










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCuicuilco
versión impresa ISSN 0185-1659
Cuicuilco vol.20 no.58 México sep./dic. 2013
Dossier: antropología genética
Análisis del DNA mitocondrial antiguo y contemporáneo: un acercamiento a las relaciones genéticas en las poblaciones indígenas de Mesoamérica
Angélica González Oliver,1 Ernesto Garfias Morales,1 Elizabeth Romero García,1 María Isabel de la Cruz Laina,1 Alín Patricia Acuña Alonzo,1 Mauricio Pérez Martínez,1 Fernando Sánchez Solís,1 Benjamín Cristian Corona Comunidad,1 David Glenn Smith2 y Alfonso Torre Blanco1
1 Departamento de Biología, Facultad de Ciencias, Universidad Nacional Autónoma de México.
2 Departamento de Antropología, Universidad de California, Davis.
Resumen
En el presente estudio analizamos los haplogrupos A, B, C y D del ácido desoxirribonucleico mitocondrial (mtDNA) en 108 individuos contemporáneos mazahuas y 68 otomíes del Estado de México. El objetivo del análisis es identificar las relaciones genéticas entre estos grupos y compararlos con otras poblaciones antiguas y contemporáneas de México. Los grupos poblacionales mazahua y otomí habitan en los mismos municipios del Estado de México, hablan lenguajes que pertenecen a la familia lingüística oto-mangue, comparten aspectos culturales y una historia en común. Los resultados mostraron que el haplogrupo B es el más frecuente en los mazahuas y el A en los otomíes. El haplogrupo C presenta frecuencias similares en ambos grupos. La población otomí presenta una baja frecuencia del haplogrupo D e incluye individuos que no presentaron ninguno de los cuatro haplogrupos del mtDNA, mientras que todos los individuos de la población mazahua pertenecieron a uno de los cuatro haplogrupos estudiados. Las poblaciones mazahua y otomí del actual Estado de México son estadísticamente diferentes por el método de ji cuadrada (p ≤ 0.05). El análisis de componentes principales basado en las frecuencias de los cuatro haplogrupos del mtDNA sugiere que entre ambos grupos poblacionales no ha habido flujo génico por vía materna o ha sido muy escaso.
Palabras clave: DNA mitocondrial, haplogrupos fundadores del DNA mitocondrial, otomíes, mazahuas, nahuas, poblaciones mexicanas.
Abstract
In this study we analyzed the founding mitochondrial DNA (mtDNA) haplogroups A, B, C and D in 108 contemporary Mazahua individuals and 73 Otomi from Estado de Mexico to understand the genetic relationship between these populations, and with other ancient and contemporary Mexican populations. The Mazahua and Otomi inhabit the same localities in Estado de México, speak languages that belong to the Oto-Manguean linguistic family and show cultural and historic similarities. Our results showed that haplogroup B is the most frequent in Mazahua whereas haplogroup A is highest among the Otomi. Haplogroup C exhibit similar frequencies. Otomi population exhibits a low frequency of haplogroup D and includes individuals that do not belong to any of the four mtDNA haplogroups, whereas all Mazahua individuals in our study belong to one of the four mtDNA lineages studied. The Mazahua and Otomi populations from Mexico State are statistically different according the results of the chi square test (p ≤ 0.05). The principal component analysis using the frequencies of four mtDNA haplogroups suggests that Mazahua and Otomi had little or no maternal genetic flow.
Keywords: mitochondrial DNA, founding Amerindian mitochondrial DNA haplogroups, Otomi, Mazahua, Nahua, Mexican populations.
Tratar de inferir las relaciones genéticas entre las poblaciones indígenas de México utilizando la información cultural y lingüística disponible llevaría a resultados inexactos y poco precisos. El lenguaje primario y las normas culturales se aprenden en la relación estrecha de los hijos con los padres biológicos y con otros miembros del núcleo familiar, lo que muchas veces implica el paso de conocimientos entre individuos no relacionados genéticamente. En cambio, la información genética se transmite directa y exclusivamente de padres a hijos en la concepción.
El DNA mitocondrial (mtDNA) se hereda exclusivamente de la madre a los hijos, tanto varones como mujeres, sin sufrir recombinación genética; lo cual permite la determinación de divergencias filogenéticas sin las ambigüedades causadas por el entrecruzamiento meiótico.
Los estudios de la variación del mtDNA han mostrado que la mayoría de los nativos americanos pertenecen a uno de los cinco haplogrupos maternos fundadores llamados A, B, C, D y X [Schurr et al., 1990; Torroni et al., 1992; Smith et al., 1999; Brown et al., 1998]. Los linajes o haplogrupos A, C, D y X se definen por mutaciones puntuales en posiciones específicas de la secuencia del mtDNA que originan la presencia o ausencia de un sitio de reconocimiento para una endonucleasa de restricción. Mientras que el haplogrupo B se define por la ausencia de nueve pares de bases en la región V del genoma mitocondrial.
Por otra parte, el análisis de la variación genética de la región control del mtDNA ha permitido identificar sitios polimórficos que comparten los individuos de un mismo haplogrupo fundador, los cuales definen haplotipos (sublinajes) específicos en cada linaje. Los sublinajes pueden ser específicos de grupos poblacionales [Malhi et al., 2002; Torroni et al., 1993] o de familias lingüísticas [Derbeneva et al., 2002], por lo que se utilizan para estudiar con mayor precisión las relaciones genéticas entre las poblaciones.
