











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
INTRODUCCIÓN
La presencia de aves en un ecosistema está estrechamente ligada a la calidad del hábitat, por lo que pueden ser excelentes indicadores del estado de conservación o perturbación (Arizmendi, 2001), tal es el caso del semillero rabadilla canela (Sporophlia torqueola), el tirano pálido (Tyrannus verticalis) y el garrapatero pijuy (Crotophaga sulcirostris), que se asocian generalmente a hábitats abiertos como potreros; también, sobresalen la paloma arroyera (Leptotila verreauxii) y la chachalaca oriental (Ortalis vetula) en vegetación secundaria, mientras que en bosque tropical lluvioso se ha destacado a la pía guardabosques (Lipaugus unirufus) y el flautín cabezón mesoamericano (Schiffornis turdinus), todas encontradas en el Sureste de México (González-Valdivia et al., 2011). La importancia ecológica de las aves radica en su papel como polinizadoras, dispersoras de semillas, controladoras de plagas de insectos o roedores, depredadoras de mamíferos y reptiles, y eliminación de cadáveres, que son procesos esenciales en la dinámica de los ecosistemas y cultivos (Wenny et al., 2011). Además, por ser un grupo que abarca especies llamativas y carismáticas, fomenta el turismo de observación de aves con una importante derrama económica equivalente a aproximadamente 23.9 millones de dólares al año en México (Cantú & Sánchez, 2011; Revollo-Fernández, 2015); particularmente se ha reportado que, en dos reservas importantes de Yucatán como Celestún y Ría Lagartos, el aviturismo genera alrededor de 17 millones de dólares (Cantú et al., 2011). Cabe destacar que, en la Península de Yucatán, aún se mantienen la tradición de utilizar sus recursos faunísticos para alimentarse mediante cacería de subsistencia como el hocofaisán (Crax rubra), el guajolote ocelado (Meleagris ocellata), la pava cojolita (Penelope purpurascens) y la chachalaca oriental (O. vetula), así como para fines medicinales, rituales y festividades (Chablé & Delfín, 2010; Ramírez-Barajas & Calmé, 2015).
México posee alrededor de 1,119 especies de aves, esto corresponde a 10.6 % de las que hay en el mundo (Berlanga et al., 2019), de las cuales entre el 18 y 20 % son endémicas del país (Navarro-Sigüenza et al., 2014; Berlanga et al., 2019). La mayor concentración de especies se presenta en la vertiente del Golfo de México y en la Península de Yucatán (Navarro-Sigüenza et al., 2014). En esta última se han registrado 543 especies, de las cuales 484 habitan en Quintana Roo (MacKinnon, 2005; Correa-Sandoval & MacKinnon, 2011).
A pesar de dicha riqueza de especies, el estatus de conservación de las aves en la Península de Yucatán ha cambiado aceleradamente en las últimas décadas debido al crecimiento poblacional humano, a la urbanización, al cambio de uso del suelo (ganadería y plantaciones), y al desarrollo turístico e inmobiliario, lo cual ha trasformado tres cuartas partes de la cobertura del suelo original (Calmé et al., 2015; Dupuy et al., 2015; Porter-Bolland et al., 2015). Amenazas como la extracción excesiva de madera, la pérdida de bosques, la contaminación y la introducción de especies exóticas, promueven la disminución y extinción de poblaciones locales, por lo que deben tomarse en consideración en las estrategias de conservación. Debido a esta situación, entre 298 y 388 especies de la avifauna mexicana (26 al 33 %, respectivamente) se encuentran en alguna categoría de riesgo, tanto en el ámbito nacional como internacional (Navarro-Sigüenza et al., 2014).
En México se reproduce el 77 % de las especies de aves registradas y que son reconocidas como residentes. No obstante, también se tienen visitantes de invierno y migratorias de paso (Navarro-Sigüenza et al., 2014). Esto hace que sea de suma importancia evaluar el estado de conservación de las comunidades de aves mediante estudios de diversidad. Predicciones generales resaltan que hábitats naturales e incluso tierras de cultivos incrementan la riqueza de especies de aves, así como la uniformidad (i. e. eveness, igualdad numérica de aves por especie en la comunidad), contrario a los hábitats urbanizados o con recursos deteriorados para las aves (Tu et al., 2020). Algunos estudios indican que la urbanización por sí misma no reduce la riqueza de especies de aves (i. e. número de especies) mientras haya abundantes recursos alimenticios, pero sí incrementa el número de aves o individuos de unas pocas especies dominantes en áreas urbanas (Bezzel, 1985; Jokimäki et al., 1996). Por ello, el objetivo de nuestro estudio fue determinar la diversidad y composición de aves en respuesta a diferentes perturbaciones de acuerdo con el estado de la vegetación en el paisaje, en un ejido forestal de Quintana Roo, México.
MATERIALES Y MÉTODOS
Área de estudio. El área de estudio se ubica en el ejido Laguna Om (8º 44’ 47” y 18º 26’ 39” N y 88º 57’ 57” y 88º 56’ 13” E; Serrano, 2013). El clima es cálido subhúmedo con una temperatura y precipitación promedio anual de 26 °C y 1,290 mm, respectivamente (Sistema Meteorológico Nacional, 2010; Serrano, 2013). Presenta varios tipos de vegetación, incluyendo selvas más o menos conservadas, como la selva baja espinosa subperennifolia o la selva mediana subperennifolia (por mencionar algunas), áreas cultivadas y potreros. También hay lagunas, sabanas y aguadas (Macario-Mendoza, 2020). El área de estudio es accesible desde la carretera Nuevo Bécar-Caobas, que cruza el ejido de norte a sur, y que es mucho menos transitada que la carretera Escárcega-Chetumal que cruza el ejido de oeste a este (Fig. 1).
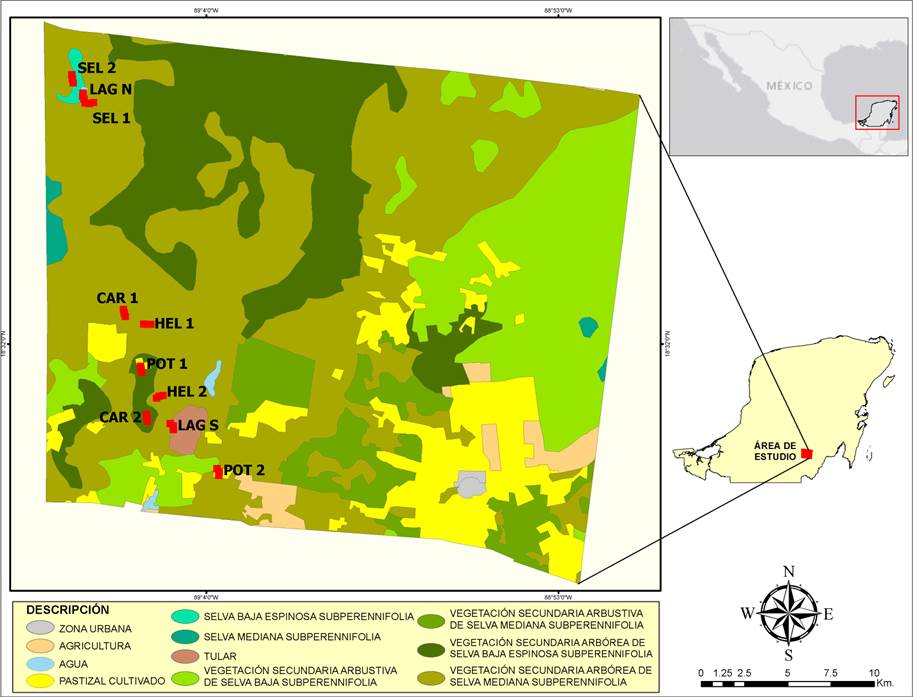
Figura 1 Ubicación del Ejido Laguna Om, Quintana Roo, México, con los tipos de vegetación. Los sitios de muestreo marcados con rojo.
