocalización tchet-chen, la cual disminuyó durante la lactancia. En cambio, los ladridos se incrementaron coincidiendo con ella. Otra vocalización interesante fueron los oaii, éstos se presentaron con mayor frecuencia sólo durante la gestación, y los llamados de alta frecuencia aumentaron ligeramente en la lactancia pero, sobre todo, combinados con distintos llamados. Durante el apareamiento se registraron más interacciones vocales entre machos, en la gestación entre hembras y en la lactancia entre machos y hembras.
Discusión y conclusiones
Los duetos y su evolución en primates con lazos de pareja: señales "honestas" y selección sexual
De acuerdo con el modelo monogámico propuesto para aves, se esperaba que las hembras y machos juveniles migraran del nicho familiar a distancias similares para evitar la endogamia, eso no se sustenta en el presente trabajo. Las hembras migraron menos que los machos, asemejándose más a un sistema matrilineal pero de vida en pareja (Méndez et al., en preparación).
Sin embargo, el traslape de los ámbitos hogareños de las parejas encontrado [Méndez y Zimmermann 2009] sugiere una monogamia espacial [Müller y Thalaman 2000; Fietz 1999; Schülke et al. 2004; Harcout y Nash 1986]. Lo mismo se aplica a los datos genéticos de paternidad: una monogamia no sólo social sino biológica, donde las hembras llevan a cabo cópulas extra pareja. Estos hallazgos permiten ilustrar los pasos previos hacia un sistema cohesivo de vida en pareja. Los resultados de paternidad indican que en las hembras la maternidad fue corroborada, pero no para la paternidad, en las cuatro parejas con crías, solamente tres machos fueron los padres biológicos, es decir, las hembras mantienen cópulas extrapareja. La baja probabilidad de paternidad, pero también de maternidad (las madres que crían no fueron 100% las madres biológicas) en los análisis genéticos de la población general, muestran que los machos son incapaces de monopolizar la reproducción de las hembras y pueden cuidar a crías de otro machos, o bien que en ausencia de uno de los padres, los juveniles y otras hembras ayudan en la vigilancia de las crías.
Estudios en primates nocturnos muestran que las señales acústicas controlan la cohesión y coordinación intragrupal [Braune et al. 2005; Rasohalarijaona et al. 2006] y nuestros resultados (figura 4) apoyan estos hallazgos [Méndez y Zimmermann 2009]. El lémur deportivo sincronizó significativamente más sus actividades después de que macho y hembra producen un despliegue vocal coordinado en "dueto". Los duetos analizados actúan como un mecanismo para regular actividades sociales como locomoción, alimentación y supervisión del área, como una forma de mantener y fortalecer el lazo de pareja, descrito previamente en primates con sistemas de tipo cohesivo y vida en pareja como en gibones [Geismann 1999].
El modelo de comunicación "honesto" y engañoso en animales [Gouzoules y Gouzoules 2002] se ha fundamentado con la idea de costos. Así, el emisor y el receptor se beneficiarán a través de una comunicación creíble y no ambigua sobre sus atributos [Smith 1977], donde las señales costosas serán la base para la comunicación "honesta", siempre y cuando la evolución de las señales tenga lugar en un contexto de conflicto de intereses. Zahavi [1975] sugiere que la "honestidad" podrá mantenerse en un sistema comunicativo sólo si los signos son costosos en alguna forma. Otra condición es cuando no existe conflicto de intereses, es decir, cuando el emisor y el receptor se favorecen de interacciones coordinadas en las cuales los individuos interactúan repetidamente y utilizan interacciones pasadas para estimar la legitimidad de los signos, entonces un sistema de señales no costoso puede evolucionar [Di Paolo 1997].
El presente estudio propone que la sincronización de las actividades sociales después del despliegue vocal en dueto puede beneficiar a ambos sexos debido a que comparten recursos territoriales y son capaces de proteger su descendencia a través de despliegues conjuntos. Estos resultados, junto con la extensa permanencia de los lazos de pareja observados, pueden explicarse por un origen no costoso de despliegues vocales coordinados, con los cuales los individuos no estiman la calidad (atributos) pero sí la confianza de señales verosímiles de larga trayectoria, basados, quizás, en un "compromiso" expresado en términos humanos como la dedicación y esfuerzo para coordinarse vocal y socialmente, donde parece no haber conflicto.
Este aprendizaje de señales "honestas", es decir, que no tienen una intención diferente o adicional a la explícita, surgió a través de las interacciones sociales en las cuales llamados individuamente distintos producen efectos específicos en el receptor [Janik y Slater 2000], que a su vez aprende a producir respuestas adaptativas. El aprendizaje podría ser específico, según la historia de las interacciones entre los individuos involucrados, como se muestra en los cantos muy sincronizados de aves y primates [Wickler 1980; Geissmann 1999]. Así, las vocalizaciones contienen características acústicas que permiten identificar individuos y la función primaria de los signos vocales influir en los receptores y modular su comportamiento [Owren y Rendall 1997], es decir, hay una intencionalidad.