Los estudios de los haplogrupos fundadores del mtDNA y de la variación genética en la región hipervariable de la región control del genoma mitocondrial han sido ampliamente usados para estudiar las relaciones de ancestro-descendencia en las poblaciones humanas, el poblamiento del continente americano, el número de migraciones realizadas, las fechas en que ocurrieron y rutas de migración de las poblaciones, entre otras [Schurr et al., 1990; Torroni et al., 1992, 1993; Tamm et al., 2007].
El descubrimiento de la presencia de ácido desoxirribonucleico en restos óseos, tejidos momificados, dientes y coprolitos humanos, y el desarrollo de técnicas moleculares para su recuperación, han permitido estudiar de manera directa la genética en las poblaciones antiguas [Kemp et al., 2010; De la Cruz et al., 2008; González-Oliver et al., 2001; Pääbo, 1985; Habelberg et al., 1989].
Mesoamérica
Los antropólogos y lingüistas han propuesto conexiones culturales y lingüísticas en las poblaciones indígenas antiguas de origen mesoamericano. Las poblaciones mesoamericanas practicaban el juego de pelota, presentaban estilos parecidos en cerámica, adornos personales, arquitectura, técnicas textiles y métodos de agricultura, siendo la del maíz la más importante, con su origen en el centro de México [Cordell, 1997; Haury, 1974; Hill, 2001 y 2002; Smith, 1994 y1995].
La región de Mesoamérica incluía una gran diversidad de grupos poblacionales que hablaban distintos lenguajes, la mayoría de éstos se localizaban en México. Existen diferentes clasificaciones lingüísticas de las poblaciones indígenas mesoamericanas [Ortiz, 2005], sin embargo, en los estudios de antropología molecular frecuentemente se utiliza la clasificación lingüística propuesta por [Campbell et al., 1986] para los grupos de Norte y Centroamérica, basada en las similitudes lingüísticas compartidas entre los lenguajes de esta amplia área geográfica. Los rasgos lingüísticos similares que comparten estos lenguajes han sido explicados por el contexto histórico o por la herencia de un proto-lenguaje. Actualmente las familias lingüísticas presentes en México son oto-mangue, yuto-azteca, totonaca, mixe-zoque y maya [Campbell et al., 1986]. La familia Yuto-azteca es considerada una de las principales familias en el mundo. Las poblaciones que hablan algún lenguaje miembro de esta familia se encuentran distribuidas desde el sur de Idaho, Estados Unidos, hasta Panamá [Miller, 1983]. La familia oto-mangue y maya están presentes desde San Luis Potosí y la Península de Yucatán, respectivamente, hasta Centroamérica.
Existen pocos estudios bioantropológicos realizados en las poblaciones antiguas de la región mesoamericana y en poblaciones indígenas modernas de México. La presencia de flujo génico entre las poblaciones indígenas mexicanas se infiere de los estudios del sistema sanguíneo ABO [Lagunas y López, 2004], los análisis del marcador nuclear Albúmina México, ampliamente distribuido en las poblaciones contemporáneas mexicanas [Smith et al., 2000], y los estudios de los linajes fundadores Q y C del cromosoma Y y A, B, C, D y X del mtDNA en grupos indígenas contemporáneos y antiguos, los cuales muestran la presencia predominante del linaje Q [Malhi et al., 2008; Zegura et al., 2004] y A, respectivamente [Kemp et al., 2010; Sandoval et al., 2009; Malhi et al., 2003; Schurr et al., 1990; Torroni et al., 1994; González-Oliver et al., 2001].
Actualmente la evidencia genética basada en el análisis del mtDNA no sugiere una relación simple entre genes mitocondriales, lenguaje y cultura en las poblaciones indígenas mexicanas [Kemp et al., 2010; Sandoval et al., 2009].
Registro histórico de los primeros grupos en el centro de México
Los grupos poblacionales otomí, mazahua, matlatzinca y nahua han compartido el mismo territorio desde la época prehispánica. Mientras que la llegada de los aztecas al centro de México se menciona en diferentes fuentes históricas y códices, no contamos con información comparable sobre la llegada de los otomíes y los mazahuas a esta misma región.
Después de la caída del Imperio tolteca (900-1170 d. C.), la población que habitaba la ciudad de Tula se dispersó, brindando la oportunidad para que otros grupos ocuparan el centro de México. Uno de esos grupos fue el de los mexicas, quienes posteriormente fundaron el Imperio azteca. Los códices refieren el origen de los aztecas en una tierra al norte llamada 8 Aztlán [Townsend, 1992].
Durante su migración al centro de México, los mexicas se refugiaron en Tizapán, que era un pueblo de refugiados toltecas que huían de Tula. Tizapán estaba cerca de Culhuacán, por lo que los mexicas se mezclaron con los culhuas y se unieron a ellos para pelear contra sus vecinos de Xochimilco. Tiempo después los mexicas fueron expulsados de Culhuacán [Townsend, 1992]. La población mexica encontró deshabitado uno de los islotes del Lago de Texcoco, y ahí fundó Tenochtitlán, aproximadamente en el año 1325 o 1345; también fundaron Tlatelolco en 1337 o quizá antes, iniciándose el nacimiento del gran Imperio azteca [Matos, 1989].
De acuerdo con Fray Toribio de Benavente-Motolinía [1989], los grupos "otomianos" son descendientes de chichimecas.
Los Anales de Cuauhtitlán mencionan que cinco tribus llegaron al altiplano central aproximadamente en el 538 d. C. en una migración. Entre los dirigentes de las tribus estaba Mazacóhuatl, quien era el jefe de los mazahuas. En una peregrinación posterior, en el siglo X, los chichimecas acaudillados por Mixcóatl fundaron Culhuacán y Tula [González y Gutiérrez, 1999]. También se ha propuesto que el pueblo más antiguo que habitó la planicie es el de los otomíes, procedentes del oriente o del sur del Golfo de México, y se les considera pertenecientes a los llamados olmecas [Serrano, 1999; Nolasco, 1999; Huitrón, 1999].