Diseño del muestreo. Seleccionamos cinco tipos de hábitat a escala de paisaje con elementos particulares de acuerdo con el grado de perturbación o presencia de agua, siendo en este caso: Selva (SEL) con vegetación de selva mediana subperennifolia poco perturbada, Laguna (LAG) que corresponde a selva mediana subperennifolia situada alrededor de un lago, Carretera (CAR) correspondiente a la vegetación situada a las orillas de una carretera poco concurrida, Potero (POT) con influencia de zonas abiertas de uso ganadero y Helechal (HEL) correspondiente a rodales del helecho cilantrillo (Pteridium aquilinum). Cabe mencionar que cada uno de estos elementos del paisaje distingue al sitio puntual donde se efectuó el muestreo. En cada hábitat ubicamos dos transectos haciendo un total de 10 transectos. En cada transecto de 700 m, se establecieron tres estaciones de monitoreo separadas entre sí por 200 m aproximadamente. Debido a las limitaciones de tiempo y recursos, en cada transecto se realizaron tres muestreos durante el periodo de agosto a septiembre de 2017.
Muestreo de conteo por puntos. Utilizamos el método de conteo por puntos para documentar la presencia de especies y la composición en relación con los diferentes tipos de hábitat, de esta forma estimamos la diversidad, la abundancia y la riqueza para algunas especies con suficientes datos. En cada punto, las observaciones se hicieron por un lapso de 10 minutos con ayuda de un técnico especializado en avifauna local, contando todas las especies observadas o escuchadas en un radio de 25 metros. El esfuerzo de muestreo para el registro de las especies fue de dos observadores por punto de muestreo, realizando tres visitas en cada punto de conteo, donde se registró la especie, número de individuos, sexo, el transecto y el punto de escucha. Las observaciones se hicieron desde el amanecer en la mañana, y por la tarde en el pico de la actividad de las aves, esto con la finalidad de identificar más especies de aves mediante el esfuerzo de muestreo.
Procesamiento de datos. Realizamos una base de datos con todas las especies observadas y escuchadas durante las sesiones de muestreo. A su vez, obtuvimos datos de abundancia relativa, así como de los gremios tróficos y el estatus migratorio de las especies de acuerdo con MacKinnon (2013) y González-Salazar et al. (2014). Durante el período de muestreo concurren diferentes estados de residencia de las aves: residentes permanentes, residentes invernales que llegan del norte en julio, residentes de verano que vienen de América del Sur y se preparan para irse en octubre, y algunas que solo están de paso por la Península de Yucatán durante su migración (MacKinnon, 2013).
Abundancia relativa. Para obtener la abundancia relativa en cada tipo de hábitat, realizamos curvas de rango abundancia para cada tipo de hábitat y para el muestreo en general (Feinsinger, 2004). Este gráfico nos ayuda a ordenar las especies en rango de mayor a menor abundancia, para comparar aspectos de la distribución de las abundancias, dominancia y uso de recursos en la comunidad.
Gremios tróficos. Realizamos una categorización de los gremios tróficos de cada una de las especies con base en el estudio de González-Salazar et al. (2014), agrupando a las especies dentro de alguna de las categorías acorde con su dieta principal: frugívoros (e. g. chachalacas, codornices), herbívoros, insectívoros (e. g. chipes, vencejos, golondrinas, mosqueros), nectarívoros (e. g. colibríes), carnívoros (e. g. aguilillas, halcones, búhos), granívoros (e. g. rascadores, gorriones), carroñeros (e. g. zopilotes) y omnívoros (e. g. zanates, charas); así mismo, agregamos la categoría de piscívoro debido a la presencia de aves en las zonas lacustres que mayormente se alimentan de peces (De Dios-Arcos, 2014).
Estatus migratorio. Para definir la cualidad migratoria se usaron las clases de Berlanga et al. (2015; 2019) que dividen a las especies de acuerdo con su residencia de la siguiente manera: 1) residentes permanentes (R), sobreviven en una misma región durante un ciclo anual; 2) migratorias de invierno (MI), se reproducen en Estados Unidos de América y Canadá en el verano y pasan el invierno en México, Centroamérica y Sudamérica; 3) migratorias de verano (MV), visitan México únicamente para reproducirse (primavera-verano); 4) accidentales (A), su área de distribución habitual no corresponde con los límites geográficos de México; 5) transitorias (T) durante la migración van de paso por nuestro país para dirigirse a sus áreas de invernación al sur en el otoño o hacia sus áreas de reproducción en el norte durante la primavera; 6) oceánicas (O), especies pelágicas que pasan la mayor parte del tiempo en el mar abierto.
Endemismo y estado de conservación. Las categorías de endemismo que tomamos en cuenta fueron las definidas por Berlanga et al. (2015; 2019): endémicas (EN), su distribución geográfica se restringe a los límites políticos del territorio mexicano; semiendémicas (SE), se distribuyen en México solamente durante cierta época del año; cuasiendémicas (CE), se distribuyen ligeramente fuera de México hacia algún país vecino. Tomamos en cuenta las categorías de la NOM-059- SEMARNAT-2010 a nivel nacional y de la UICN a nivel mundial (UICN, 2020; SEMARNAT, 2010).
Análisis estadísticos. Para evaluar la completitud del muestreo de las aves del área de estudio se realizó una curva de cobertura de la muestra que mide la proporción que representan los individuos de cada especie en la muestra, con respecto al número total de individuos registrados (Chao & Jost, 2012). Este análisis se llevó a cabo utilizando el programa iNext (Hsieh et al., 2013). Los análisis se realizaron mediante 100 aleatorizaciones y extrapolando a 500 individuos (Chao & Jost, 2012). Para interpretar la estructura del ensamble de la comunidad de aves se empleó la serie de números de diversidad de Hill (Hill, 1973) utilizando la abundancia de las especies. Este análisis permite detectar la magnitud con la que contribuye cada grupo de especies (por ejemplo, abundantes y medianamente abundantes). Se tomó en cuenta la diversidad de orden 0 (riqueza de especies), orden 1 (muestra las especies típicas) y orden 2 (expresa el número de especies dominantes). Para obtener la serie de números de Hill se utilizó el programa estadístico RStudio, versión 1.4.1106 (Rstudio Team, 2021).
Para comparar la similitud en la estructura de la comunidad entre sitios de muestreo dentro de cada hábitat se realizó una prueba no paramétrica de Escalamiento No Métrico Multidimensional (NMDS) utilizando una matriz de similitud de Bray-Curtis, donde 0 indica que no hay similitud y 100 indica máxima similitud (Clarke & Warwick, 2001). Esta prueba permite identificar los patrones de variación de la composición de las especies, además de que muestra una configuración espacial que da una idea de la relación de similitud entre los diferentes tipos de hábitat (Field et al., 1982). Para complementar la prueba NMDS, se realizó un Análisis Multivariado de Similaridad (ANOSIM) basado en las abundancias relativas de cada especie. Este permite obtener un valor de prueba estadístico que define si las diferencias entre los tipos de hábitat fueron significativas, para este análisis también se utiliza la misma matriz de similitud antes mencionada (Clarke & Warwick, 2001). Para ello, utilizamos el programa estadístico PAST versión 4.05.
RESULTADOS
Se registraron un total de 103 especies distribuidas en 38 familias. Tyrannidae fue la más abundante con 11 especies; 14 familias estuvieron representadas por un solo individuo de una especie (Fig. 2). Las especies más representativas fueron el tirano dorso negro (Tyrannus tyrannus), seguida del loro cachete amarillo (Amazona autumnalis), perico pecho sucio (Eupsittula nana) y el luis gregario (Myiozetetes similis) (Fig. 3). La curva de cobertura de la muestra arrojó que el inventario tuvo una completitud por arriba del 90 %, excepto en la selva, donde fue del 87 % (Fig. 4; Cuadro 1).