En el lémur deportivo, los parámetros relacionados con la frecuencia enfatizada y las propiedades temporales muestran una alta especificidad individual [Rasoloharijaona et al. 2006] en llamados como el "chillido aislado" y el "chillido continuo" de los machos y el "ladrido-oaii" de las hembras (figura 1). Encontramos que estos llamados fueron utilizados por las parejas para generar los duetos. Esto nos sugiere que pueden jugar un papel en la identificación social y espacial de los individuos. Los resultados de paternidad, junto con los despliegues vocales de tipo cooperativo y de producción no costosa, replantean la hipótesis sobre el origen del lenguaje con base en una forma de comunicación altamente estereotipada y compleja, que emergió como un sistema de comunicación confiable [Fitch y Hauser 2002]. Los duetos se originaron no necesariamente por la selección de parentela [Fitch 2010], sino por medio de la selección sexual de pareja, en circunstancias tales como la defensa conjunta del territorio y de las crías, lo cual daría lugar al desarrollo de una atención coordinada y una comunicación honesta, en donde la paternidad social adquiere mayor relevancia.
Modelos para explicar la evolución de los duetos
De los modelos propuestos para explicar la evolución de los duetos, el de defensa conjunta de territorio fue sustentado por nuestros resultados y no así el de competencia sexual, que también se puso a prueba con los experimentos de playback. Estos resultados muestran que los solos de hembras durante el periodo reproductivo, probablemente tengan la función de atraer a los machos; y en la etapa no reproductiva, quizá las hembras representen competencia para las que están lactando, aunque solamente observamos una reacción coordinada de macho y hembra ante la intrusión acústica de una hembra. En la fase reproductiva hay una baja frecuencia de duetos, y durante los experimentos ni los machos ni las hembras emitieron duetos, ni como una forma de competencia ni de retención de la pareja, cuando otros machos o hembras se introdujeron en su campo acústico. Sin embargo, se necesitan más estudios para comprender las preferencias de machos y hembras hacia ciertas vocalizaciones que podrían indicar las cualidades o atributos de la pareja.
Los resultados también indican que los duetos se emiten durante conflictos territoriales (observación personal) y con una frecuencia significativamente mayor en los sitios de forrajeo que en los de dormir (figura 3b). Así la pareja puede optimizar su éxito reproductivo con una defensa conjunta del territorio que provea suficientes hojas frescas ricas en proteínas para la hembra durante la lactancia. En el periodo reproductivo, una hembra mostró una reacción agresiva y vocal hacia un playback de otra hembra, pero este conflicto intrasexual [Mitani 1984 y 1987] no fue apoyado por el macho, es decir, no hubo dueto. En contraste, durante la etapa de lactancia un macho defendió a su hembra y respondió agnósticamente al playback de un solo de hembra intrusa, y se produjo el dueto, lo cual apunta a que las hembras pudieran ser un factor mayor de riesgo para las hembras y sus crías, que los machos. Mayor sentido de territorialidad y agresividad se señala en hembras de peces y reptiles, e infanticidio por hembras en lobos [Stockley y Campbell 2013; Forsgren et al. 2004; While et al. 2009; McLeod, 1990].
En resumen, nuestros resultados sugieren un sistema social más cohesivo de lo que se pensaba, con mayor inversión de tiempo en pareja, coordinándose vocalmente durante la crianza, y menor cooperación durante el periodo reproductivo, y casi inexistente para la etapa de gestación. Por lo cual concluimos que los duetos son un despliegue cooperativo utilizado para la defensa de recursos durante la etapa de mayor estrés nutricional de las hembras, que es cuando amamantan a sus crías, con lo que se reduciría el riesgo de infanticidio, que se ha reportado en los primeros meses de vida [Rasohalarijaona et al. 2000]. Sin embargo, no se descarta que los duetos se encuentren bajo selección sexual, pues según la hipótesis de [Thalmann 2001], la defensa de los recursos alimentarios son una inversión indirecta de los machos para encontrar pareja, reproducirse y alimentar a las crías. Para examinar si la función de los duetos en otras especies coincide con nuestros hallazgos y explorar si su origen es cooperativo más que competitivo, como se encontró en este trabajo, serán necesarios más estudios en primates con distintos sistemas sociales y comparar los periodos reproductivos.
La biosemiótica en el estudio de la comunicación: características lingüísticas que comparten primates humanos y no humanos
La transición que existe de una respuesta orientada después de emitir artificialmente una vocalización de un intruso en el territorio de una pareja y que ésta sea percibida auditivamente, a una reacción vocal coordinada y de defensa, motivada únicamente por este llamado, corresponde a la transición de una semiosis indexical (reactiva espacio-temporalmente) a una semiosis simbólica. El objeto de la señal, en este último caso, no es un objeto sino una clase de objetos (los vecinos y los extraños) y por ello no hay necesidad de que exista un evento singular. Es decir, si existe un índice umbral, mayor al símbolo, entonces, sería posible identificar conductualmente la transición a partir de un objeto que es eventual a un objeto que es de una clase reconocible, por ejemplo, un objeto que no necesita estar presente como un objeto o referente externo particular.