En relación con el origen del pueblo mazahua, se propuso que descienden del grupo otomí [Pérez, 1999; Morón, 1999 y Clavijero, 2003]. Los mazahuas fueron invadidos repetidas veces por teotihuacanos, toltecas, chichimecas, tecpanecas, aztecas y finalmente por los españoles [Garduño, 1999].
Después de la llegada de los otomíes y mazahuas al actual estado de México, la región fue dominada por los mexicas, quienes moraron ahí antes de la fundación de Tenochtitlán [Nolasco, 1999].
La presencia de los mexicas o tenochcas en la región otomí se infiere de la cerámica y escultura encontrada en el sitio arqueológico de Huamango, que fue el principal centro ceremonial y comercial de la región otomí.
Los grupos mazahuas tuvieron como centro de gobierno a Nguemore, que hoy es Jocotitlán en el Estado de México; ahí se recibían los tributos para los aztecas. A su vez, la región mazahua en Jocotitlán formaba parte del reino otomí de Xaltocan [Garduño, 1999].
Finalmente los mazahuas y otomíes se unieron a los mexicas para defenderse de los españoles [Garduño, 1999]. Después de la derrota, la población indígena fue reducida dramáticamente debido a enfermedades como la viruela, el exceso de trabajo y la violencia. Aunque los españoles reorganizaron a la población indígena en la tierra recién conquistada, algunas veces los asentamientos de la gente indígena incluían poblaciones no relacionadas para conformar una nueva comunidad [Gibson, 1964].
Situación actual de las poblaciones contemporáneas mazahua, otomí y nahua
Población mazahua. El grupo mazahua se extiende en la parte noroeste del Estado de México. También comprende una pequeña área del oriente del Estado de Michoacán correspondiente al municipio de Zitácuaro [Scheffler, 1992] y en el Estado de México comparte territorio con otomíes.
El censo del Instituto Nacional de Información Estadística Geografía e Informática (INEGI) del año 2005 registró 228 568 individuos mazahuas en el Estado de México. La población total de mazahuas en México es de 268 216 individuos [INEGI, 2005] y la principal actividad económica es la agricultura, igual que para los otomíes [INI, 1982].
Población otomí. Los otomíes se distribuyen en una extensa región del noroeste del Estado de México. Además, habitan en los estados de Hidalgo, Puebla, Tlaxcala, Veracruz, Guanajuato y Querétaro [Lagunas y López, 2004].
Existen 209 714 individuos otomíes en el Estado de México. La población total otomí es de 494 480 individuos [INEGI, 2005] y las lenguas otomí y mazahua pertenecen a la familia lingüística oto-mangue [Campbell et al., 1986].
Población nahua. El grupo nahua es el más numeroso de México. Se encuentran distribuidos en 12 estados. Muchos nahuas están viviendo en comunidades rurales cercanas a la Ciudad de México, a la cual migran temporalmente para trabajar [Scheffler, 1992]. El lenguaje nahua es miembro de la familia lingüística yuto-azteca y fue el principal lenguaje durante el Imperio azteca.
Existen 1 448 936 nahuas en México. En el Distrito Federal hay 37 450 hablantes de nahua [INEGI, 2005].
En el presente estudio analizamos los haplogrupos o linajes fundadores del mtDNA en las poblaciones contemporáneas mazahua y otomí del centro de México y las comparamos con otras poblaciones antiguas y modernas de la misma región. El objetivo de esto es tratar de entender las relaciones genéticas entre los primeros pobladores del Altiplano Central que pertenecen a las familias lingüísticas oto-mangue y yuto-azteca.
Materiales y métodos
Poblaciones estudiadas
Se colectaron muestras de frotis bucal de 68 otomíes y 108 mazahuas del Estado de México. Los individuos no son familiares y habitan en los municipios de Ixtlahuaca, Jocotitlán, San Felipe de Progreso, Temascalcingo, Almoloya de Juárez, Atlacomulco y Aculco.
Se obtuvo el consentimiento escrito, informado y voluntario de los individuos para participar en el proyecto, el cual garantiza la privacidad de los participantes conforme a las normas internacionales de los datos genéticos humanos, las cuales fueron dadas a conocer en forma clara y expresa a cada individuo así como a las autoridades municipales, civiles y religiosas de las comunidades visitadas.
Extracción del DNA y amplificación por la reacción en cadena de la polimerasa (PCR)
El DNA fue extraído de los frotis bucales con el mini kit comercial QIAamp DNA Blood de Qiagen. Se amplificaron segmentos específicos del DNA mitocondrial que contienen los sitios de reconocimiento para los haplogrupos A, B, C y D. En estas amplificaciones se utilizaron primers, que se han utilizado anteriormente para analizar estos mismos marcadores [González Oliver et al., 2011 y 2001].
Los productos de PCR de los haplogrupos del mtDNA fueron analizados en geles de poliacrilamida al 12 % (linajes A, C y D) o 18 % (linaje B) en el amortiguador de Tris-Borato-EDTA [Sambrook, Fritsch y Maniatis, 1989]. Posteriormente los geles se tiñeron con bromuro de etidio y se determinó el tamaño de los productos por comparación con el marcador phi- X174 cortado con Hae III (Gibco BRL).
