










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCuicuilco
versión impresa ISSN 0185-1659
Cuicuilco vol.18 no.50 México ene./abr. 2011
Dossier
¿Sabe el mono araña el resultado de una suma o entiende la acción de poner?
Bernardo Yáñez* y Pilar Chiappa**
* Centro de Estudios Filosóficos, Políticos y Sociales. Vicente Lombardo Toledano, México.
** Departamento de Etología, Instituto Nacional de Psiquiatría Ramón de la Fuente, México.
Resumen
En este artículo describimos un estudio sobre la representación de magnitudes en monos araña de mano negra (Ateles geoffroyi). Desde hace décadas, los científicos que trabajan en este campo han discutido si la representación es exacta o aproximada. Aquí utilizamos el procedimiento de la violación de la expectativa. Esto implica que un investigador manipula objetos frente a un animal como si actuara "uno más uno igual a uno, dos o tres". Este procedimiento supone que un resultado inesperado provoca más atención que uno esperado. Operativamente, la medida de la atención es el tiempo que el individuo dedica a observar el resultado. Con este procedimiento el investigador puede clasificar los resultados en categorías que derivan de sus hipótesis cognitivas. Por ejemplo, según una hipótesis aritmética, en la suma "uno más uno", el resultado de "dos" es "posible", mientras que los resultados de "uno" y "tres" son "imposible". El investigador compara los tiempos de observación entre categorías y hace inferencias cognitivas. En esta ocasión, clasificamos los resultados de acuerdo con dos hipótesis, una simulativa y otra aritmética. Nuestros resultados apoyan la primera hipótesis. Discutimos a favor de la idea de que la representación de magnitudes en estos animales es de tipo aproximado, implicando un sistema cognitivo que les permite entender acciones como quitar y poner, más que un sentido innato del número que les permite resolver operaciones aritméticas sencillas como sumar y restar.
Palabras clave: representación de magnitudes, violación de la expectativa, primates, mono araña.
Abstract
In this paper we address the magnitude representation in black-handed spider monkeys (Ateles geoffroyi). From decades ago scientists working in this field have discussed if the representation is exact or approximate. Here we used the expectancy violation procedure. This means a researcher manipulating objects in front of an animal, as if acting "one plus one equals one, two or three". This procedure assumes that a non-expected outcome elicits more attention than an expected one. Operationally, the attention measure is the individual time spent observing the outcome. With this procedure the researcher can classify the outcomes into categories derived from cognitive hypotheses. For instance, according to an arithmetical hypothesis, in the sum "one plus one", the outcome "two" is "possible", while the outcomes "one" and "three" are "impossible". The researcher compares the observing time between categories and makes cognitive inferences. We classify the outcome in accordance to a "simulative hypothesis" and an "arithmetical hypothesis". Our data support the first one. We argue in favor of a representational system enabling the understanding of actions like "put on" and "take off" rather than an innate number sense which supports the arithmetic capability of adding and subtracting.
Keywords: magnitude representation, expectancy violation, primates, spider monkey.
LOS ESTUDIOS PREVIOS SOBRE LA REPRESENTACIÓN DE MAGNITUDES
Una de las tramas más intrigantes de las ciencias cognitivas comparadas es la de la representación de magnitudes. En efecto, muchos estudiosos hemos quedado enredados tan sólo con leer algunos títulos, como el de Adición y sustracción en bebés [traducido por los autores (Wynn, 1992)] o el de ¿Qué piensan los animales acerca de los números? [traducido por los autores (Hauser, 2000)]. Tal vez por eso, en la actualidad, dicho tema cuenta con un cúmulo grande y variado de estudios que han enfocado organismos de distintas especies animales y de diversas edades, además de que engloban diferentes metodologías. A continuación presentamos una muestra de este tipo de estudios que, a pesar de estar centrada en las últimas dos décadas, en nuestra opinión es lo suficientemente representativa como para introducir el tema.