En este estudio, el patrón acústico que definimos como un consorcio o comunicación coordinada en duetos, también puede describirse como un topo-sonido, cuando está asociado a sitios específicos como los de forrajeo y ha sido reportado en los límites territoriales de los grupos o parejas (observación personal) sociales. En aves, el topo-sonido se ha definido como un canto coordinado de distribución no aleatoria que crea un medio ambiente sónico común [Farina 2011]. Esto permite a las aves compartir recursos.
En el caso de los primates también revela y comunica las fuentes de recursos y los sitios de seguridad o guaridas, pero en este caso, aunque la información se ha distribuido con toda la comunidad, los recursos son compartidos únicamente con individuos del mismo grupo social, que alertan a los individuos fuera de los territorios definidos. Por ello, el topo-sonido está dirigido por procesos conductuales [Peake 2005]. La hipótesis de la existencia de un eco-campo o socio-campo común nos provee de un modelo prometedor, con el objetivo de explicar algunos patrones acústicos tanto en aves como en primates, donde la distribución heterogénea de cantos y llamados no puede explicarse simplemente por una vía causal o la vegetación y la distribución de los recursos, sino por las interacciones sociales en el tiempo (estacionalidad) y el espacio (superposición de territorios hogareños).
Por otro lado, el esquema de interpretación basado en el contexto anterior, señal (estímulo), contexto posterior, es una representación de la tríada semiótica de Peirce: objeto, signo e intérprete; del eje del tiempo y de las secuencias de comportamiento. La semántica, entonces, tendrá dos dimensiones: una referencial, que parte de la emisión como resultado de un ambiente previo percibido por las estructuras receptoras del animal (estudio observacional), y la otra funcional, que busca averiguar cómo se recibe el efecto o estímulo, o en qué punto motiva una respuesta (estudio experimental). Con ambas dimensiones obtuvimos un repertorio de significados o repertorio vocal y el etograma específico para la especie en cuanto a la estructura y la función.
Dado que los significados se infieren del uso secuencial de los signos en un determinado contexto, el observador puede interpretar de distintos modos una misma señal, si el análisis se hace con base en ésta y la relación de su estado interno o el contexto de emisores y receptores. Para la biología la señal también nos informa de los aspectos adaptativos de la especie, un mismo signo tiene varias funciones (contextos) o bien una función (contexto) puede conllevar distintos signos. Esto puede confundirse con una especie de zoo-pragmática que estudia la dimensión de la semiosis para la cual el significado depende del contexto; aunque esto representa una limitación metodológica, porque las descripciones de significado no pueden hacerse con los propios signos animales, sí podemos considerar como signos la representación gráfica de los sonidos (espectrogramas).
Finalmente, con los avances en el estudio de la comunicación animal, proponemos qué características del lenguaje consideradas exclusivamente humanas se extiendan al lenguaje animal, para brindar la oportunidad de corroborarlas científicamente. Nos referimos a la dualidad de patrón, fundamentado en la capacidad combinatoria o protosintaxis de unidades mínimas vocálicas en unidades distintivas con una función tanto en el dueto como en las diferentes combinaciones vocales asociadas a contextos específicos. El desplazamiento, referentes distantes que pueden ser comprendidos y puestos en evidencia por los experimentos de playback donde se presentan objetos y sujetos lejanos en tiempo y espacio, y que aun sin su presencia la respuesta conductual es congruente. La productividad, donde nuevas manifestaciones pueden ser comprendidas y la transmisión tradicional, la cual se refiere a que los lenguajes se aprenden y no están genéticamente determinados, se evidencia por la capacidad de incrementar y cambiar los repertorios vocales dependiendo de los periodos reproductivos.
Biólogos, antropólogos, etólogos, psicólogos y bio lingüistas necesitamos elaborar un catálogo paciente y exhaustivo de comportamientos de la especie y su repertorio vocal, sobre el cual hacer a posteriori interpretaciones contexto-dependientes y comparaciones onto y filogenéticas, y crear un código como contexto global de las conductas, es decir, un etograma-código donde el enfoque biosemiótico tendrá un papel fundamental.
Asimismo, se deben incrementar los estudios de la combinación de unidades acústicas sin significado, en secuencias con significado, y analizar las reglas de combinación y sus restricciones. Dado que los mecanismos que subyacen —la producción vocal y la percepción— incluyen la formación de algún tipo de representación mental sobre el estímulo que evoca el llamado, esta información determina cómo un llamado se produce y cómo será interpretado. Es necesario considerar y utilizar la representación mental de Peirce en la cual se sustituye mente por "semiosis" o por producción de significado y pensamiento por "signo".
La aproximación biosemiótica nos ayuda a reflexionar sobre la semiosis y la comunicación como elementos centrales de los procesos de organización y desarrollo de la vida, así como todo organismo vivo requiere como condición mínima de su existencia la capacidad de interactuar a través de signos, necesita de procesos de semiosis interna y externa y procesos socio-cognitivos donde la información sea interpretada y traducida a los códigos indispensables para su supervivencia. La interpretación de la información interna (biológica) y la información compartida entre organismos (biosociales) genera la evolución de la vida misma. Es el proceso evolutivo de los sistemas semióticos, cercanos a las particularidades biológicas y a las codificaciones sociales.