Análisis de restricción
Los productos de PCR de los haplogrupos A, C y D del mtDNA fueron incubados a 37° C toda la noche con cinco unidades de la endonucleasa de restricción específica para cada haplogrupo (Hae III, Hinc II y Alu I). El producto de la digestión se analizó en un gel de poliacrilamida al 12 % como se describió arriba.
Se compararon las frecuencias de los haplogrupos fundadores del mtDNA entre las dos poblaciones estudiadas aquí y con otras citadas en la literatura utilizando la prueba estadística de Ji cuadrada.
Se realizó un análisis de componentes principales (ACP) con el complemento estadístico XLSTAT, versión 2011, del programa Microsoft Office Excel 2010 [Addinsoft, 2011]. Se utilizaron las frecuencias normalizadas de los cuatro haplogrupos fundadores del mtDNA de las poblaciones de México.
Cabe aclarar que en el análisis estadístico los individuos no pertenecientes a los haplogrupos A, B, C y D del mtDNA fueron excluidos, ya que pueden pertenecer al haplogrupo X o representar mezcla no nativa americana. Algunas poblaciones citadas en la bibliografía no se utilizaron en el presente estudio debido a que los mismos individuos indígenas fueron analizados por diferentes grupos de investigación, por ejemplo Kemp et al. [2010], Sandoval et al. [2009] y Peñaloza Espinosa et al. [2007]; (Rosenda Peñaloza Espinosa, comunicación personal, 2013).
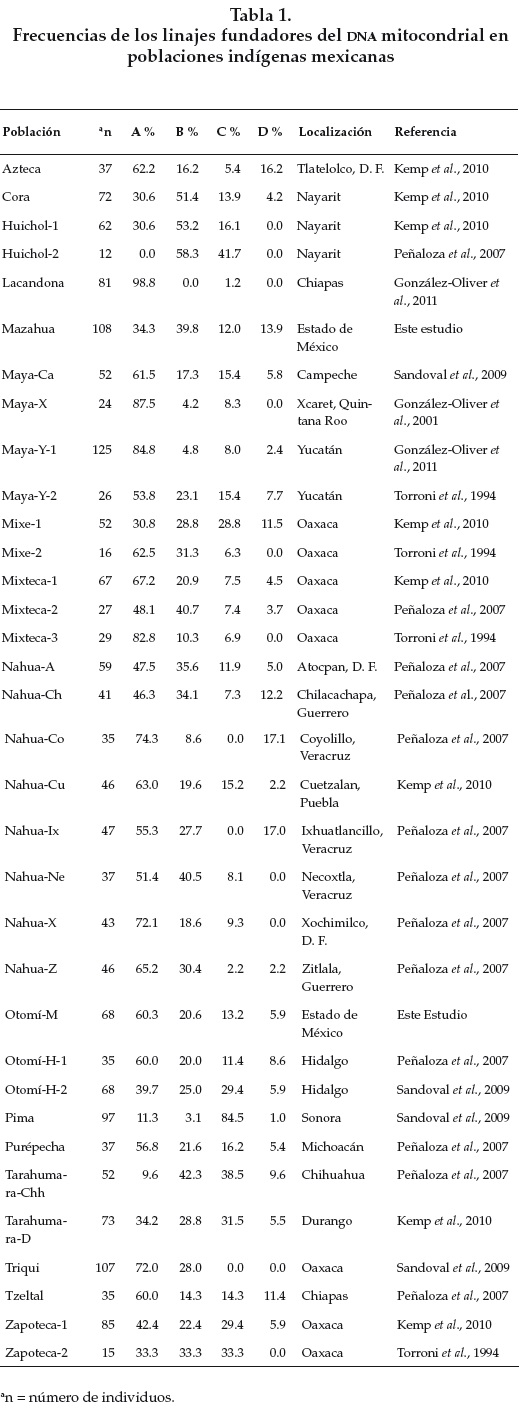
Para referirnos a los grupos poblacionales del mismo estado o localidad usaremos la nomenclatura que aparece en la tabla 1.
Resultados
Las poblaciones mazahua y otomí del Estado de México muestran la presencia de los cuatro linajes o haplogrupos fundadores del mtDNA.
En la población otomí el haplogrupo A es el más frecuente (60 %), el D es el menos frecuente (6 °%) y los haplogrupos B y C presentan frecuencias intermedias de 21 y 13 % respectivamente.
En la población mazahua el haplogrupo B es el más frecuente (40 %%), seguido del A (34 %) y frecuencias intermedias de los haplogrupos C y D (12 y 14 %% respectivamente).
Hay individuos otomíes que no mostraron ninguno de los cuatro haplogrupos del mtDNA (4 %%), por lo que se excluyeron del análisis estadístico. Posteriormente se analizarán para saber si pertenecen al haplogrupo X. Otras seis poblaciones mexicanas citadas en la bibliografía: lacandona de Chiapas, maya de Yucatán-2, maya antigua de Xcaret, nahua de Coyolillo, Veracruz, pima de Sonora y zapoteca de Oaxaca-1 también presentan individuos que no pertenecen a ninguno de los haplogrupos A, B, C y D; estos individuos también fueron excluidos del análisis.
La tabla 1 y la figura 1 muestran la ubicación geográfica y las frecuencias de los haplogrupos fundadores del mtDNA de las poblaciones antiguas y contemporáneas con las que fueron comparados los resultados de este estudio.