Estudios con el procedimiento violación de la expectativa. Wynn [1992] mostró que los bebés de cinco meses fijan la mirada por menos tiempo cuando observan la serie de acciones que representa la operación "uno más uno igual a dos" que cuando observan las acciones correspondientes a las operaciones de "uno más uno igual a uno" y "uno más uno igual a tres". La autora hizo tres experimentos. En el primero, dividió a 32 bebés de aproximadamente cinco meses en dos grupos. Cada bebé del grupo "uno más uno" vio en un escenario: 1. Un objeto; 2. Una pequeña pantalla alzándose y ocultando el objeto; 3. Una mano poniendo un segundo objeto (idéntico al primero) y ubicándolo atrás de la pantalla; y 4. La pantalla bajándose y dejando ver uno o dos objetos finales. La autora repitió seis veces el procedimiento descrito desde el primer paso hasta el cuarto, alternando el número de objetos finales entre uno y dos. Cada bebé del grupo "dos me-nos uno" vio: 1. Un escenario con dos objetos; 2. Una pantalla, alzándose y ocultando los objetos; 3. Una persona quitando uno de los objetos del escenario; y 4. La pantalla bajándose y dejando ver uno o dos objetos finales. Nuevamente, la autora repitió seis veces el procedimiento descrito desde el segundo paso hasta el cuarto, alternando el número de objetos finales entre uno y dos. Después comparó los tiempos de observación del bebé ante los resultados con un objeto y con dos objetos para cada grupo de bebés. Es importante notar que la autora, aunque no lo haya hecho explícitamente, utilizó una "hipótesis aritmética" para clasificar los resultados en "posibles" e "imposibles", siguiendo la lógica de que si el bebé supiera hacer sumas y restas simples, la expectativa sería de "dos" y de "uno", respectivamente. Por tanto, la observación ante un resultado aritméticamente correcto o "posible" duraría menos tiempo que ante un resultado aritméticamente incorrecto o "imposible", como en el caso de "uno más uno igual a uno". Con el segundo experimento la autora replicó sus resultados. En cambio, con el tercero intentó descartar la posibilidad de que los bebés sólo supieran que el resultado de una operación aritmética implica un cambio numérico sin percatarse de la dirección o del tamaño exacto de tal cambio. El procedimiento que usó en este último es similar al de las sumas en los anteriores, ya que sólo varía en que el resultado final puede ser de dos o de tres objetos. En este caso, los resultados también mostraron que los bebés observan los resultados "posibles" durante menos tiempo que aquellos "imposibles". Según la autora, sus experimentos muestran que "los infantes poseen conceptos numéricos verdaderos [esto es] tienen acceso al ordenamiento de las relaciones numéricas entre números pequeños y pueden manipular estos conceptos en modos numéricamente significativos" [traducido por los autores (Wynn, 1992:750)].
Hauser et al. [1996] mostraron que los monos rhesus (Macaca mulatta) cambian el tiempo de observación frente a operaciones con resultados aritméticamente "posibles" e "imposibles", aunque miran por más tiempo el resultado de "uno más uno igual a uno" en comparación con el tiempo en que miran el resultado de "uno más uno igual a dos". Ellos usaron una caja de dos compartimentos (formados por una división baja), sin tapa, con un lado removible y otro de doble fondo para: 1. Poner un objeto en un compartimento y dejar el otro vacío, o bien, poner un objeto en cada compartimento; 2. Colocar el lado removible (obstruyendo la vista del mono hacia el interior de la caja), poner otro objeto por arriba (ya sea en el primer fondo o en el segundo) y remover el lado, mostrando un resultado "posible" o "imposible".
Aunque con algunas variaciones, el procedimiento de violación de la expectativa también ha sido usado en otras especies de primates y ha mostrado resultados que concuerdan con los ya descritos. En efecto, después de ver la operación de "uno más uno", también los tamarinos cabeza de algodón (Saguinus oedipus) observan durante menos tiempo el resultado "dos" que los resultados "uno" y "tres" [Uller et al., 2001]. Asimismo, Hauser y Carey [2003] mostraron que los macacos rhesus de igual modo observan por más tiempo el resultado de "uno más uno igual a tres", que el de "uno más uno igual a dos". Más aún, resultados similares se han obtenido en cuatro especies de prosimios (Eulemur fulvus, Eulemur mongoz, Lemur catta y Varecia rubra) [Santos et al., 2005].
Estudios con el procedimiento habituación. Xu y Spelke [2000] mostraron que los bebés de seis meses de edad pueden discriminar entre conjuntos de ocho y de 16 elementos empleando un procedimiento de habituación. Esto es, sentaron a cada bebé frente a una pantalla donde repetidamente aparecía un nuevo conjunto de puntos. Al principio el nuevo conjunto tenía el mismo número de puntos que el anterior (p. ej. ocho), aunque variara su tamaño o disposición. Eventualmente, el nuevo conjunto tenía otra cantidad de puntos (p. ej. 16). Las autoras compararon la duración de la mirada del bebé sobre el primer conjunto con la duración correspondiente al segundo y encontraron diferencias estadísticamente significativas. A primera vista, estos resultados parecían indicar que los bebés pueden discriminar los conjuntos por el número de puntos que los conforman. Sin embargo, di-versos estudios subsecuentes mostraron que se trata de una discriminación aproximada, ya que depende de ciertas circunstancias. Justamente, la precisión de la discriminación: 1. Disminuye conforme se reduce la proporción entre las dos cantidades que se presentan [Xu y Spelke, 2000]; 2. Aumenta con el desarrollo [Lipton y Spelke, 2003; Xu y Arriaga, 2007]; y 3. Se pierde cuando los números que se comparan son menores que cuatro (p. ej., dos puntos contra cuatro puntos) [Xu, 2003].