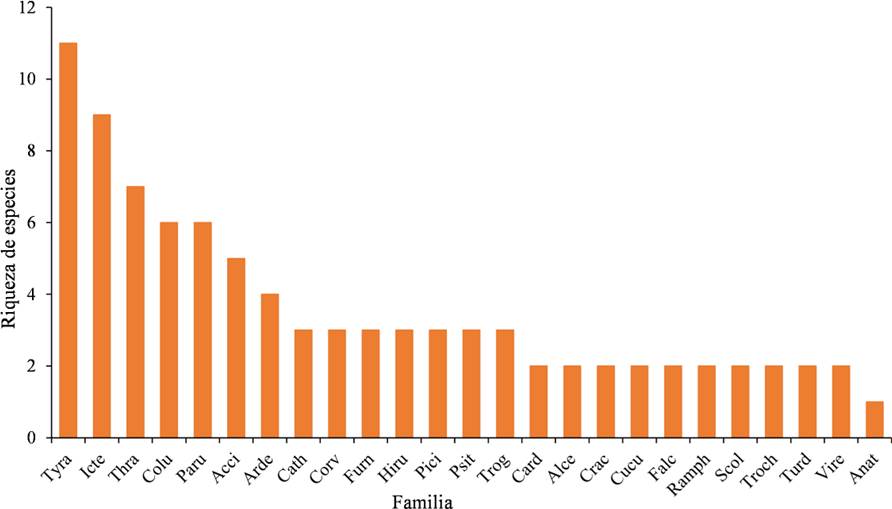
Figura 2 Riqueza de especies de aves por familia en el ejido Laguna Om. Abrev: Tyrannidae (Tyra), Icteridae (Icte), Thraupidae (Thra), Columbidae (Colu), Parulidae (Paru), Accipitridae (Acci), Ardeidae (Arde), Cathartidae (Cath), Corvidae (Corv), Furnariidae (Furn), Hirundinidae (Hiru), Picidae (Pici), Psittacidae (Psit), Troglodytidae (Trog), Cardinalidae (Card), Cerylidae (Cery), Cracidae (Crac), Cuculidae (Cucu), Falconidae (Falc), Ramphastidae (Ramph), Scolopacidae (Scol), Trochilidae (Troch), Turdidae (Turd), Vireonnidae (Vire) y Anatidae (Anat) con una sola especie en el muestreo junto con otras 13 familias (Aramidae, Cotingidae, Jacanidae, Mimidae, Momotidae, Odontophoridae, Oxyruncidae, Paserellidae, Phalacrocoracidae, Rallidae, Tinamidae, Tityridae y Trogonidae).
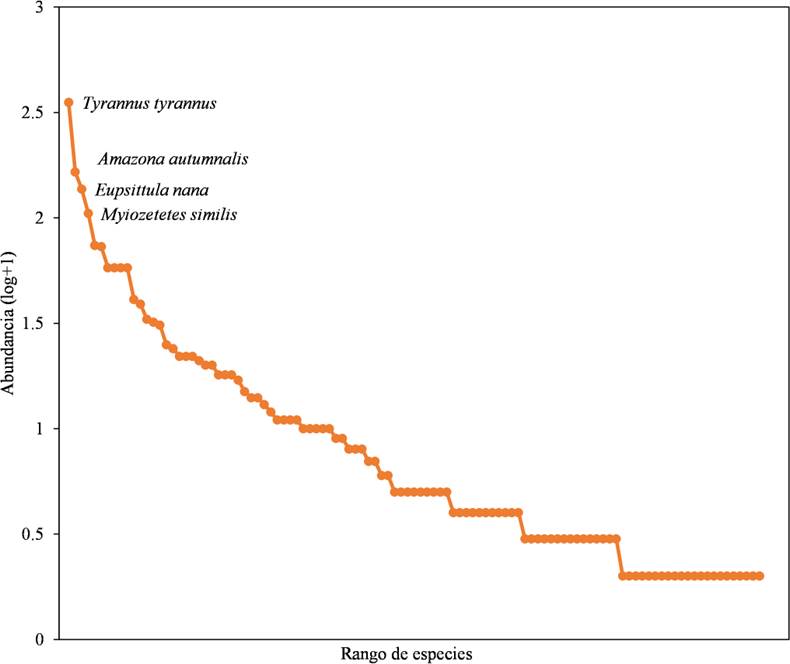
Figura 3 Curva de rango-abundancia del total de las especies de aves del ejido Laguna Om, Quintana Roo, México. El eje x representa la riqueza y el eje y la abundancia relativa de las especies.
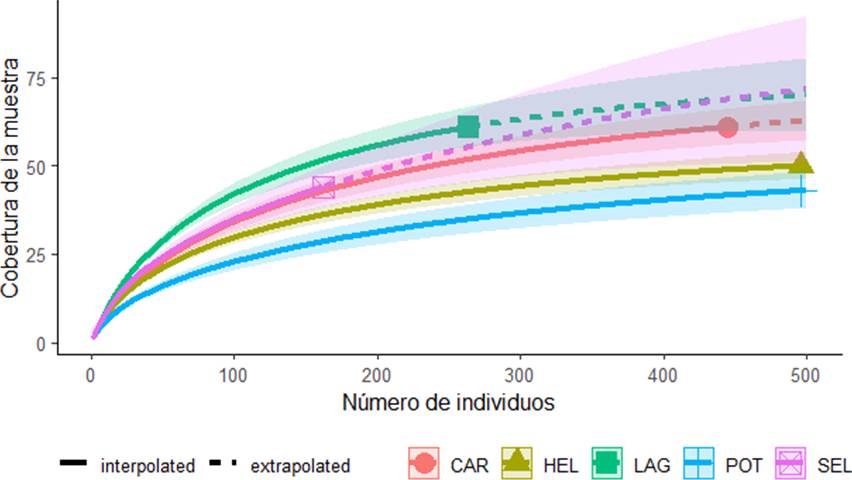
Figura 4 Cobertura de la muestra de aves del ejido Laguna Om, Quintana Roo, México. El eje x representa el número de individuos y el eje y representa la cobertura de la muestra. Abrev: Carretera (CAR), Helechal (HEL), Laguna (LAG), Potrero (POT) y Selva (SEL).
Cuadro 1 Números de Hill y completitud del muestreo de aves. Donde q0 es la riqueza de especies, q1 muestra las especies típicas y q2 expresa el número de especies dominantes en la muestra.
| Hábitat | q0 | q1 | q2 | Completitud |
|---|---|---|---|---|
| Laguna | 61 | 39.02 | 26.48 | 0.9322 |
| Selva | 44 | 26.18 | 17.66 | 0.8708 |
| Potrero | 43 | 11.31 | 4.83 | 0.9759 |
| Helechal | 50 | 21.53 | 11.19 | 0.9799 |
| Carretera | 61 | 26.26 | 13.96 | 0.9642 |
La diversidad de orden 0 para la Laguna fue de 61 especies, de las cuales 39 fueron más abundantes (q1), y de estas, 26 fueron dominantes (q2); la Laguna y la Carretera fueron los hábitats con mayor riqueza y, de acuerdo con los intervalos de confianza al 95 %, hubo diferencias estadísticamente significativas entre este último sitio y los demás (Fig. 5; Cuadro 1).