Agradecimientos
Se agradece a la Coordinación de Humanidades de la Universidad Nacional Autónoma de México (UNAM) por la beca posdoctoral, otorgada a MGMC, al Grupo Académico de Antropología Física del Instituto de Investigaciones Antropológicas y en particular el apoyo académico de la profesora, doctora Elke Zimmerman, durante la tesis doctoral de MGMC financiada por el DAAD.
Referencias
Arnold, Kate y Klaus Zuberbühler 2006 Semantic Combinations in Primate calls. Nature (441): 303. [ Links ]
Braune, Pia, Sabine Schmidt y Elke Zimmermann 2005 Spacing and Group Coordination in a Nocturnal Primate, the Golden Brown Mouse Lemur (Microcebus ravelobensis). Behav. Ecol. Sociobiol. (58): 587-596. [ Links ]
Catchpole, Clive y Peter Slater 1995 Bird Song: Biological Themes and Variations. Cambridge University Press. Cambridge. [ Links ]
Cheney, Dorothey y Robert R. Seyfarth 1990 How Monkeys See the World: Inside the Mind of Another Species. University of Chicago Press. Chicago. [ Links ]
Clutton-Brock, Tim 1989 Mammalian Mating Systems. Proc. Roy. Soc. Lond, B 236: 339-372. [ Links ]
Clutton-Brock, H. Tim y Paul H. Harvey 1977 Primate Ecology and Social Organization. J. Zool. Lond (183): 1-39. [ Links ]
Corballis, Michael 2003 From Hand to Mouth: the Gestural Origins of Language, en Language Evolution, M. H. Christiansen y S. Kirby (eds.). Oxford University Press. Oxford. [ Links ]
Crook, John 1970 The Socio-Ecology of Primates, en Social Behaviour in Birds and Mammals, J. Crook (ed.). Academic Press. Londres: 103-166. [ Links ]
Crook, John y Steven Gartlan 1966 Evolution of Primate Societies. Nature (210): 1200-1203. [ Links ]
Dabelsteen, Torben et al. 1997 The Signal Function of Overlapping Singing in Male Robins. Animal Behaviour (53): 249-256. [ Links ]
Darwin, Charles 1871 The Descent of Man and Selection in Relation to Sex. John Murray. Londres. [ Links ]
Di Paolo, Ezequiel 1997 Social Coordination and Spatial Organization: Steps Towards the Evolution of Communication, en Proceedings of the Fourth European Conference on Artificial Life (ECAL'97), P. Husbands e I. Harvey (eds.). MIT Press-Bradford Books. Cambridge: 464-473. [ Links ]
Dunbar, Robin 2003 The Social Brain: Mind, Language, and Society in Evolutionary Perspective. Annu. Rev. Anthropol (32): 163-181. [ Links ]
Emlen, Stephen y Lewis Oring 1977 Ecology, Sexual Selection, and the Evolution of Mating Systems. Science (197): 215-223. [ Links ]
Farabaugh, Susan 1982 The Ecological and Social Significance of Duetting", en Acoustic Communication in Birds, Donald Kroodsma y Edward Miller (eds.). Ac Press. Nueva York: 85-124. [ Links ]
Farina, Almo, Emanuele Lattanzi, Rachele Malavasi, Nadia Pieretti y Luigi Piccioli 2011 Avian Soundscapes and Cognitive Landscapes: Theory, Application and Ecological Perspectives. Landscape Ecology, 26(9): 1257-1267. [ Links ]
Fitch, Tecumseh 1999 Acoustic Exaggeration of Size in Birds by Tracheal Elongation: Comparative and Theoretical Analyses. J. Zool (248): 31-49. Discussed in News & Views, Natures (399): 109. [ Links ]
Fitch, Tecumseh y Marc Hauser 2002 Unpacking "Honesty": Vertebrate Vocal Production and the Evolution of Acoustic Signals, en Acoustic Communication, A. M. Simmons, R. R. Fay y A. N. Popper (eds.). Springer. Nueva York: 65-137. [ Links ]
Fichtel, Claudia, Kurt Hammerschmidt y Uwe Jürgens 2001 On the Vocal Expression of Emotion. A Multi-Parametric Analysis of Different States of Aversion in the Squirrel Monkey. Behaviour, 138 (1): 97-116. [ Links ]
Fietz, Joanna 1999 Monogamy as a Rule Rather than a Exception in Nocturnal Lemurs: the Case of the Fattailed Dwarf Lemur, Cheirogaleus Medius. Ethology (105): 259-272. [ Links ]
---------- 2010 The Evolution of Language, Cambridge University Press. Cambridge. [ Links ]
Forsgren, Elisabet et al. 2004 Unusually Dynamic Sex Roles in a Fish. Nature (429): 551-554. <doi:10.1038/nature02562> [ Links ].