Estudios con el procedimiento ir por más. Hauser et al. [2000] lograron que cada uno de 200 macacos rhesus, que viven en semilibertad, viera a una persona mientras ésta metía una rebanada de manzana tras otra, primero en una caja y luego en otra, hasta que cada caja contuviera un total. Al finalizar estas acciones, la persona se alejaba de las cajas para permitir que el mono tomara el contenido de alguna de éstas. Finalmente, los autores analizaron en qué circunstancias la probabilidad de que los monos eligieran la caja con el número mayor de manzanas estaría por arriba del azar y encontraron que los monos eligieron el número mayor cuando la elección estaba entre "uno y dos", "dos y tres", o bien, entre "tres y cuatro"; sin embargo, no distinguieron entre "cuatro y cinco", "cuatro y seis", "cuatro y ocho", ni entre "tres y ocho". Al parecer, los monos podrían distinguir exactamente sólo hasta cuatro rebanadas de manzana.
Con la prueba de ir por más, Feigenson et al. [2002] encontraron que los bebés de aproximadamente un año eligen el bote con más galletas después de haber visto depositar cantidades distintas de galletas en dos botes, cuando comparan "una" con "dos" y "dos" con "tres"; pero eligen al azar cuando comparan "dos" con "cuatro", "tres" con "cuatro" y "tres" con "seis". Estos autores interpretaron que "tres" es la cantidad máxima posible para las representaciones de magnitudes exactas.
Asimismo, Uller et al. [2003] mostraron que las salamandras de espalda roja eligen el conjunto más grande de moscas cuando comparan entre conjuntos de "una" y de "dos" moscas o cuando éstos contienen "dos" y "tres" moscas. Sin embargo, al igual que los bebés, estos animales no parecen distinguir entre conjuntos de "tres" y de "cuatro" moscas, ni entre los de "cuatro" y los de "seis" moscas.
Experimentos de razonamiento numérico no verbal. Entre los estudios sobre la discriminación de magnitudes destaca uno que fue realizado con un grupo de humanos adultos que hablan una lengua en la que no hay palabras para designar de manera exacta ninguna magnitud. En efecto, si bien desde hace mucho tiempo se sabe que hay lenguas con muy pocas palabras para designar magnitudes [Conant, 1896], en realidad hace muy poco que los estudiosos aluden al significado no numérico que estas palabras pueden tener en algunos casos. Por ejemplo, en el caso de los pirahã —de la región amazónica cercana al río Maici en Brasil—, la palabra hói puede usarse para indicar "uno", pero también para indicar "pocos" o "relativamente menor", como en la referencia a dos pescados pequeños en relación con un pescado grande [Gordon, 2004; Frank et al., 2008]. Gordon [2004] llevó a cabo ocho pruebas de razonamiento numérico no verbal con los pirahã. Una de estas pruebas implicaba que el investigador metiera cierta cantidad de dulces en una caja que tenía un dibujo con el mismo número de peces en la tapa. El investigador ponía la caja detrás de su espalda y luego presentaba dos cajas, la original y otra con más pescados en la tapa. Para comparaciones pequeñas como "tres" versus "cuatro", la respuesta correcta rara vez superó 50%.
LAS INTERPRETACIONES CONTEMPORÁNEAS DE LOS ESTUDIOS SOBRE LA REPRESENTACIÓN DE MAGNITUDES
En una revisión relativamente reciente, Feigenson et al. [2004] sostuvieron que la capacidad para la representación de magnitudes que muestran los humanos infantes y otros animales la forman dos sistemas nucleares. El primero de éstos permitiría la distinción de poco y mucho, en otras palabras, correspondería a la capacidad de representar magnitudes de manera aproximada y de relacionarlas por igualdad o desigualdad. En cambio, el segundo sistema permitiría la significación de "uno", "dos" y "tres", o sea, la representación exacta de algunas magnitudes (sólo hasta cuatro) y su operación en sumas o restas sencillas. Sin embargo, como mostramos a continuación, el primer sistema tiene un soporte experimental más sólido que el segundo.