Relación genética de los otomíes y mazahuas con otros grupos poblacionales de México
El análisis de las frecuencias de los cuatro linajes fundadores del mtDNA mostró que el linaje A es el más frecuente en las poblaciones contemporáneas y antiguas de México, excepto en las poblaciones mazahua, cora de Nayarit, tarahumara de Chihuahua y las dos poblaciones huicholes de Nayarit. El linaje es el segundo más frecuente en las poblaciones mexicanas. Los linajes C y D son poco frecuentes, excepto en los pima de Sonora (84 % del linaje C), siendo el linaje D el menos frecuente de los dos. El linaje X solamente se ha detectado en las poblaciones huichol-2 y tarahumara de Chihuahua.
Las poblaciones mazahua y otomí del actual Estado de México son estadísticamente diferentes entre sí por el método de ji cuadrada (p ≤ 0.05). El grupo mazahua no es estadísticamente diferente de las poblaciones cora (p = 0.128), mixe-1 (p = 0.068), mixteca-2 (p = 0.329), nahua A (p = 0.087), nahua Ch (p = 0.555), nahua Ix (p = 0.052), maya Y-2 (p = 0.209), mixe-2 (p = 0.120) y zapoteca-2 (p = 0.097).
Mientras que la población otomí no muestra diferencias estadísticas con: azteca (p = 0.226); maya de Campeche (p = 0.965), maya-Y-2 (p = 0.951) y de Xcaret (p = 0.083); mixe-2 (p = 0.545); mixteca-1 (p = 0.690), mixteca-2 (p = 0.238), mixteca-3 (p = 0.159); nahua A (p = 0.086), nahua Ch (p = 0.188), nahua Cu (p = 0.804), nahua X (p = 0.088), nahua Z (p = 0.095); otomi-H-2 (p = 0.061), otomi-H-1 (p = 0.958); purépecha (p = 0.974); tzeltal (p = 0.698); zapoteca-2 (p = 0.098), zapoteca-1 (p = 0.074) y es estadísticamente diferente del resto de las poblaciones mexicanas.
Ambos grupos no son estadísticamente diferentes de las poblaciones: nahua Ch, nahua Ix, mixteca-2, maya-Y-2, mixe-2, zapoteca-2 y nahua A. La figura 2 muestra la gráfica del ACP en dos dimensiones; 51.08 % de la variación corresponde al factor 1 (eje de las X) y 26.65 % al factor 2 (eje de las Y). Se observa que las poblaciones mazahua y otomí analizadas en este estudio están separadas, sugiriendo un escaso o ausente flujo génico por vía materna entre ambos grupos a pesar de que: a) viven en los mismos municipios del Estado de México y han ocupado este territorio desde la época prehispánica, b) comparten aspectos culturales y filiación lingüística y c) tienen una historia regional en común. Muy probablemente las costumbres matrimoniales propias de cada población y las diferencias entre ambos grupos a lo largo de su historia los han mantenido separados a través del tiempo.
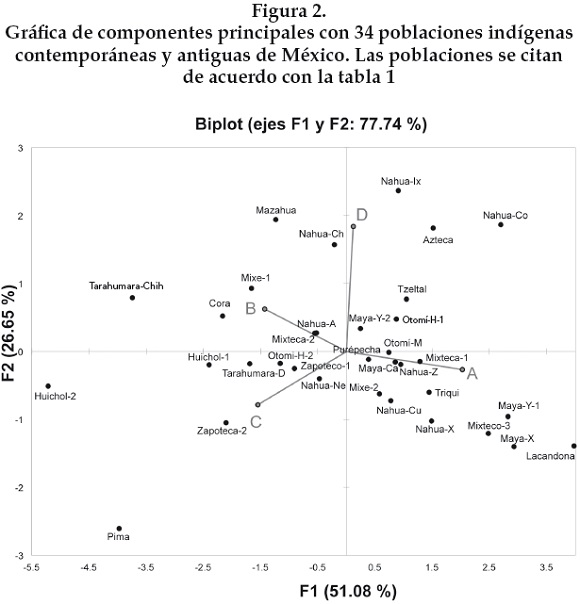
La población mazahua también se observa separada de las poblaciones otomíes de Hidalgo-1 y 2 en el ACP aunque se encuentra genéticamente más cercana a la población otomí de Hidalgo-2. Por otra parte, las tres poblaciones otomíes, la del Estado de México y las dos de Hidalgo, no son genéticamente cercanas entre sí, aunque los otomíes del Estado de México y de Hidalgo-1 son genéticamente más cercanos entre sí. Cabe resaltar que los dos grupos otomíes de Hidalgo-1 y 2 no están genéticamente cercanos entre sí. Los autores no mencionan a qué localidades o municipios pertenecen las muestras.
La población mazahua estudiada aquí está genéticamente relacionada con la población nahua de Chilacachapa, y muestra mayor cercanía genética con la nahua de Atocpan que con el resto de las poblaciones nahuas. Mientras que el grupo otomí está relacionado genéticamente con la nahua de Zitlala y de Cuetzalan.
Los aztecas de Tlatelolco del periodo prehispánico son genéticamente más cercanos a los grupos nahua de Coyolillo e Ixhuatlancillo que a los nahuas de Atocpan y Xochimilco. Los aztecas están distantes de las poblaciones otomíes y mazahua, aunque están más cercanos a los otomíes de Hidalgo-1 y del Estado de México.
Los nahuas de Atocpan están más cercanos a los nahuas de Necoxtla que a los nahuas de Xochimilco. Los nahuas de Zitlala y Xochimilco muestran cercanía génica entre sí en el ACP. Por otra parte, los nahuas de Coyolillo y de Ixhuatlancillo son los grupos más alejados de la mayoría de las poblaciones nahuas. En la población nahua de Coyolillo se identificó el haplogrupo L (8%) que representa la mezcla de origen africano [Peñaloza et al., 2007].