Freeberg, Tood 2006 Social Complexity Can Drive Vocal Complexity: Group Size Influences Vocal Information in Carolina Chickadees. Psychological Science (17): 557-561. [ Links ]
Fuentes, Agustín 2002 Patterns and Trends in Primate Pair Bonds. Int J Primatol (23): 953-978. [ Links ]
Ganzhorn, Jörg 2002 Distribution of a Folivorous Lemur in Relation to Seasonally Varying Food Resources: Integrating Quantitative and Qualitative Aspects of Food Characteristics. Oecología (131): 427-435. [ Links ]
Gautier, J.P. y A. Gautier 1977 Communication in Old World Monkeys, en How Animals Communicate, T. A. Seboek (ed.). Indiana University Press. Bloomington, Indiana: 890-964. [ Links ]
Geissmann, Thomas 1999 Duet Songs of the Siamang, Hylobates Syndactylus: II. Testing the Pair-Bonding Hypothesis During a Partner exchange. Behaviour (136): 1005-1039. [ Links ]
Ghazanfar, Asif et al. 2001 The Units of Perception in the Antiphonal Calling Behavior of Cotton-Top Tamarins (Saguinus oedipus): Playback Experiments with Long Calls. J Comp Physiol A(187): 27-35. [ Links ]
Gouzoules, Harold y Sarah Gouzoules 2002 Primate Communication: by Nature Honest, or by Experience Wise? Int J Primatol (23): 821-848. [ Links ]
Haimoff, Elliott 1986 Convergence in the Duetting of Monogamous Old World Primates. J Hum Evol (15): 51-59. [ Links ]
Haimoff Elliot y Paul Gittins 1985 Individuality in the Songs of Wild Agile Gibbons (Hylobates agilis) of Peninsular Malaysia. Am J Primatol (8): 239-247. [ Links ]
Hall, Michelle 2000 The Function of Duetting in Magpie-Larks: Conflict, Cooperation or Commitment? Anim Behavior (60): 667-677. [ Links ]
---------- 2004 A Review of Hypotheses for the Functions of Avian Duetting. Behavior Ecol Sociobiol (55): 415-430. [ Links ]
Hamilton, William 1964 The Genetical Evolution of Social Behavior. Int J theor Biol (7): 1-16. [ Links ]
Hauser, Marc 1996 The Evolution of Communication, Cambridge, MIT Press. Massachusetts. [ Links ]
---------- 1998 Functional Referents and Acoustic Similarity: Field Playback Experiments with Rhesus Monkeys. Animal Behavior (55): 1647-1658. [ Links ]
Hauser, Marc, Noam Chomsky y Tecumseh Fitch 2002 The Faculty of Language: What is it, Who has it, and How Did it Evolve? Science (29822): 1569-1579. [ Links ]
Hauser, Marc y W. Tecumseh Fitch 2003 What are the Uniquely Human Components of the Language Faculty?, en Language Evolution, M. H. Christiansen y S. Kirby (eds.). Oxford University Press. Oxford. [ Links ]
Helversen, Dagmar von 1980 Structure and Function of Antiphonal Duets. Proc Int Ornithol Congress Berl (17): 682-688. [ Links ]
Herman, Louis M., Richards Doublas G. y James P. Wolz 1984 Comprehension of Sentences by Bottlenosed Dolphins. Cognition (16): 129-219. [ Links ]
Hewitt, Gwen, Ann MacLarnon y Kate Jones 2002 The Functions of Laryngeal Air Sacs in Primates: a New Hypothesis. Folia Primatol (73): 70-94. [ Links ]
Hinde, Robert 1983 Primate Social Relationships: An Integrated Approach, Blackwell, Oxford. [ Links ]
Hockett, Charles 1958 A Course in Modern Linguistics. The Macmillan Company. Nueva York. [ Links ]
---------- 1960 Logical Considerations in the Study of Animal Communication, en Animal Sounds and Communication, W. E. Lanyon y W. N. Tavolga (eds.). American Institute of Biological Sciences. Washington: 392-430. [ Links ]
---------- 1963 The Problem of Universals in Language, en Universals of Language, J. Greenberg (ed.). MIT Press. Cambridge: 1-29. [ Links ]
Jakobson, Roman 1960 Linguistic and Poetics, en Style in Language, T. A. Seabeok (ed.). MIT Press. Cambridge (traducción: "Lingüística y poética". Ensayos de lingüística general. Seix Barral. Barcelona. 1981: 347-395). [ Links ]
Janik, Vincent y Peter Slater 2000 The Different Roles of Social Learning in Vocal Communication. Animal Behavior (60): 1-11. [ Links ]
Jolly, Alison 1998 Pair-Bonding, Female Aggression and the Evolution of Lemur Societies. Folia Primatol, 69 (sl.1): 1-13. [ Links ]