LOS RASGOS DISTINTIVOS DEL SISTEMA DE REPRESENTACIÓN DE MAGNITUDES APROXIMADAS
El sistema de representación de magnitudes aproximadas se distingue porque sigue la ley psicofísica de Weber, que desarrolló Ernst Heinrich Weber en la cuarta década del siglo XIX, que trabajó Gustav Theodor Fechner en la década siguiente y que amplió Stanley Smith Stevens un siglo más tarde [Unidad de Psicología, Departamento de Neurociencia, Universidad de Upsala, 2004].
En pocas palabras, esta ley describe la relación entre la medición de un cambio de magnitud y la intensidad de la percepción de tal cambio. En los estudios originales de Weber, la relación entre la percepción de un cambio de peso no dependía de su magnitud absoluta, sino de su magnitud relativa. Por ejemplo, sería más fácil percibir el incremento de magnitud que hay desde "uno" hasta "dos", que distinguir el que hay desde "1000" hasta "1001". En efecto, aunque la diferencia absoluta en ambos casos sea la misma, la diferencia entre las magnitudes del primer par corresponde a 50% de la magnitud final, mientras que la diferencia de magnitudes en el segundo par corresponde a 0.09% de la magnitud final. La posibilidad de distinguir estos cambios o notabilidad depende de estas proporciones [Fechner, 1912].
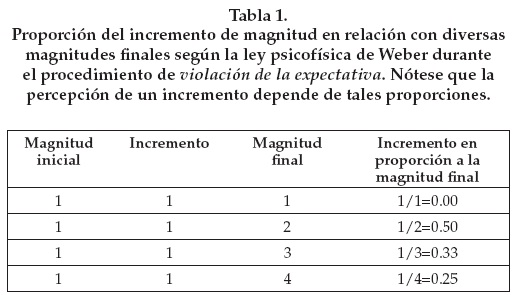
Un sistema de representación basado en esta ley no es eficaz para distinguir incrementos que sean proporcionalmente chicos en relación con la magnitud final. Más aún, cuando un investigador utiliza el procedimiento de violación de la expectativa con las acciones de "uno más uno igual a uno, dos o tres", pone a prueba la percepción de incrementos proporcionalmente distintos. En efecto, la proporción del incremento desde una magnitud inicial de "uno" hasta una magnitud final de "uno" implica que el incremento de "cero" es nulo en relación con la cantidad final; de "dos" implica que el incremento de "uno" es "un medio" en relación con la magnitud final; de "tres" implica un incremento de "dos tercios" en relación con la cantidad final, y así sucesivamente. La percepción de un incremento depende isométricamente de tales proporciones. Por tanto, conforme disminuya la proporción, disminuye su percepción. En la Tabla 1 representamos de otro modo estas relaciones.
LOS RASGOS DISTINTIVOS DEL SISTEMA DE REPRESENTACIÓN DE MAGNITUDES EXACTAS
Según diversos autores, el sistema de representación de magnitudes exactas se puede reconocer porque tiene un límite, que está entre "tres" y "cuatro" [p. ej., Hauser, 2000, 2003; Feigenson et al., 2004]. En efecto, los resultados de los estudios conductuales como los que hemos descrito parecen indicar que los vertebrados, incluyendo a los humanos de cuatro o cinco meses de edad, estamos dotados con una capacidad que nos permite distinguir con exactitud las magnitudes de dos conjuntos de hasta cuatro elementos cada uno. Lo que es más, por lo menos entre los primates, esta capacidad se puede relacionar con el entendimiento de acciones tales como poner o quitar. Además, este sistema de representación no está sujeto a la ley de Weber. En otras palabras, su precisión no muestra un efecto de proporcionalidad.
ALGUNAS INCONGRUENCIAS EN LA LAS INTERPRETACIONES CONTEMPORÁNEAS DE LOS ESTUDIOS SOBRE LA REPRESENTACIÓN DE MAGNITUDES
Hay varias razones que cuestionan la existencia del sistema de representación de magnitudes exactas.
1. El sistema de representación de magnitudes aproximadas podría simular uno de magnitudes exactas. En efecto, el sistema de representación de magnitudes aproximadas permite un buen desempeño perceptual cuando las proporciones resultantes están por encima del umbral de notabilidad. Por ejemplo, si un sujeto a prueba con el procedimiento de violación de la expectativa tiene un umbral de notabilidad ubicado entre un tercio y un cuarto, no podrá distinguir entre el resultado de "uno más uno igual a tres" y el de "uno más uno igual a cuatro" (Tabla 1).
2. El sistema de representación de magnitudes exactas es permeable a la experiencia [Wolters et al. 1987], lo cual pone en duda su origen.
3. Balakrishnan y Asby [1992] mostraron una continuidad en el tiempo de respuesta media a numerosidades con cuatro y más elementos, lo cual refuerza al argumento de la simulación de un sistema por el otro que mencionamos en el párrafo anterior.