Los nahuas contemporáneos de Atocpan y de Xochimilco no están genéticamente relacionados entre sí, y tampoco con los aztecas antiguos de Tlatelolco, a pesar de que son localidades que se encuentran geográficamente próximas. A este respecto cabe mencionar que el lenguaje nahuátl fue el principal lenguaje hablado durante el Imperio azteca. Este lenguaje funcionaba como una lengua franca en el centro de México cuando inició el contacto español [Fagan, 1984]. Actualmente es el lenguaje más hablado en México, los hablantes de la lengua nahuátl representan 1.3% de la población actual de México [INEGI, 2005]. Muchos grupos indígenas antiguos hablaban nahuátl, lo cual no implica que haya relación genética con los aztecas.
Recientemente se propuso un patrón diferente en las frecuencias de los haplogrupos fundadores del mtDNA entre las poblaciones de las regiones culturales de Mesoamérica y el suroeste americano. La mayoría de las poblaciones del suroeste exhibe una alta frecuencia del haplogrupo B y una baja frecuencia o ausencia del haplogrupo A. Mientras que las altas frecuencias del haplogrupo A son caracteríscas de las poblaciones de Mesoamérica [Kemp et al., 2010]. En términos generales los grupos pima de Sonora, tarahumara de Chihuahua y huichol de Nayarit están ubicados lejos de la mayoría de las poblaciones de México en el ACP. Los huicholes están ubicados cerca del límite geográfico propuesto para la región cultural entre Mesoamérica y el suroeste americano, aunque se ha documentado que el límite superior de Mesoamérica ha variado a través del tiempo [Manzanilla y López, 1993]. Por otra parte, los grupos poblacionales pima y tarahumara se ubican en la región cultural del suroeste americano. Cabe mencionar que las poblaciones huichol-1 y cora están genéticamente relacionadas entre sí, y ambas son más cercanas a los tarahumaras-Chh que a la otra población huichol-2. Los coras también se ubican en el límite entre las dos regiones culturales. Tampoco las dos poblaciones tarahumaras de Chihuahua y de Durango muestran cercanía genética.
En relación con los grupos poblacionales del estado de Oaxaca no existe una relación genética entre los mixe-1 y 2. Los mixtecos-1, 2 y 3 están lejanamente relacionados entre sí; igual que los zapotecos-1 y 2. Cabe resaltar que la población mixteca-2 está estrechamente relacionada con la nahua de Atocpan. Las poblaciones del sureste que muestran la mayor cercanía génica son la zapoteca-1, la mixe-2 y la triqui. Para ayudar a entender diferencias regionales entre las poblaciones sería importante conocer las localidades o municipios donde se colectaron las muestras, pero en estos casos los autores no las reportaron.
Todas las poblaciones de origen maya, antiguas y modernas, de la península de Yucatán, están ubicadas cercanamente en el ACP, lo que apoya la continuidad genética entre las poblaciones mayas de las tierras bajas propuesta previamente en González-Oliver et al. [2001 y 2011]. Los grupos poblacionales mayas han experimentado diferentes realidades históricas pero comparten patrones culturales y lingüísticos que los agrupan en una unidad cultural. Los resultados del presente estudio indican que los mayas son el grupo cultural con mayor relación genética entre las poblaciones analizadas por diferentes grupos de investigación, a diferencia de los otros grupos culturales estudiados. La lengua maya es la segunda más hablada en México después del náhuatl.
En contraste con los grupos mayas, los resultados de las poblaciones nahuas contemporáneas y azteca antigua muestran que la gran mayoría de éstas no se agrupan cercanamente en el ACP, sugiriendo que no existe una relación genética continua por vía materna en el grupo cultural y lingüístico nahua.
Futuros estudios de la variación genética de la región hipervariable I del mtDNA y del cromosoma Y en las poblaciones modernas y antiguas de otomíes y mazahuas, y de otros grupos de la región cultural de Mesoamérica, ayudarán a entender con mayor precisión las relaciones genéticas entre los grupos de México.
Agradecimientos. Agradecemos la generosidad de todas las personas mazahuas y otomíes que voluntariamente participaron en el presente estudio. De forma especial al señor Crisanto Maya Miranda, jefe supremo mazahua de San Antonio de las Huertas en el municipio de San Felipe del Progreso e integrante del Consejo Estatal para el Desarrollo de los Pueblos Indígenas del Estado de México (CEDIPIEM). Al señor Arturo Martínez Hernández, jefe supremo mazahua del municipio de Villa Victoria. Al Centro Bachillerato Tecnológico Agropecuario (CBTA) núm. 128 de San Felipe del Progreso, principalmente al señor Hernán Hernández Martínez director del plantel. Al señor Benigno Becerril Sánchez, delegado municipal de Guadalupe Cachi, municipio de Ixtlahuaca, y al señor Macario Ventura Garduño de la Casa de la Cultura de Ixtlahuaca; al médico Alfredo Alegría Reyes, quien labora en la clínica de Guadalupe Cachi. Al CBTA núm. 150 del municipio de Acambay por su participación en la colecta de las muestras de personas otomíes, especialmente a la profesora Laura Hernández Herrera. Y al doctor Martín García Varela del Instituto de Biología de la Universidad Nacional Autónoma de México por su participación en la colecta de muestras mazahuas y otomíes.
Este estudio se realizó con el apoyo de UC-Mexus-Conacyt 2006, dentro de la convocatoria Proyectos de Investigación en Colaboración (Collaborative Research Grant); UC-Mexus 2008 convocatoria Small Grant Program from UC Mexus. Number: 08-003787; Conacyt, Investigación Científica Básica 2008, No. 101791; Proyecto PAPIIT IN229409-2, Universidad Nacional Autónoma de México y Proyecto SEP-Conacyt 83541.