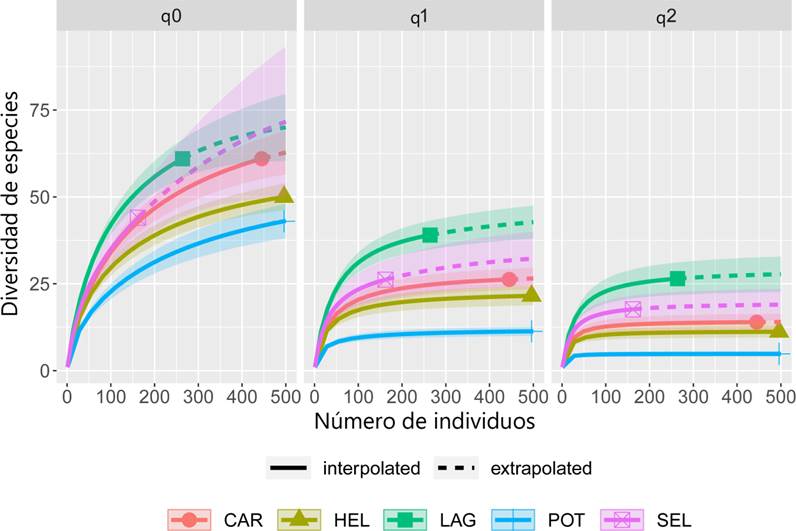
Figura 5 Diversidad verdadera o serie de números de Hill de las aves del ejido Laguna Om. Abrev: q0 = la riqueza de especies, q1 = especies típicas y q2 = especies dominantes en la muestra. Carretera (CAR), Helechal (HEL), Laguna (LAG), Potrero (POT) y Selva (SEL).
Abundancia
La composición de especies varió significativamente en cada hábitat (ANOSIM global R = 0.5106, p < 0.001; Fig. 6). El NMDS muestra que el Potrero presentó una composición de especies diferente a los demás tipos de hábitat. A su vez, el Helechal con la Carretera tienen una composición de especies más similar, mientras que la Selva con la Laguna también, aunque ambos pares muestran diferencias significativas en la composición de sus especies (stress = 0.2696) (Fig. 6).
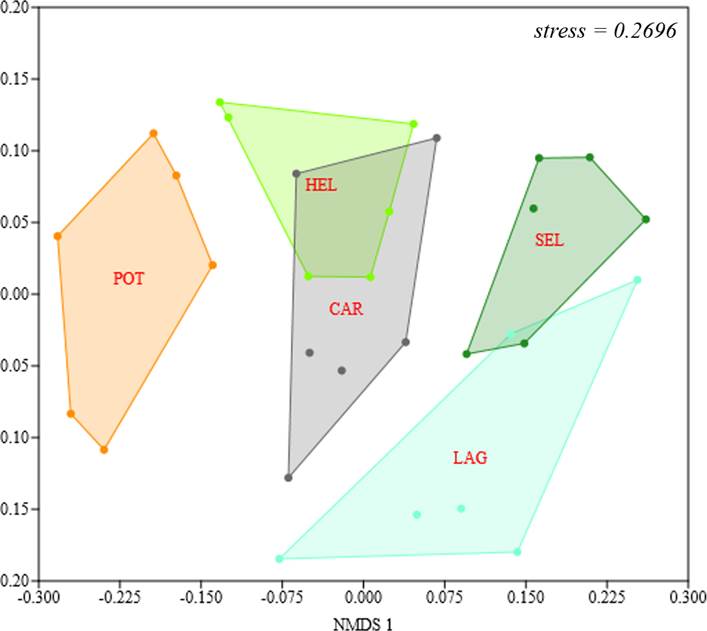
Figura 6 Variación de la composición de las especies de aves entre los sitios de muestreo en cada tipo de hábitat del ejido Laguna Om, Quintana Roo, México. Abrev: Potrero (POT), Helechal (HEL), Carretera (CAR), Selva (SEL) y Laguna (LAG).
La curva de rango-abundancia por tipos de hábitat muestra que en los hábitats Potrero, Helechal y Carretera hay menor equitatividad en la repartición de recursos, siendo pocas especies (aunque más abundantes) las que compiten fuertemente por los recursos, como el tirano dorso negro, el loro cachete amarillo, el perico pecho sucio y la chachalaca vetula, ya que la inclinación de la pendiente en estas curvas es muy pronunciada; contrario a lo que ocurre en la Laguna y la Selva, donde las curvas un poco más homogéneas, lo que sugiere una repartición de recursos es más equitativa (Fig. 7).
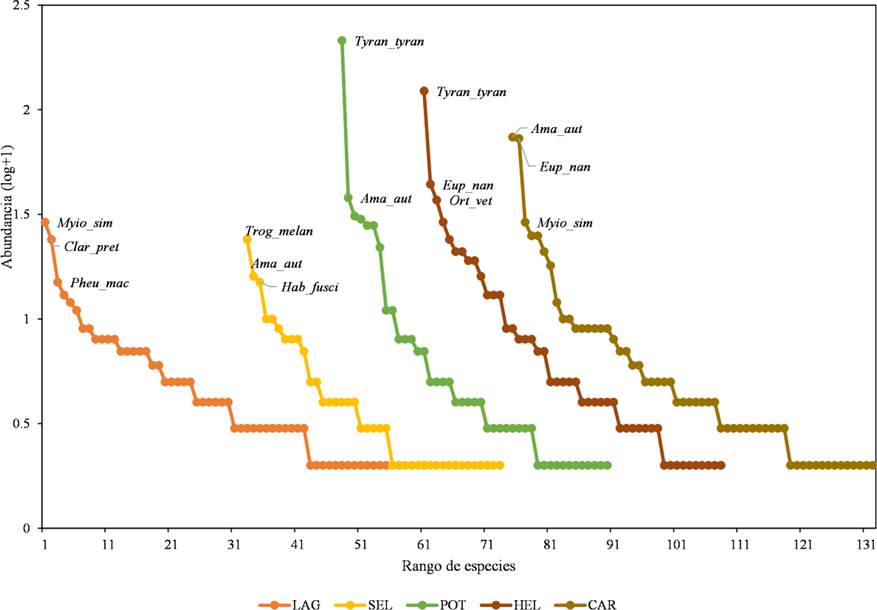
Figura 7 Curva de rango-abundancia de las especies de aves por tipo de hábitat en el ejido Laguna Om. El eje x representa la riqueza de especies mientras que el eje y muestra la abundancia relativa de las especies. Abrev: Myiozetetes similis (Myio_sim), Claravis pretiosa (Clar_pret), Pheugopedius maculipectus (Pheu_mac), Trogon melanocephalus (Trog_melan), Amazona autumnalis (Ama_aut), Habia fuscicauda (Hab_fusci), T. Tyrannus (Tyran_tyran), Eupsittula nana (Eup_nan), Ortalis vetula (Ort_vet). Laguna (LAG), Selva (SEL), Potrero (POT), Helechal (HEL) y Carretera (CAR).