Jürgens, Uwe 1979 Neural Control of Vocalizations in Nonhuman Primates, en Neurobiology of Socialcommunication in Primates, H. D. Steklis y M. J. Raleigh (eds.). Academic Press. Nueva York: 11-44. [ Links ]
Kappeler, Peter 1997 Determinants of Primate Social Organization: Comparative Evidence and New Insights from Malagasy Lemurs. Biological Revue Cambridge Philosophical Society (72): 111-151. [ Links ]
Kappeler, Peter y Carl van Schaik 2002 Evolution of Primate Social Systems. Int J Primatol (23): 707-740. [ Links ]
Kenward, Robert , South Andy y Sven Walls 2002 Ranges 6: for the Analysis of Tracking and Location Data. Anatrack Ltd, Wareham. Dorset, Reino Unido. [ Links ]
Koenig, Andreas 2002 Competition for Resources and its Behavioral Consequences Among Female Primates. Int J Primatol (23): 759-783. [ Links ]
Krause, B. L. 1987 Bio-Acoustics: Habitat Ambience and Ecological Balance. Whole Earth Review (57): 14-18. [ Links ]
Krebs, John y Nicholas Davies 1997 Behavioral Ecology. 4a ed. Blackwell. Londres. [ Links ]
Kull, Kalevi 1999 Biosemiotics in the Twentieth Century: a View from Biology, en Semiótica, 127. Walter de Gruyter. Berlín: 385-414. [ Links ]
---------- 2010 Ecosystems are Made of Semiosic Bonds: Consortia, Umwelten, Biophony and Ecological Codes. Biosemiotics, 3(3): 347-357. [ Links ]
Kunkel, Peter 1974 Mating Systems of Tropical Birds: the Effects of Weakness or Absence of External Reproduction-Timing Factors with Special Reference to Prolonged Pair Bonds. Z Tierpsychol (34): 265-307. [ Links ]
Langmore, Naomi 2002 Vocal Duetting: Definitions, Discoveries and Directions. Trends Ecol Evol (17): 451-452. [ Links ]
Lehman, Shawn 2006 Conservation Biology of Malagasy Strepsirhines: a Phylogenetic Approach. Am J Phys Anthropol (130): 238-253. [ Links ]
Leinweber, Ina 2008 Soziogenetische Strukturen in Einer Population Freilebender, Milne Edwards Wieselmakis (Lepilemur edwardsi). Diplomarbeit, Fachbereich Biologie der Universität Hannover, Referentin: PD Dr. Ute Radespiel. [ Links ]
Lieberman, Philip 2003 Motor Control, Speech, and the Evolution of Human Language, en In Language Evolution, M. H. Christiansen y S. Kirby (eds.). Oxford University Press. Oxford. [ Links ]
Lovejoy, Owen 1981 The Origins of Man. Science (211): 341-350. [ Links ]
Malacarne, Giorgio, Marco Cucco y Camanni 1991 Coordinated Visual Displays and Vocal Duetting in Different Ecological Situations Among Western Palearctic Non-Passerine Birds. Ethol Ecol Evol (3): 207-219. [ Links ]
Marler, Peter 1968 Aggregation and Dispersal: Two Functions in Primate Communication, en Primates: Studies in Adaptation and Variability, P. C. Jay (ed.). Holt, Rinehart & Winston. Nueva York: 420-438. [ Links ]
Marshall, Joe y Elsie Marshall 1976 Gibbons and Their Territorial Songs. Science (193): 235-237. [ Links ]
Martinet, André 1960 Éléments de linguistique générale. Armand Colin. París. [ Links ]
Maynard, Smith 1991 Honest Signaling: the Philip Sidney Game. Animal Behavior (42): 1034-1035. [ Links ]
McComb, Karen y Stuart Semple 2005 Coevolution of Vocal Communication and Sociality in Primates. Biol. Lett. (1): 381-385. [ Links ]
McLeod, Peter J. 1990 Infanticide by Female Wolves. Canadian Journal of Zoology, 68(2): 402-404, 10.1139/z90-058. [ Links ]
Méndez-Cárdenas, María Guadalupe et al. 2008 Geographic Variation in Loud Calls of Sportive Lemurs (Lepilemur ssp.) and their Implications for Conservation. American Journal of Primatology (70): 1-11. [ Links ]
Méndez-Cárdenas, María Guadalupe y Elke Zimmermann 2009 Duetting-A Mechanism to Strengthen Pair Bonds in a Dispersed Pair-Living Primate (Lepilemur edwardsi)? Am. J. Phys. Anthropol (139): 523-532. [ Links ]
Méndez-Cárdenas María Guadalupe et al. (En prep.). Seasonality and Paternity in a Population of Milne-Edwards' Sportive Lemur (Lepilemur edwardsi) in NWMadagascar.