4. Los neuroimagenólogos han encontrado y descrito los correlatos neuronales para el sistema de representación de magnitudes aproximadas, pero no han logrado lo mismo para el sistema de representación de magnitudes exactas [Nieder y Miller, 2005; Feigenson et al., 2004].
5. Finalmente, un argumento más en contra de la interpretación de que las capacidades de representación de magnitudes en humanos (bebés y pirahã adultos) y en otras especies de primates surgen de un "sentido del número" [Dehaene, 1997] es que ésta no concuerda con la historia de las matemáticas, cuyo estudio permite establecer que el concepto del número debe considerarse un elemento formal de esta disciplina científica [Lakoff y Núñez, 2001].
A continuación presentamos un intento por satisfacer la necesidad de una interpretación de los resultados del procedimiento de violación de la expectativa que sea congruente con las pruebas derivadas de distintas disciplinas.
HACIA UNA INTERPRETACIÓN INTERDISCIPLINARIAMNTE COHERENTE
En primates, el entendimiento de las acciones observadas puede estar mediado por un sistema de neuronas espejo [Rizzolatti y Sinigaglia, 2006], es decir, por un sistema de neuronas que se activan tanto durante la ejecución de una acción determinada como durante su observación [Gallese et al., 1996]. Con ello en mente, nos preguntamos si durante un procedimiento de violación de la expectativa un animal utiliza un sistema de representación de magnitudes exactas mediado por el entendimiento de la acción "sumar", o bien, usa un sistema de representación de magnitudes aproximadas mediado por el entendimiento de la acción de "poner". En otras palabras, nos propusimos averiguar si los monos araña representan el "uno más uno" como un "sumar" o bien como un "poner". Llamamos a la primera forma de representación hipótesis aritmética, y a la segunda, hipótesis simulativa, y de cada una derivamos una serie de expectativas. Específicamente, según la hipótesis aritmética las acciones "uno igual a uno" y "uno igual a dos" no violarían la expectativa, mientras que las acciones "uno más uno igual a uno" y "uno más uno igual a tres" sí lo harían. En contraste, según la hipótesis simulativa, las acciones "uno igual a uno", "uno más uno igual a dos" y "uno más uno igual a tres" no violarían la expectativa, mientras que la acción "uno más uno igual a uno" sí lo haría. En consecuencia, la categorización de las cuatro acciones en las categorías de "posibles" e "imposibles" sería distinta para cada hipótesis. Operativamente, la comparación estadística del tiempo de observación al resultado entre ambas categorías podría arrojar luz sobre cuál de las dos hipótesis cognitivas explica mejor el comportamiento de los monos.
LOS ASPECTOS METODOLÓGICOS DE ESTE ESTUDIO
Material y métodos
Animales. El estudio que se presenta a continuación implica el comportamiento de seis monos araña de mano negra (Ateles geoffroyi). Estos animales forman parte de un grupo alojado en el Instituto Nacional de Psiquiatría Ramón de la Fuente en la Ciudad de México. Al inicio del estudio el grupo lo integraban tres machos y seis hembras; sin embargo, una hembra murió durante el transcurso de nuestro estudio. El grupo habita en una jaula exterior de concreto, techada con malla metálica, que tiene la forma de una gran rebanada de pastel (6.3 m de alto, 6.00 m de largo, 6.20 m de ancho en el lado mayor y 1.7 m de ancho en el lado menor) y que contiene plataformas, tubos y cuerdas. El lado menor de la jaula está comunicado con un edificio de dos plantas. En la planta baja hay varias puertas, diseñadas para permitir distintas labores, como el mantenimiento de la jaula, la manipulación de los animales o los estudios de cognición. En la planta alta hay una ventana que permite la observación de los animales. Una persona lava diariamente la jaula. Los animales disponen de agua corriente continuamente por medio de un bebedero de palanca y, una vez al día, reciben una dotación de vegetales frescos y croquetas comerciales para monos del Nuevo Mundo. Los ejemplares que forman este grupo tienen dos orígenes: algunos eran mascotas y sus dueños los donaron, mientras que otros los recuperaron del tráfico ilegal las autoridades competentes. Estos orígenes impiden conocer la edad de los animales al inicio del estudio.
Por diversas razones bioéticas estos animales ya no deben ser liberados y, por tanto, se aprovechan para la investigación de algunos aspectos difíciles de abordar en animales silvestres. Como se especifica en el apartado "Diseño experimental", no todos los animales del grupo se utilizaron para este estudio.