Gremios tróficos
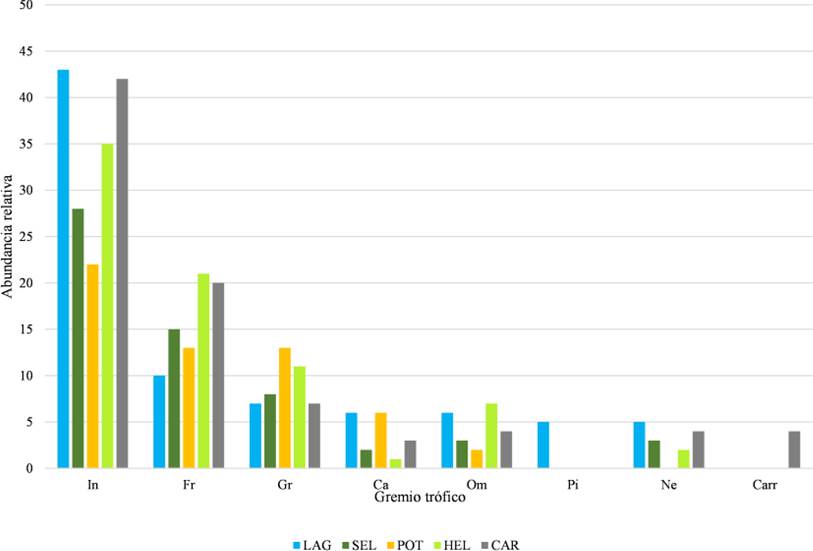
En cuanto a los gremios tróficos, encontramos que el grupo más representativo fue el de los insectívoros con 53 especies, seguido de los frugívoros (14 especies) y de los granívoros (11 especies; Fig. 8; Cuadro 2). Laguna fue el hábitat que presentó mayor número de especies insectívoras, seguido de Carretera, Helechal y Selva. Las aves frugívoras, por su parte, fueron más abundantes en Helechal y Carretera; las granívoras fueron más abundantes en Potrero y Helechal; las carnívoras estuvieron más presentes en Laguna y Potrero; las omnívoras en Laguna y Helechal, las piscívoras solamente estuvieron presentes en Laguna, las nectarívoras predominaron en Laguna y Carretera; y finalmente, las carroñeras solamente se registraron en Carretera (Fig. 9).
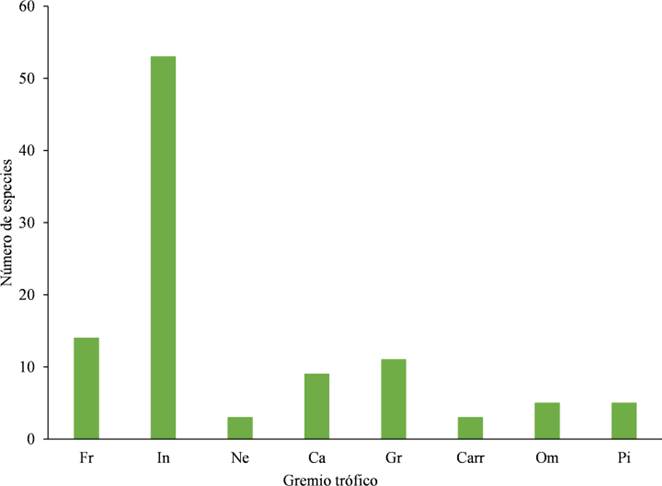
Figura 8 Riqueza de especies de aves en los Gremios tróficos en el ejido Laguna Om. Abrev: Frugívoro (Fr), Insectívoro (In), Nectarívoro (Ne), Carnívoro (Ca), Granívoro (Gr), Carroñero (Carr), Omnívoro (Om) y Piscívoro (Pi).
Cuadro 2 Lista de especies de aves registradas en el ejido Laguna Om. Se muestra la clasificación de los gremios tróficos de las aves, así como el tipo de residencia que tiene cada una en México basado en Berlanga et al. (2015). Abrev: Gremio trófico: Frugívoros (Fr), Insectívoros (In), Nectarívoros (Ne), Carnívoros (Ca), Granívoros (Gr), Piscívoros (Pi), Carroñeros (Carr), Omnívoros (Om). Estatus migratorio (Berlanga et al., 2019): Residentes permanentes (R), Migratorias de invierno (MI), migratorias de verano (MV), Transitorias (T) y Accidentales (A). Categoría de riesgo (NOM-059-Semarnat-2010): Probablemente extinta en medio silvestre (E), En peligro de extinción (PE), Amenazadas (AZ), Sujetas a protección especial (PR) y Sin Categoría (sc). Estado de conservación UICN (2019): Extinto (EX), En peligro (EN), Vulnerable (VU), Casi amenazado (NT), Preocupación menor (LC), Datos insuficientes (DD). Origen y categoría de endemismo (EN/EX): Endémica (EN), Cuasiendémica (CE), Semiendémica (SE), Exótica (Exo) y No endémica (ne). Gremio trófico (Gt), Residencia (R).
| Orden | Familia | Nombre científico | Nombre español | Gt | R | NOM-059 | UICN | EN/EX |
|---|---|---|---|---|---|---|---|---|
| Tinamiformes | Tinamidae | Crypturellus cinnamomeus | Tinamú Canelo | Fr | R | Pr | LC | ne |
| Anseriformes | Anatidae | Dendrocygna autumnalis | Pijije Alas Blancas | Gr | R | sc | LC | ne |
| Galliformes | Odontophoridae | Colinus nigrogularis | Cordoniz Yucateca | Gr | R | sc | LC | CE |
| Galliformes | Cracidae | Crax rubra | Hocofaisán | Fr | R | A | VU | ne |
| Galliformes | Cracidae | Ortalis vetula | Chachalaca Oriental | Fr | R | sc | LC | ne |
| Suliformes | Phalacrocoracidae | Phalacrocorax brasilianus | Cormorán Oliváceo | Pi | R | sc | LC | ne |
| Pelecaniformes | Ardeidae | Butorides virescens | Garcita Verde | Pi | R/MI | sc | LC | ne |
| Pelecaniformes | Ardeidae | Egretta caerulea | Garza Azul | Pi | MI/R | sc | LC | ne |
| Pelecaniformes | Ardeidae | Egretta tricolor | Garza Tricolor | Pi | MI/R | sc | LC | ne |
| Pelecaniformes | Pelecanidae | Tigrisoma mexicanum | Garza Tigre Mexicana | Pi | R | Pr | LC | ne |
| Accipitriformes | Accipitridae | Buteo plagiatus | Aguililla Gris | Ca | R | sc | LC | ne |
| Accipitriformes | Accipitridae | Buteogallus urubitinga | Aguililla Negra Mayor | Ca | R | Pr | LC | ne |
| Accipitriformes | Cathartidae | Cathartes aura | Zopilote Aura | Carr | R | sc | LC | ne |
| Accipitriformes | Cathartidae | Coragyps atratus | Zopilote Común | Carr | R | sc | LC | ne |
| Accipitriformes | Accipitridae | Elanus leucurus | Milano Cola Blanca | Ca | R | sc | LC | ne |
| Accipitriformes | Accipitridae | Geranoaetus albicaudatus | Aguililla Cola Blanca | Ca | R | Pr | LC | ne |
| Accipitriformes | Accipitridae | Rupornis magnirostris | Aguililla Caminera | In | R | sc | LC | ne |
| Accipitriformes | Cathartidae | Sarcoramphus papa | Zopilote Rey | Carr | R | P | LC | ne |
| Gruiformes | Rallidae | Aramides cajaneus | Rascón Cuello Gris | Om | R | sc | LC | ne |
| Gruiformes | Aramidae | Aramus guarauna | Carrao | Ca | R | A | LC | ne |
| Charadriiformes | Scolopacidae | Actitis macularius | Playero Alzacolita | In | MI | sc | LC | ne |
| Charadriiformes | Scolopacidae | Calidris mauri | Playero Occidental | In | MI/T | sc | LC | ne |
| Charadriiformes | Jacanidae | Jacana spinosa | Jacana Norteña | In | R | sc | LC | ne |
| Columbiformes | Columbidae | Claravis pretiosa | Tórtola Azul | Gr | R | sc | LC | ne |
| Columbiformes | Columbidae | Columbina talpacoti | Tortolita Canela | Gr | R | sc | LC | ne |
| Columbiformes | Columbidae | Geotrygon montana | Paloma perdiz común | Gr | R | sc | LC | ne |
| Columbiformes | Columbidae | Leptotila verreauxi | Paloma Arroyera | Gr | R | sc | LC | ne |
| Columbiformes | Columbidae | Patagioenas flavirostris | Paloma Morada | Fr | R | sc | LC | ne |
| Columbiformes | Columbidae | Patagioenas speciosa | Paloma Escamosa | Fr | R | Pr | LC | ne |
| Cuculiformes | Cuculidae | Crotophaga sulcirostris | Garrapatero Pijuy | In | R | sc | LC | ne |
| Cuculiformes | Cuculidae | Piaya cayana | Cuclillo Canela | In | R | sc | LC | ne |
| Apodiformes | Trochilidae | Amazilia candida | Colibrí Cándido | Ne | R/MI | sc | LC | ne |
| Apodiformes | Trochilidae | Amazilia yucatanensis | Colibrí Yucateco | Ne | R | sc | LC | CE |
| Trogoniformes | Trogonidae | Trogon melanocephalus | Coa Cabeza Negra | Fr | R | sc | LC | ne |
| Coraciiformes | Alcedinidae | Chloroceryle americana | Martín Pescador Verde | Ca | R | sc | LC | ne |
| Coraciiformes | Alcedinidae | Megaceryle torquata | Martín Pescador de Collar | Ca | R | sc | LC | ne |
| Coraciiformes | Momotidae | Momotus coeruliceps | Momoto Corona Azul | In | R | sc | LC | EN |
| Piciformes | Picidae | Campephilus guatemalensis | Carpintero Pico Plata | In | R | Pr | LC | ne |
| Piciformes | Picidae | Dryocopus lineatus | Carpintero Lineado | In | R | sc | LC | ne |
| Piciformes | Picidae | Melanerpes aurifrons | Carpintero Cheje | In | R | sc | LC | ne |
| Piciformes | Ramphastidae | Pteroglossus torquatus | Tucancillo Collarejo | Fr | R | Pr | LC | ne |
| Piciformes | Ramphastidae | Ramphastos sulfuratus | Tucán Pico Canoa | Fr | R | A | LC | ne |