Mitani, John 1984 The Behavioral Regulation of Monogamy in Gibbons (Hylobates muelleri). Behavior Ecol Sociobiol (15): 225-229. [ Links ]
---------- 1987 Territoriality and Monogamy among Agile Gibbons (Hylobates agilis). Behavior Ecol Sociobiol (20): 265-269. [ Links ]
Morris, Charles 1958 [1938] Fundamentos de la teoría de los signos. UNAM. México. [ Links ]
Müller, Alexandra y Urs Thalmann 2000 Origin and Evolution of Primate Social Organisation: a Reconstruction. Biology Review (75): 405-435. [ Links ]
Müller, Alexandra y Gustl Anzenberger 2002 Duetting in the Titi Monkey Callicebus Cupreus: Structure, Pair Specificity and Development of Duets. Folia Primatol (73): 104-115. [ Links ]
Naguib, Marc y Haven Wiley 2001 Estimating the Distance to a Source of Sound: Mechanisms and Adaptations for Long-Range Communication. Animal Behavior (62): 825-837. [ Links ]
Nietsch, Alexandra 1999 Duet Vocalizations among Different Populations of Sulawesi Tarsiers. Int J Primatol (20): 567-583. [ Links ]
---------- 2003 Outline of the Vocal Behaviour of Tarsius Spectrum: Call Features Associated Behaviours, and Biological Functions, en Tarsiers: Past, Present, and Future. P. C. Wright, El Simons y S. Gursky (eds.). Rutgers University Press. Nueva Brunswick: 196-220. [ Links ]
Ouattara, Karim, Alban Lemasson y Klaus Zuberbüler 2009 Campbell's Monkeys ConcatenateVvocalizations Into Context-Specific Call Sequences. PNAS (106): 22026-22031. [ Links ]
Owren, Michael y Drew Rendall 1997 An Affect-Conditioning Model of Nonhuman Primate Vocal Signaling, en Perspectives in Ethology: Communication, vol. 12, D. H. Owings, M. D. Beecher y N.S. Thompson (eds.). Plenum Press. Nueva York: 299-346. [ Links ]
Palombit, Ryne 1999 Infanticide and the Evolution of Pair Bonds in Nonhuman Primates. Evol Anthro (7): 117-129. [ Links ]
Peake, T. 2005 Eavesdropping in Communication Networks, en Animal Communication Networks, P. K. McGregor (ed.). Cambridge University Press. Cambridge: 13-37. [ Links ]
Peirce, Charles Sanders 1998 The Essential Peirce. Selected Philosophical Writings (1893-1913), 2. The Peirce Edition Project-Indiana University Press. Bloomington e Indianapolis. [ Links ]
Pollock, Jon I. 1975 Field Observations on Indri Indri: A Preliminary Report, en Lemur Biology, I. Tattersall y R. Sussman (eds.). Plenum. Nueva York: 28-31l. [ Links ]
---------- 1986 The Song in the Indris (Indri Indri; Primates: Lemuroidea): Natural History, form and Function. Int J Primatol (7): 225-264. [ Links ]
Rasmussen, Dennis 1981 Pair-Bond Strength and Stability and Reproductive Success. Psychol Rev (88): 274-290. [ Links ]
Rasoloharijaona, Solofonirina et al. 2003 Pairspecific Usage of Sleeping Sites and their Implications for Social Organization in a Nocturnal Malagasy Primate, the Milne Edwards' Sportive Lemur (Lepilemur edwardsi). Am J Phys Anthropol (122): 251-258. [ Links ]
---------- 2006 Loud Calling, Spacing, and Cohesiveness in a Nocturnal Primate, the Milne Edwards' Sportive Lemur (Lepilemur edwardsi). Am J Phys Anthropol (129): 591-600. [ Links ]
Riba, Carles 1990 La comunicación animal: un enfoque zoo semiótico. Anthropos. Barcelona: 463 pp. [ Links ]
Robinson, John 1979 Vocal Regulation of Use of Space by Groups of Titi Monkeys Callicebus moloch. Behavior Ecol Sociobiol (5): 1-15. [ Links ]
Robinson, John, Patricia Wright y Warren Kinzey 1987 Monogamous Cebids and their Relatives: Intergroup Calls and Spacing, en Primate Societies, B. B. Smuts et al. University of Chicago Press. Chicago: 44-53. [ Links ]
Saussure, Ferdinand 1986 Curso de lingüística general. Alianza Editorial. Madrid. [ Links ]
Savage-Rumbaugh, E. Sue et al. 1993 Language Comprehension in Ape and Child. Monogr. Soc. Res. Child Dev (58). [ Links ]
Schaik, Carel van y Peter Kappeler 2003 The Evolution of Social Monogamy in Primates, en Monogamy: Mating Strategies and Partnerships in Birds, Humans and Other Mammals, U. H. Reichard, C. Boesch (eds.). Cambridge University Press. Cambridge: 59-80. [ Links ]
Schaik, Carel van y Robin Dunbar 1990 The Evolution of Monogamy in Large Primates: a New Hypothesis and Some Crucial Tests. Behaviour (115): 30-62. [ Links ]
Scheumann, Marina, Andratahina Rabesandratana y Elke Zimmermann 2007 Predation, Communication and Cognition in Lemurs, en Primate Antipredator Strategies, S. L. Gursky, Kai Nekaris (eds.). Springer. Nueva York: 100-126. [ Links ]
Schön, Miguel 1971 The Anatomy of the Resonating Mechanism in Howling Monkeys. Folia Primatologica (15): 117-132. [ Links ]
Schülke, Oliver, Peter Kappler y Hans Zischler 2004 Small Testes Size Despite High Extra-Pair Paternity in the Pair-Living Nocturnal Primate Phaner furcifer. Behavior Ecol Sociobiology (55): 293-301. [ Links ]
Shultz, Sussane, Christopher Opie y Quentin Atkinson 2011 Stepwise Evolution of Stable Sociality in Primates. Nature (479): 219-222. [ Links ]
Smith, John 1977 The Behavior of Communicating (An Ethological Approach). Harvard University Press. Londres. [ Links ]
Snowdon, Charles 1997 Part I Persectives in New World Primates: Is Speech Special? Lessons for New World Primates, en New World Primates: Ecology, Evolution, and Behavior, W. G. Kinzey. Walter de Gruyter. Nueva York. [ Links ]
Sonnenschein, Edith y Heinz-Ulrich Reyer 1983 Mate-Guarding and Other Functions of Antiphonal Duets in the Slate-Coloured Boubou (Laniarius funebris). Z Tierpsychol (63): 112-140. [ Links ]
Stevenson, Pablo y María Clara Castellanos 2000 Feeding Rates and Daily Path Range of the Colombian Woolly Monkeys as Evidence for Between and within Group Competition. Folia Primatol (71): 399-408. [ Links ]
Stockley, Paula y Anne Campbell 2013 Female Competition and Aggression: Interdisciplinary Perspectives. Phil Trans R Soc, B 368: 20130073. <http://dx.doi.org/10.1098/rstb.2013.0073> [ Links ].