Equipo. Una jaula de aluminio cúbica (85 cm por lado) con una pared transparente y sin fondo. Una mesa de madera rectangular (80 cm de largo, 50 cm de ancho y 32 cm de altura). Una tapa cuadrada de madera de cuatro caras (35 cm de largo, 30 cm de ancho y 6 cm de profundidad) sin fondo y sin un lado. Un banco. Una cámara de video digital. Un equipo de cómputo con los siguientes programas: Microsoft Excel, SPSS y Windows Movie Maker. Dos cronómetros. Diversos objetos alimentarios: frutos secos (uvas pasas), semillas (cacahuates), cereales (Froot-loops) y galletas de dos tamaños (Canelitas grandes y mini).
Procedimiento. Usamos el procedimiento conocido como violación de la expectativa, el cual consiste en mostrar un objeto, ocultarlo y añadir o sustraer objetos sin que el individuo a prueba pueda ver el resultado hasta el final. Se trata de un procedimiento bien conocido en la literatura especializada, que se fundamenta en el supuesto de que si el sujeto es capaz de representar el resultado de la acción que está observando, variará su tiempo de observación frente al mismo en función de la comparación de ambos (su propia representación y el resultado que observa), por lo que dicho tiempo será menor cuando perciba concordancia que cuando perciba discrepancia [Wynn, 1992]. La ventaja de este procedimiento es que permite manipular los resultados, haciéndolos "posibles" o "imposibles" según una hipótesis cognitiva. Por ejemplo, con una hipótesis que implica una representación numérico-aritmética, un procedimiento "uno más uno" tiene un resultado posible de "dos" e infinitos resultados "imposibles" como "uno". El experimento comprendió varias sesiones por animal y cada sesión implicó varias operaciones. Los detalles de en cuántas sesiones participó cada animal y de cuántas operaciones vio en total se ofrecen más adelante. Una sesión se iniciaba cuando un primer experimentador abría la puerta de guillotina de la jaula, permitía que uno de los monos del grupo entrara a una jaula sin fondo y cerraba la puerta. Un segundo experimentador se sentaba frente al mono con una mesa en medio de ambos. El primer experimentador empezaba a filmar, enfocando la cara del mono. El segundo experimentador iniciaba una secuencia de acciones. Cada una de éstas consiste en lo siguiente: el segundo experimentador ponía encima de la mesa una cantidad de objetos; luego, con una tapa, ocultaba los objetos de la vista del mono; después, ponía o quitaba objetos, de manera tal que el mono pudiera seguir su acción hasta que la caja se lo impidiera; y finalmente, quitaba la caja de madera, dejando una cantidad de objetos a la vista del mono. Al finalizar la décima operación concluía una sesión. Entonces, el primer experimentador apagaba la cámara, abría la puerta y alentaba al mono para que saliera.
Obtención de datos. A partir de las filmaciones, dos jueces cronometraron el tiempo de observación (expresándolo en segundos) de cada animal frente al resultado de cada acción, o sea, la variable de análisis. Obtuvimos el coeficiente de concordancia de Kendall entre los juicios de los jueces, el cual, estando en el límite de la significación estadística (tau_b de Kendall= 0.8, p= 0.05, N= 5), indica que sus mediciones del tiempo de observación al resultado concuerdan bastante y fundamenta el uso de los registros de un solo "juez" en los análisis posteriores. A diferencia de los experimentadores, los jueces no sabían a qué operación correspondía la observación que estaban cronometrando, es decir, los jueces eran "ciegos", en el sentido de que tuvieron acceso a la filmación sólo desde el momento en que se destapó el resultado.
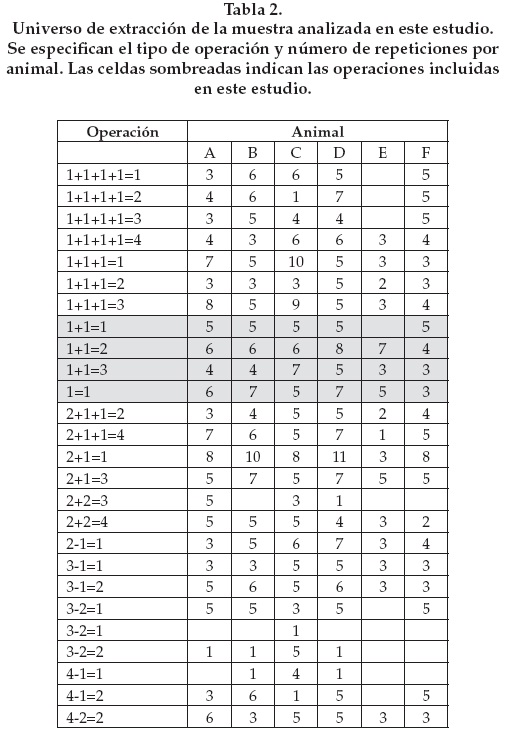
Tamaño de la muestra. Originalmente planeamos obtener cinco valores del tiempo de observación expresado en segundos para cada operación y para cada uno de los nueve animales que conforman el grupo; sin embargo, 1. La participación de los monos era voluntaria, y 2. Una mona murió durante el transcurso del experimento. De este modo, al final de un periodo de seis semanas de intentos diarios sólo participaron seis animales, de los cuales, algunos sobrepasaron las cinco repeticiones y otros no las alcanzaron, por lo que cada uno vio un número distinto de operaciones y un número distinto de veces cada operación (Tabla 2). Con la finalidad de balancear el número de repeticiones por animal, los análisis estadísticos que presentamos incluyen sólo tres registros de tiempo de observación al resultado de una operación por cada individuo, expresado en segundos. Además, para evitar al máximo el error de cronometraje (v. el apartado "Obtención de datos"), éstos corresponden a los valores mayores o tiempos mayores, pero cabe aclarar que esta manipulación no afecta el resultado de las pruebas estadísticas (Tabla 3).