| Falconiformes | Falconidae | Falco rufigularis | Halcón Murcielaguero | Ca | R | sc | LC | ne |
| Falconiformes | Falconidae | Herpetotheres cachinnans | Halcón Guaco | Ca | R | sc | LC | ne |
| Psittaciformes | Psittacidae | Amazona albifrons | Loro Frente Blanca | Fr | R | Pr | LC | ne |
| Psittaciformes | Psittacidae | Amazona autumnalis | Loro Cachetes Amarillos | Fr | R | sc | LC | ne |
| Psittaciformes | Psittacidae | Eupsittula nana | Perico Pecho Sucio | Fr | R | Pr | LC | ne |
| Passeriformes | Icteridae | Agelaius phoeniceus | Tordo Sargento | In | R/MI | sc | LC | ne |
| Passeriformes | Icteridae | Amblycercus holosericeus | Cacique Pico Claro | Om | R | sc | LC | ne |
| Passeriformes | Emberizidae | Arremonops chloronotus | Rascador Dorso Verde | Gr | R | sc | LC | ne |
| Passeriformes | Tyrannidae | Attila spadiceus | Mosquero Atila | In | R | sc | LC | ne |
| Passeriformes | Parulidae | Basileuterus culicivorus | Chipe Cejas Negras | In | R | sc | LC | ne |
| Passeriformes | Cardinalidae | Cardinalis cardinalis | Cardenal Rojo | Gr | R | sc | LC | ne |
| Passeriformes | Tyrannidae | Contopus cinereus | Papamoscas Tropical | In | R | sc | LC | ne |
| Passeriformes | Thraupidae | Cyanerpes cyaneus | Mielero Patas Rojas | Ne | R | sc | LC | ne |
| Passeriformes | Corvidae | Cyanocorax yncas | Chara Verde | Om | R | sc | LC | ne |
| Passeriformes | Corvidae | Cyanocorax yucatanicus | Chara Yucateca | Om | R | sc | LC | CE |
| Passeriformes | Vireonidae | Cyclarhis gujanensis | Vireón Cejas Canelas | In | R | sc | LC | ne |
| Passeriformes | Icteridae | Dives dives | Tordo Cantor | In | R | sc | LC | ne |
| Passeriformes | Fringiilidae | Euphonia hirundinacea | Eufonia Garganta Amarilla | Fr | R | sc | LC | ne |
| Passeriformes | Parulidae | Geothlypis trichas | Mascarita Común | In | MI/R | sc | LC | ne |
| Passeriformes | Cardinalidae | Habia fuscicauda | Piranga Hormiguera Garganta Roja | In | R | sc | LC | ne |
| Passeriformes | Hirundinidae | Hirundo rustica | Golondrina Tijereta | In | MV, MI, R, T | sc | LC | ne |
| Passeriformes | Turdidae | Hylocichla mustelina | Zorzal Moteado | In | MI | sc | NT | ne |
| Passeriformes | Icteridae | Icterus chrysater | Calandria Dorso Amarillo | In | R | sc | LC | ne |
| Passeriformes | Icteridae | Icterus cucullatus | Calandria Dorso Negro Menor | In | MI, MV, R | sc | LC | SE |
| Passeriformes | Icteridae | Icterus gularis | Calandria Dorso Negro Mayor | In | R | sc | LC | ne |
| Passeriformes | Icteridae | Icterus mesomelas | Calandria Cola Amarilla | In | R | sc | LC | ne |
| Passeriformes | Icteridae | Icterus prosthemelas | Calandria Caperuza Negra | In | R | sc | LC | ne |
| Passeriformes | Mimidae | Mimus gilvus | Centzontle Tropical | In | R | sc | LC | ne |
| Passeriformes | Parulidae | Mniotilta varia | Chipe Trepador | In | MI | sc | LC | ne |
| Passeriformes | Tyrannidae | Myiarchus yucatanensis | Papamoscas Yucateco | In | R | sc | LC | CE |
| Passeriformes | Tyrannidae | Myiodynastes maculatus | Papamoscas Rayado Cheje | In | MV | sc | LC | ne |
| Passeriformes | Tyrannidae | Myiozetetes similis | Luisito Común | In | R | sc | LC | ne |
| Passeriformes | Tyrannidae | Oncostoma cinereigulare | Mosquerito Pico Curvo | In | R | sc | LC | ne |
| Passeriformes | Tyrannidae | Onychorhynchus coronatus | Mosquero Real | In | R | P | LC | ne |
| Passeriformes | Tityridae | Pachyramphus aglaiae | Cabezón Degollado | In | R | sc | LC | ne |
| Passeriformes | Parulidae | Parkesia motacilla | Chipe Arroyero | In | MI | sc | LC | ne |
| Passeriformes | Troglodytidae | Pheugopedius maculipectus | Saltapared Moteado | In | R | sc | LC | ne |
| Passeriformes | Tyrannidae | Pitangus sulphuratus | Luis Bienteveo | In | R | sc | LC | ne |
| Passeriformes | Tyrannidae | Platyrinchus cancrominus | Mosquerito Pico Chato | In | R | Pr | LC | ne |
| Passeriformes | Hirundinidae | Progne chalybea | Golondrina Pecho Gris | In | R, MV | sc | LC | ne |
| Passeriformes | Corvidae | Psilorhinus morio | Chara Pea | Om | R | sc | LC | ne |
| Passeriformes | Thraupidae | Saltator atriceps | Saltador Cabeza Negra | Fr | R | sc | LC | ne |
| Passeriformes | Thraupidae | Saltator coerulescens | Saltador Chucho Páez | Fr | R | sc | LC | ne |
| Passeriformes | Parulidae | Seiurus aurocapilla | Chipe Suelero | In | MI | sc | LC | ne |
| Passeriformes | Furnaridae | Sittasomus griseicapillus | Trepatroncos Cabeza Gris | In | R | sc | LC | ne |
| Passeriformes | Emberizidae | Sporophila torqueola | Semillero de Collar | Gr | R | sc | LC | EN |
| Passeriformes | Hirundinidae | Stelgidopteryx serripennis | Golondrina Yucateca | In | R, MI | sc | LC | ne |
| Passeriformes | Furnaridae | Synallaxis erythrothorax | Hormiguero Pepito | In | R | sc | LC | ne |
| Passeriformes | Troglodytidae | Thryothorus ludovicianus | Saltapared de Carolina | In | R | sc | LC | ne |
| Passeriformes | Emberizidae | Tiaris olivaceus | Semillero Oliváceo | Gr | R | sc | LC | ne |
| Passeriformes | Tityridae | Tityra semifasciata | Titira Puerquito | In | R | sc | LC | ne |
| Passeriformes | Troglodytidae | Troglodytes aedon | Saltapared Común | In | R, MI, T | sc | LC | ne |
| Passeriformes | Turdidae | Turdus grayi | Mirlo Café | In | R | sc | LC | ne |
| Passeriformes | Tyrannidae | Tyrannus melancholicus | Tirano Pirirí | In | R | sc | LC | ne |
| Passeriformes | Tyrannidae | Tyrannus tyrannus | Tirano Dorso Negro | In | T | sc | LC | ne |
| Passeriformes | Troglodytidae | Uropsila leucogastra | Saltapared Vientre Blanco | In | R | sc | LC | CE |
| Passeriformes | Parulidae | Vermivora cyanoptera | Chipe Alas Azules | In | MI | sc | LC | ne |
| Passeriformes | Vireonidae | Vireo magister | Vireo Yucateco | In | R | sc | LC | CE |
| Passeriformes | Emberizidae | Volatinia jacarina | Semillero Brincador | Gr | R | sc | LC | ne |
| Passeriformes | Furnaridae | Xiphorhynchus flavigaster | Trepatroncos Bigotudo | In | R | sc | LC | ne |
| Passeriformes | Icteridae | Icterus spurius | Calandria Castaño | In | MI, MV | sc | LC | ne |
Estacionalidad
Estacionalmente, de las 103 especies registradas, 82 fueron residentes, siete migratorias y una transitoria. Respecto a los endemismos, encontramos que seis especies corresponden a la categoría de cuasiendémica, una semiendémica y una endémica (Tabla 2).