Bibliografía
Addinsoft 2011 XLSTAT, Programa estadístico utilizado para análisis de datos y estadísticas para Microsoft Excel, versión 2011, www.xlstat.com. [ Links ]
Brown, Michael D., Seyed H. Hosseini et al. 1988 "mtDNA Haplogroup X: An Ancient Link Between Europe/Western Asia and North America?", American Journal Human Genetic, vol. 63, núm. 6, pp. 1852-1861. [ Links ]
Campbell, Lyle, Kaufman Terrence et al. 1986 "Meso-America as a linguistic área", Language, vol. 62, núm. 3, pp. 530-570. [ Links ]
Clavijero, Francisco J. 2003 Historia antigua de México, México, 10a edición, Editorial Porrúa, 879 pp. [ Links ]
Cordell, Linda S. 1997 Archaeology of the Southwest, 2a edición San Diego, Academic Press. [ Links ]
De la Cruz, Isabel, Angélica González-Oliver et al. 2008 "Sex Identification of Infants Sacrificed to the Ancient Aztec Rain Gods in Tlatelolco, Mexico", Current Anthropology, vol. 49, núm. 3, pp. 520-526. [ Links ]
Derbeneva, Olga. A., Rem I. Sukernik et al. 2002 "Analysis of Mitochondrial DNA Diversity in the Aleuts of the Commander Islands and its Implications for the Genetic History of Beringia", American Journal of human Genetics, vol. 71, núm. 2, pp. 415-421. [ Links ]
Fagan, Brian M. 1984 The Aztecs, Nueva York, W. H. Freeman, 322 pp. [ Links ]
Garduño Cervantes, Julio 1999 Temascalcingo, monografía municipal, Toluca, México, Instituto Mexiquense de Cultura-Asociación Mexiquense de Cronistas Municipales-Gobierno del Estado de México, 168 pp. [ Links ]
Gibson, Charles 1964 Aztecs under Spanish Rule: a History of the Indians of the Valley of Mexico, Stanford, Stanford University Press, pp. 1519-1810. [ Links ]
González Gómez, José Daniel y Pedro Gutiérrez Arzaluz 1999 Villa Victoria, monografía municipal, Toluca, México, Instituto Mexiquense de Cultura-Asociación Mexiquense de Cronistas Municipales-Gobierno del Estado de México, 132 pp. [ Links ]
González-Oliver, Angélica, Alín P. Acuña Alonzo et al. 2011 "Análisis genético de poblaciones mayas y lacandonas de las tierras bajas", en Memorias del IV Simposio Internacional el Hombre Temprano en América, México, IIA-UNAM, pp. 59- 74. [ Links ]
González-Oliver, Angélica, Lourdes Márquez-Morfin et al. 2001 "Founding Amerindian Mitochondrial DNA Lineages in Ancient Maya from Xcaret, Quintana Roo", American Journal of Physical Anthropology. vol. 116, pp. 230-235. [ Links ]
Hagelberg, Erica, Brian Sykes et al. 1989 "Ancient Bone DNA Amplified", Nature, vol. 342, núm. 6249, p. 485. [ Links ]
Haury, Emil W. 1974 "The Problem of Contacts Between the Southwestern United States and Mexico", en Basil Calvin Hedrich, The Mesoamerican Southwest: Reading in Archaeology, Ethnohistory and Ethnology, Illinois, Southern Illinois University Press, pp. 92-102. [ Links ]
Hill, Jane H. 2001 "Proto-Uto Aztecan: a Community of Cultivators in Central Mexico?", American Anthropologist, vol, 103, pp. 913-934. [ Links ]
---------- 2002 "The Origins of the Uto-Aztecans", ponencia presentada en el V. Coloquio Paul Kirchoff "Desiertos y fronteras, una lectura antropológica de la historia", México, Distrito Federal. [ Links ]
Huitrón Huitrón, Antonio 1999 Jilotepec, monografía municipal, Toluca, México, Instituto Mexiquense de Cultura-Asociación Mexiquense de Cronistas Municipales-Gobierno del Estado de México, 186 pp. [ Links ]
Instituto Nacional de Estadística Geografía e Informática (INEGI) 2005 II conteo de población y vivienda, México, INEGI. [ Links ]
Instituto Nacional Indigenista (INI) 1982 Grupos étnicos de México, México, INI. [ Links ]
Kemp, Brian M., Angélica González-Oliver et al. 2010 "Evaluating the Farming/Language Dispersal Hypothesis with Genetic Variation Exhbited by Populations in the Southwest and Mesoamerica", Proceedings National Academy of Sciences USA, vol. 107, núm. 15, pp. 6759-6764. [ Links ]
Lagunas Rodríguez, Z. y Sergio López Alonso 2004 "Antropología física en grupos humanos de filiación otopame", Ciencia Ergo Sum, vol. 1, pp. 47-58. [ Links ]
Malhi, Ripan S., Holly M. Mortensen et al. 2003 "Native American mtDNA Prehistory in the American Southwest", American Journal of Physical Anthropology, vol. 120, núm. 2, pp. 108-124. [ Links ]
Malhi, Ripan S., Jason A. Eshleman et al. 2002 "The Structure of Diversity within New World Mitochondrial DNA Haplogroups: Implications for the Prehistory of North America", American Journal Human Genetic, vol. 70, núm. 4, pp. 905-919. [ Links ]