Análisis de datos. Agrupamos estos datos en dos pares de categorías. El primer par corresponde a la agrupación de nuestras acciones en "posibles" e "imposibles" de acuerdo con las expectativas de la hipótesis aritmética, mientras que el segundo corresponde a tal agrupación de acuerdo con las expectativas de la hipótesis simulativa. Esta categorización implica que una operación puede ser posible simulativamente, pero imposible aritméticamente. Con ello obtuvimos cuatro variables (Tabla 4). Finalmente, aplicamos la prueba de rangos asignados de Wilcoxon para comparar los tiempos de observación "posibles" e "imposibles" según cada hipótesis.

RESULTADOS
La prueba de rangos asignados de Wilcoxon indica que no hay diferencias estadísticamente significativas entre el tiempo de observación a los resultados aritméticamente posibles y el correspondiente a los imposibles (xpos = 1.09 ± 0.38 y ximpo = 1.36 ± 0.58: z=-0.94, p=0.345). En contraste, dicha prueba indicó diferencias significativas entre el tiempo de observación a los resultados simulativamente posibles en comparación con el correspondiente a los imposibles (xpos= 0.71 ± 0.25 y ximpo= 2.61 ± 0.96: z=-2.02, p=0.043) (Gráfica 1).

DISCUSIÓN
Este artículo describe un estudio sobre las capacidades para la discriminación de magnitudes realizado con monos araña. Usamos el procedimiento conocido como violación de la expectativa, el cual permite manipular los resultados de manera que éstos sean "imposibles" (cuando "violan" una expectativa cognitiva determinada) o "posibles" (cuando éstos "concuerdan" con tal expectativa). Determinamos las expectativas de una hipótesis aritmética y una hipótesis simulativa. Este abordaje es novedoso, ya que, hasta donde sabemos, ningún otro autor ha publicado un estudio donde se pruebe una hipótesis simulativa con este procedimiento. Además, ningún otro autor ha explicitado que sus predicciones se derivaron de una hipótesis aritmética.
Esto abre la posibilidad de futuros análisis sobre el comportamiento asociado a la presentación del resultado final como un indicativo del entendimiento de lo que ocurre.
Nuestros resultados muestran que la hipótesis simulativa explica mejor el comportamiento de los monos araña que la hipótesis aritmética, apoyando la idea de que la representación de magnitudes en estos animales implica un sistema cognitivo que les permite entender acciones como quitar y poner, más que un sentido del número que les permita hacer operaciones aritméticas sencillas, como sumar y restar.
Agradecimientos.
Agradecemos a Chiara Casti, Elizabeth Hernández y Diana Gómez su participación en la conducción de los experimentos, y a Óscar Rosales su ayuda con los programas electrónicos. Esta investigación corresponde al Proyecto NC093320.4 del Instituto Nacional de Psiquiatría Ramón de la Fuente.