Stokes, Allen y Williams Warrington 1968 Antiphonal Calling in Quail. Auk (85): 83-89. [ Links ]
Stumpf, Rebecca M. et al. 2011 Sexual Conflict in Primates. Evolutionary Anthropology (20): 62-75. [ Links ]
Thalmann, Urs 1998 Sociality in the Nocturnal Lemurs Avahi and Lepilemur: A Comparison, en Abstract 064, XVII, Congress of the International Primatological Society, University of Antananarivo, Madagascar. [ Links ]
---------- 2001 Food Resource Characteristics in Two Nocturnal Lemurs with Different Social Behavior: Avahi Occidentalis and Lepilemur edwardsi. Int J Primatol (22): 287-324. [ Links ]
Thorpe, William Homan 1972 Duetting and Antiphonal Song in Birds. Behaviour, supl. (18): 1-193. [ Links ]
Tilson, Ronald y Richard Tenaza 1976 Monogamy and Duetting in an Old World Monkey. Nature (263): 320-321. [ Links ]
Tobías, Martha, Sandya Viswanathan y Darcy Kelley 1998 Rapping a Female Receptive Call, Initiates Malefemale Duets in the South African Clawed Frog. Proc Nat Acad Sci (95): 1870-1875. [ Links ]
Trivers, Robert 1972 Parental Investment and Sexual Selection, en Sexual Selection and the Descent of Man, B. Campbell (ed.). Aldine. Chicago: 136-179. [ Links ]
---------- 1985 Social Evolution, Benjamin Cummings. Menlo Park, California. [ Links ]
Tubaro, Pablo, Enrique Segura y Paul Handford 1993 Geographic Variation in the Song of the Rufouscollard Sparrow in Eastern Argentina. Cóndor (95): 588-595. [ Links ]
Wickler, Wolfgang 1976 Evolution-Oriented Ethology, Kin Selection, and Altruistic Parasites. Z Tierpsychol (42): 206-214. [ Links ]
---------- 1980 Vocal Duetting and the Pair Bond. I. Coyness and Partner Commitment. A Hypothesis. Z Tierpsychol (52): 201-209. [ Links ]
Wickler, Wolfgang y Uta Seibt 1982 Song Splitting in the Evolution of Duetting. Z Tierpsychol (59): 127-140. [ Links ]
Wiley, Haven 1991 Associations of Song Properties with Habitats for Territorial Oscine Birds of Eastern North America. Am Nat (138): 973-993. [ Links ]
Wiley, Haven y Douglas Richards 1978 Physical Constraints on Acoustic Communication in the Atmosphere: Implications for the Evolution of Animal Vocalizations. Behavior Ecol Sociobiology (3): 69-94. [ Links ]
While, Geoffrey, David Sinn y Erik Wapstra 2009 Female Aggression Predicts Mode of Paternity Acquisition in a Social Lizard. Proc. R. Soc. B 276: 2021-2029. <doi:10.1098/rspb.2008.1926> [ Links ].
Zahavi, Amotz 1975 Mate Selection: a Selection for a Handicap. J. Theoretical Biology (53): 205-214. [ Links ]
Zimmermann, Elke, John Newman y Uwe Jürgens 1995 Current Topics in Primate Vocal Communication. Plenum Press. Nueva York. [ Links ]
1 Reproducción de la grabación de una señal acústica de los monos realizada por los investigadores con el fin de analizar sus respuestas.
2 Análisis no paramétrico de la varianza en medidas repetidas.
3 Prueba no paramétrica que compara entre dos muestras o variables dependientes.
4 Prueba no paramétrica que compara entre dos muestras o variables independientes.