Malhi, Ripan S., Angélica González-Oliver et al. 2008 "Y Chromosome Diversity among Native Americans: Investigating Athapaskan and Uto-Aztecan Populations Histories", American Journal Physical Anthropology, núm. 137, pp. 412-424. [ Links ]
Manzanilla, Linda y Leonardo López Luján 1993 Atlas histórico de Mesoamérica, México, Larousse, 201 pp. [ Links ]
Matos Moctezuma, E. 1989 The Aztecs, Nueva York, Rizzoli, 239 pp. [ Links ]
Miller Wick, R. 1983 "Uto-Aztecan Languages", en W. C. Sturtevant (ed.), Southwest, Handbook of North American Indians, vol. 10, Washington, D. C., Smithsonian Institution, pp. 113-124. [ Links ]
Morón Becerril, Carlos C. 1999 Villa de Allende, monografía municipal, Toluca, México, Instituto Mexiquense de Cultura-Asociación Mexiquense de Cronistas Municipales-Gobierno del Estado de México, 156 pp. [ Links ]
Motolinía, fray Toribio 1989 El libro perdido, México, Consejo Nacional para la Cultura y las Artes, 648 pp. [ Links ]
Nolasco Alvarez, Miguel A. 1999 Aculco, monografía municipal, Toluca, México, Instituto Mexiquense de Cultura-Asociación Mexiquense de Cronistas Municipales-Gobierno del Estado de México, 118 pp. [ Links ]
Ortiz Alvarez, María I. 2005 La población hablante de lenguas indígenas en México, México, IG-UNAM (Temas Selectos de Geografía de México, núm. 1.3.3.), 102 pp. [ Links ]
Pääbo, Svante 1985 "Molecular Cloning of Ancient Egyptian Mummy DNA", Nature, núm. 314, pp. 644-645. [ Links ]
Peñaloza Espinoza, Rosenda, Diego Arenas-Aranda et al. 2007 "Characterization of mtDNA Haplogroups in 14 Mexican Indigenous Populations", Human Biology, vol. 79, núm. 3, pp. 313-320. [ Links ]
Pérez Alvirde, Rosalío M. 1999 Zinacantepec, monografía municipal, Toluca, México, Instituto Mexiquense de Cultura-Asociación Mexiquense de Cronistas Municipales-Gobierno del Estado de México, 126 pp. [ Links ]
Sambrook, Joseph, Edward F. Fritsch et al. 1989 Molecular Cloning. A Laboratory Manual, Nueva York, Cold Spring HarborLaboratory Press, 3 vols. [ Links ]
Sandoval, Karla., Leonor Buentello-Malo et al. 2009 "Linguistic and Maternal Genetic Diversity are not Correlated in Native Mexicans", Human Genetics, vol. 126, núm. 4, pp. 521-531. [ Links ]
Serrano Pérez, Edgar A. 1999 Acambay, monografía municipal, Toluca, México, Instituto Mexiquense de Cultura-Asociación Mexiquense de Cronistas Municipales-Gobierno del Estado de México, 96 pp. [ Links ]
Scheffler, Lilian 1992 Los indígenas mexicanos, México, Panorama, 250 pp. [ Links ]
Schurr, Theodore G., Scott W. Ballinger et al. 1990 "Amerindian Mitochondrial DNAs have Rare Asian Mutations at High Frequencies, Suggesting they Derived from Four Primary Maternal Lineages", American Journal Human Genetic, vol. 46, núm. 3, pp. 613-623. [ Links ]
Smith, Bruce D. 1994 "The Origins of Agriculture in the Americas", Evolutionary Anthroplogy, vol. 3, núm. 5, pp. 174-184. [ Links ]
---------- 1995 The Emergence of Agriculture, Nueva York, Scientific American Library, 231 pp. [ Links ]
Smith, David G., Joseph G. Lorenz et al. 2000 "Implications of the Distribution of Albumin Naskapi and Albumin Mexico for New World Prehistory", American Journal of Physical Anthropology, vol. 111, pp. 557-572. [ Links ]
Smith, David G., Ripan S. Malhi et al. 1999 "Distribution of mtDNA Haplogroup X among Native North Americans", American Journal of Physical Anthropology, vol. 110, núm. 3, pp. 271-284. [ Links ]
Tamm, Erika, Toomas Kivisild et al. 2007 "Beringian Standstill and Spread of Native American Founders", Plos One, vol. 2, núm. 9, p. e829. [ Links ]
Torroni, Antonio., Theodore G. Schurr et al. 1992 "Native American Mitochondrial DNA Analysis Indicates that the Amerind and the Nadene Populations Were Founded by Two Independent Migrations", Genetics, vol. 130, núm. 1, pp.153-162. [ Links ]
---------- 1993 "Asian Affinities and Continental Radiation of the Four Founding Native American mtDNAs", American Journal Human Genetic, vol. 53, núm. 3, pp. 563-590. [ Links ]
Torroni, Antonio, Yu-Sheng Chen et al. 1994 "mtDNA and Y-Chromosome Polymosphisms in four Native American Populations from Southern Mexico", American Journal Human Genetic, vol. 54, núm. 2, pp. 303-318. [ Links ]
Townsend, Richard F. 1992 Aztecs, Londres, Thames and Hudson, 256 pp. [ Links ]
Zegura, Stephen L., Tatiana M. Karafet et al. 2004 "High-Resolution SNPs and Microsatellite Haplotypes Point to a Single, Recent Entry of Native American Y Chromosomes into the Americas", Molecular Biology and Evolution, vol. 21, núm. 1, pp. 164-175. [ Links ]