Vulnerabilidad
Por otro lado, el hocofaisán y el zorzal moteado (Hylocichla mustelina) se encuentran en la categoría de Vulnerable y Casi amenazada, respectivamente (UICN, 2019); mientras que en la NOM-059-SEMARNAT-2010, 10 especies se encuentran en la categoría de Sujeta a protección especial, tres amenazadas y dos en Peligro de extinción: el mosquero real (Onychorhynchus coronatus) y el zopilote rey (Sarcoramphus papa; Tabla 2).
DISCUSIÓN
Las estimaciones de diversidad y composición de las especies en distintos tipos de hábitat o gradientes de perturbación permiten entender de qué manera se estructuran las comunidades. En este estudio obtuvimos el 21 % del total de especies de aves reportadas para la Península de Yucatán, y el 53 % de las familias reportadas para el estado (Correa-Sandoval & MacKinnon, 2011). El esfuerzo de muestreo tuvo una buena cobertura de la muestra debido a que también se tomaron en cuenta los registros auditivos, pues se ha reportado que permiten detectar a las especies raras, poco conspicuas o que son muy activas en el dosel y se dificulta su registro visual (Celis-Murillo et al., 2012; Suárez-García et al., 2017; Espinoza-Méndez et al., 2020). De las aves terrestres de Quintana Roo, las familias Tyrannidae y Parulidae han sido reportadas con mayor número de especies (Correa-Sandoval & MacKinnon, 2011); en el presente estudio Tyrannidae fue más representada que Parulidae, con 11 y seis especies, respectivamente.
En cuanto a la riqueza, Laguna y Carretera presentaron el mayor número de especies. En contraste, Potrero fue el de menor riqueza específica, lo cual es similar a lo obtenido por González-Valdivia et al. (2012), quienes encontraron una menor riqueza en el área de potreros en comparación con las áreas mejor conservadas. El hecho de que no se encontrara mayor riqueza en Selva, que es el hábitat que se encuentra en mejor estado de conservación junto con Laguna, puede deberse a diversos factores, entre ellos, a la densa cobertura vegetal que caracteriza a las selvas de la Península de Yucatán, a la experiencia del observador y el tipo de muestreo (Espinoza-Méndez et al., 2020). Así mismo, se ha reportado que en follajes densos la detección de aves se torna más difícil, esto aunado al hecho de que en las fechas en las que se realizó el muestreo (agosto-septiembre) las aves son menos activas vocalmente, principalmente las residentes (González-Ortega & Morales-Pérez, 1998).
Otros autores como Schondube et al. (2018) reportaron que los hábitats con monocultivos o asentamientos urbanos mostraron valores más altos de riqueza y abundancia, a pesar de que los hábitats urbanizados son estructuralmente muy distintos al hábitat original. Esto se explica por dos razones: 1) la intensidad de la perturbación no es el principal factor que determina la composición y estructura de las comunidades de aves; y/o 2) los elementos estructurales de las zonas urbanizadas pueden permitir la entrada de especies que no se encuentran en ambientes muy perturbados, es decir, que pueden funcionar como sustitutos de los hábitats nativos (MacGregor-Fors & Schondube, 2011). En este estudio, Carretera presentó el mismo valor de riqueza que Laguna, a pesar de ser un ambiente menos conservado, esto puede ser a causa del efecto de borde que, al cambiar la estructura de la vegetación, modifica la distribución y abundancia de las especies y, por tanto, la oferta de alimento para la fauna. Estos cambios en la estructura de la vegetación de los márgenes pueden favorecer la aparición de un mayor número de especies, que generalmente tienden a ser especies adaptadas a espacios abiertos y que encuentran este tipo de ambientes más favorables para su supervivencia y reproducción, a pesar de que la perturbación pueda parecer más evidente. Por su parte, la composición de especies fue notoriamente distinta entre Carretera y Laguna, mientras que el Helechal con Carretera compartieron la mayoría de sus especies. Esto puede explicarse por factores extrínsecos a las aves, como la disponibilidad de recursos, las características estructurales del hábitat que, aunque estén modificados, puede resultar beneficioso para diversas especies, y las condiciones fisiográficas y climáticas de la región (Hutto, 1985; Carrara et al., 2015).
Los datos de riqueza y abundancia de especies de aves por sí solos no reflejan la intensidad de la perturbación en el paisaje. Sin embargo, las curvas de rango/abundancia nos permitieron visualizar la respuesta de las comunidades de aves a las modificaciones en el ambiente (MacGregor-Fors & Schondube, 2012; Maya-Elizarrás & Schondube, 2015). De esta manera, las especies más abundantes dentro del estudio fueron el tirano dorso negro seguido del loro cachete amarillo, del perico pecho sucio y del luis gregario, especies que generalmente están asociadas a hábitats abiertos, suburbanos o cultivos; es decir, hábitats que ya presentan cierto grado de perturbación.
De los cinco tipos de hábitat muestreados en este estudio, la pendiente más pronunciada se reflejó en Potrero, Helechal y Carretera, que son precisamente los hábitats que se consideran más perturbados, y donde las especies dominantes fueron de la familia Tyrannidae, que se benefician de los ambientes abiertos (Fagan & Komar, 2016). Por el contrario, en Laguna y Selva, las pendientes se mostraron menos pronunciadas, lo que indica que el estado de conservación del área es mejor y que la repartición de recursos es más equitativa (Whittaker, 1965; Magurran, 2004; Moreno et al., 2018). Así también, las rapaces y carroñeros dominaron en las áreas abiertas como Potrero, Helechal y Carretera, similarmente a lo reportado por González-Valdivia et al. (2012), ya que en áreas abiertas es más fácil cazar y obtener su alimento. Así mismo, estas curvas concuerdan con las predicciones generales sobre el poco efecto de las perturbaciones en la riqueza y uniformidad de la abundancia de las aves (evenness) mientras existan recursos alimenticios, y sobre la dominancia de algunas pocas especies en los sitios perturbados o abiertos (Bezzel, 1985; Jokimäki et al., 1996; Tu et al., 2020).