BIBLIOGRAFÍA
Balakrishnan, J. D. y F. Gregory Ashby 1992 "Subitizing: Magical numbers or mere superstition?", en Psychological Research, vol. 54, pp. 80-90. [ Links ]
Conant, Levi Leonard 1896 The number concept: its origins and development, Nueva York, McMillan Company. [ Links ]
Dehaene, Stanislas 1997 The number sense, Nueva York, Oxford University Press. [ Links ]
Fechner, Gustav Theodor 1912 "Elements of psychophysics. Sections VII and XVI", Herbert Sidney Langfeld (trad.), en Green C. D., Classics in the History of Psychology. An internet resource, York University, Toronto [http://psychclassics.yorku.ca/Fechner/]. Consultado el 16 de mayo de 2011. [ Links ]
Feigenson, Lisa; Susan Carey et al. 2002 "The representations underlying infants choice of more: object files versus analog magnitudes", en Psychological Science, vol. 13, pp. 150–156. [ Links ]
Feigenson, Lisa; Stanislas Dehaene et al. 2004 "Core systems of number", en Trends in Cognitive Sciences, vol. 8, pp. 307–314. [ Links ]
Frank, Michael C.; Daniel L. Everett et al. 2008 "Number as a cognitive technology: evidence from Pirahã language and cognition", en Cognition, vol. 108, pp. 819–824. [ Links ]
Gallese, Vittorio; Luciano Fadiga et al. 1996 "Action recognition in the premotor cortex", en Brain, vol. 119, pp. 593–609. [ Links ]
Gordon, Peter 2004 "Numerical cognition without words: evidence from Amazonia", en Science, vol. 306, pp. 496–499. [ Links ]
Hauser, Marc D. 2000 "What do animals think about numbers?", en American Scientist, vol. 8, pp. 144-151. [ Links ]
---------- 2003 "Primate cognition", en Gallaher, M., R. J. Nelson y Weiner I. B. (eds.), Handbook of Psychology, Hoboken, John Wiley and Sons, pp. 561-594. [ Links ]
Hauser, Marc D.; Pogen Macneilage et al. 1996 "Numerical representations in primates", en Proccedings of National Academy of Sciences, vol. 93, pp. 1514–1517. [ Links ]
Hauser, Marc D. y Susan Carey 2003 "Spontaneous representations of small numbers of objects by rhesus macaques: examinations of content and format", en Cognitive Psychology, vol. 47, pp. 367–401. [ Links ]
Hauser, Marc D.; Susan Carey et al. 2000 "Spontaneous number representation in semi-free-ranging rhesus monkeys", en Proceedings of the Royal Society of London B, vol. 267, pp. 829–833. [ Links ]
Lakoff, George y Rafael E. Núñez 2001 Where mathematics come from, Nueva York, Basic Books. [ Links ]
Lipton, Jennifer S. y Elizabeth S. Spelke 2003 "Origins of number sense: large-number discrimination in human infants", en Psychological Science, vol. 14, pp. 396–401. [ Links ]
Nieder, Andreas y Earl K. Miller 2005 "Neural correlates of numerical cognition in the neocortex of nonhuman primates", en S. Dehaene, J-R Duhamel, M. Hauser y G. Rizzolatti (eds.), From monkey brain to human brain, Cambridge, MIT. [ Links ]
Rizzolatti, Giacomo y Corrado Sinigaglia 2006 Las neuronas espejo, Barcelona, Paidós. [ Links ]
Santos, Laurie R.; Jennifer L. Barnes et al. 2005 "Expectations about numerical events in four lemur species (Eulemur fulvus, Eulemur mongoz, Lemur catta and Varecia rubra)", en Animal Cognition, vol. 8, pp. 253–262. [ Links ]
Uller, Claudia; Marc Hauser et al. 2001 "Spontaneous representation of number in cotton-top tamarins (Saguinus oedipus)", en Journal of Comparative Psychology, vol. 115, pp. 248–257. [ Links ]
Uller, Claudia; Robert Jaeger et al. 2003 "Salamanders (Plethodon cinereus) go for more: rudiments of number in an amphibian", en Animal Cognition, vol. 6, pp. 105–112. [ Links ]
Unidad de Psicología, Departamento de Neurociencia, Universidad de Upsala 2004 Weber's law. [http://www.neuro.uu.se/fysiologi/gu/nbb/lectures/WebFech.html]. Consultado el 16 de mayo de 2011. [ Links ]
Wolters, Gezinua; Hanneke van Kempen et al. 1987 "Quantification of small numbers of dots: subitizing or pattern recognition?", en American Journal of Psychology, vol. 100, pp. 225–237. [ Links ]
Wynn, Karen 1992 "Addition and substraction by human infants", en Nature, vol. 358, pp. 749–750. [ Links ]
Xu, Fei 2003 "Numerosity discrimination in infants: evidence for two systems of representations", en Cognition, vol. 89, pp. B15–B25. [ Links ]
Xu, Fei y Elizabeth S. Spelke 2000 "Large number discrimination in 6–month old infants", en Cognition, vol. 74, pp. B1–B11. [ Links ]
Xu, Fei y Rosa I. Arriaga 2007 "Number discrimination in 10-month-old infants", en British Journal of Developmental Psychology, vol. 25, pp. 103-108. [ Links ]