Por su parte, la gran similitud en la composición de especies de Laguna y Selva, en comparación con Potrero, sugiere que las especies que se encuentran en los lugares más conservados en este estudio como el mosquero real, el colibrí cándido (Amazilia candida), el hocofaisán, y la paloma canela (Geotrygon montana), no tienden a desplazarse a los ambientes más perturbados, pues son menos tolerantes a la perturbación o son exclusivas de cierto tipo de hábitat (Sekercioglu et al., 2002).
La composición de especies en los distintos tipos de hábitat en el presente estudio corresponde al grado de perturbación del hábitat en el que se encuentran. González-Valdivia et al. (2012) reportaron que en los potreros y áreas abiertas predominaron las especies de hábitos insectívoros, lo cual coincide con este estudio, donde el tirano dorso negro presentó más abundancia en Potrero y Helechal que es un área abierta, al igual que los granívoros como el semillero rabadilla canela (S. torqueola) y el semillero brincador (Volatinia jacarina). Esto se debe a que las áreas abiertas o agroecosistemas como el hábitat Potrero favorecen la presencia de algunas aves como el tordo sargento (Agelaius phoeniceus), el garrapatero pijuy (C. sulcirostris), el tordo ojos rojos (Molotrus aeneus), el zanate mayor (Quiscalus mexicanus), el jilguerito dominico (Spinus psaltria), el semillero brincador (V. jacarina) y la paloma alas blancas (Zenaida asiatica), que son generalistas y están relacionadas a los ambientes antropizados, esto coincide con lo reportado por Álvarez-Álvarez et al. (2018).
En este estudio, el gremio de los insectívoros mostró mayor riqueza en Laguna y Carretera, similar a lo obtenido por Espinoza-Méndez et al. (2020), quienes encontraron especies insectívoras abundantes en zonas perturbadas, como Carretera y en zonas más conservadas, como el caso de Laguna para este estudio. Esta complementariedad de la riqueza de especies entre hábitats conservados (Laguna y Selva) y perturbados (Carretera y Helechal) puede ser una evidencia del efecto del disturbio intermedio que propuso Connell (1978). Este autor propone que los ecosistemas sujetos a perturbaciones con frecuencia moderada permiten una mayor diversidad, siempre y cuando las poblaciones de especies puedan recuperarse entre eventos de disturbio. Por otro lado, las aves insectívoras son sensibles y vulnerables a las perturbaciones, y su persistencia depende de la capacidad de desplazarse entre ambientes perturbados, si las aves no se desplazan los suficiente o son exclusivas de algún tipo de hábitat se ven más propensas a desaparecer (Sekercioglu et al., 2002).
Los gremios frugívoro, insectívoro y granívoro detectados en nuestro estudio, fueron representativos como los encontrados en otros estudios (Rangel-Salazar et al., 2009: Espinoza-Méndez et al., 2020). Estos gremios son de suma importancia, ya que se encargan de la dispersión de semillas y el control de plagas, además de que se asocian a los ambientes con mayor complejidad estructural y florística (Ramírez-Albores, 2006). No se puede dejar de lado que los gremios se ven afectados en su preferencia por sitios debido a las condiciones ambientales (e. g., temperatura, humedad relativa y altitud), así como por variables estructurales de la vegetación como la cobertura o altura del dosel (González-Valdivia et al., 2012).
En Quintana Roo habitan especies que no se encuentran en otra parte de México, un ejemplo es el vireo yucateco (Vireo magister), el cual fue registrado en este estudio y es endémico del estado. Así mismo, aves migratorias terrestres y acuáticas utilizan la costa y las islas para su viaje al sur del continente americano. Las parvadas del tirano dorso negro (que fue la especie más abundante en este estudio) migran a Sudamérica y en su paso se alimentan del chechem negro (Metopium brownei; Jacq. Urban, 1908) que domina en la vegetación de la costa (Correa-Sandoval & MacKinnon, 2011). De esta manera, las selvas de Quintana Roo proveen las condiciones para aves que no se encuentran en otros estados y es propicio para que arriben especies migratorias.
La pérdida del hábitat es la mayor amenaza para la avifauna, en Quintana Roo se debe principalmente a la expansión de la mancha urbana, los monocultivos y la ganadería extensiva. Además, el cambio de uso de suelo para el desarrollo turístico constantemente está creciendo, desplazando así a toda la fauna que habita la vegetación nativa (Correa-Sandoval & MacKinnon, 2011). En este sentido, hay especies más vulnerables a dichos cambios en el paisaje, por lo que actualmente se encuentran en alguna categoría de riesgo de acuerdo con la norma oficial mexicana (NOM-059-SEMARNAT-2010) o en la UICN (2019). Tal es el caso del hocofaisán, que está presente en el ejido Laguna Om y se encuentra en la categoría de amenazada (NOM-059-SEMARNAT-2010) y vulnerable (UICN, 2019). Por otra parte, el zorzal moteado se encuentra casi amenazado según la UICN (2019). Es importante destacar que la presencia del hocofaisán en el hábitat de Selva sugiere que este sitio se encuentra en buen estado de conservación, ya que esta especie es susceptible a la cacería y a las perturbaciones (Ramírez-Barajas et al., 2012).
Se puede concluir en este estudio que las diferencias en la composición de las especies se deben a distintos factores como: las modificaciones del paisaje por actividades agrícolas, forestales o ganaderas, por los asentamientos urbanos y la construcción de carreteras y caminos para el desplazamiento humano, que a su vez provocan cambios en la estructura de la vegetación. También es importante considerar que la introducción de especies exóticas es otro factor que desplaza a las especies nativas, la captura y comercio ilegal, que es más común para los loros y aves canoras. Así mismo, las variables físicas y ambientales o de la estructura de la vegetación juegan un papel vital para que los gremios tróficos se vean afectados en su preferencia de hábitat. No obstante, los ambientes con cierto grado de perturbación pueden proveer las condiciones óptimas para una amplia diversidad de especies tolerantes a perturbaciones antrópicas, como el caso de Carretera en este estudio.
Por otro lado, existen especies indicadoras de la calidad del hábitat, como el mosquero real o el hocofaisán presentes en nuestro estudio, que permiten diferenciar la condición de cada tipo de hábitat. De esta manera podemos concluir que, si una especie representativa de áreas conservadas no se encuentra en sitios con cierto grado de perturbación, entonces se vuelven especies útiles para monitorear el estado de conservación del ejido forestal de Laguna Om. Por otro lado, especies como el tirano dorso negro, el garrapatero pijuy, el zanate mayor y la paloma alas blancas pueden ser indicadoras del estado de perturbación de áreas abiertas.
También es importante destacar que hay que encaminar esfuerzos para las actividades de conservación para mantener la comunidad de aves diversa y funcional, aún en ambientes heterogéneos como el ejido Laguna Om. Una manera de lograrlo es valorar el papel de la vegetación en el mantenimiento de la diversidad de la avifauna, principalmente cuando las perturbaciones antrópicas reducen los recursos alimenticios y la cobertura y estructura arbórea.
Finalmente, conocer la respuesta de las aves ante los distintos tipos de perturbación es crucial para generar planes de manejo, restauración y conservación, sobre todo en áreas consideradas como corredores biológicos como en el presente estudio que conecta las dos grandes reservas de la biosfera de la Península de Yucatán: Sian Ka’an y Calakmul.

