











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
México presenta una elevada diversidad de aves, ya que aproximadamente se distribuyen 1,115 especies que representan el 10% de la diversidad mundial (Berlanga et al., 2017). La península de Yucatán y la vertiente del Pacífico son las áreas dónde se concentra una mayor riqueza (Navarro et al., 2014). La mayoría de las aves son de hábitos terrestres y cumplen con distintas funciones ecológicas como la dispersión de semillas y polinización, por esta razón son consideradas como “ingenieros de los ecosistemas” (Sekercioglu, 2006); son los vertebrados terrestres con la mayor capacidad de movimiento y que pueden cubrir amplias distancias durante diferentes etapas de su historia de vida (Cox, 2010). Para muchas especies la migración es parte de su historia de vida y es considerado un proceso ecológico-evolutivo (Newton, 2008), en el que las aves tienen una respuesta adaptativa a la heterogeneidad espacial y temporal en los recursos.
México ha sufrido una de las mayores tasas de modificación de sus ecosistemas y deforestación a nivel de América Latina, tan solo en las últimas dos décadas más de 100,000 ha de bosques han sido deforestados en México (Ellis et al., 2017). Entre 1993 y 2002 Campeche tenía una tasa de deforestación de 30,968 ha por año (Céspedes & Moreno, 2010), causada por la intensificación de actividades agropecuarias (Smith et al., 2001). En Campeche el 74% del territorio estatal corresponde a terrenos que han sido modificados por actividades humanas y, solamente el 25.1% restante está cubierto por vegetación natural (INEGI, 2015). En la región de Calakmul, Campeche, el sistema de propiedad ejidal ha sido el principal promotor del cambio de uso de suelo en la región (Chowdhury, 2006). El resultado es un paisaje manejado conteniendo un mosaico de distintos tipos de vegetación con diferentes etapas de sucesión (Warkentin et al., 1995; Smith et al., 2001). Un ejemplo de esto es la presencia de acahuales o hubchés (Martínez & Galindo, 2002), que son comunidades secundarias de la selva baja o mediana asociadas a la producción agrícola de la roza-tumba y quema (Levy & Aguirre, 2000). Algunas comunidades de la región reconocen al menos cuatro estadios dependiendo de su edad, en los cuáles la estructura y tipo de vegetación varía dependiendo del tiempo de abandono (Granados et al., 1999). Cuando estas prácticas se realizan a pequeña escala y quedan parches intactos de vegetación se puede promover el mantenimiento de la biodiversidad (Smith et al., 2001). En este sentido, se ha considerado que los disturbios intermedios producen un mosaico de hábitats que, en paisajes transformados, pueden tener una diversidad de especies mayor. Esta hipótesis considera que en pequeños disturbios la exclusión competitiva produce una menor diversidad, mientras que, en disturbios grandes, las especies suelen ser extirpadas por no poder colonizar nuevos espacios. Otra condición que se menciona facilita este incremento en la diversidad es la heterogeneidad en el paisaje, siendo de alguna manera importante para la supervivencia de distintas comunidades animales (Fahrig, 2003, 2017; Connell, 1978) entre ellas, las aves. Sin embargo, esta hipótesis ha sido debatida ya que son pocos los estudios que han demostrado que los valores más altos de diversidad se presenten en niveles intermedios de disturbio (Fox, 2013). La vegetación presente en los sitios como resultado del uso de suelo, puede relacionarse con la presencia de distintos grupos de aves, como es el caso del uso de los cafetales por aves migratorias, o los paisajes agroforestales en los que las aves utilizan preferentemente el dosel (Greenberg et al., 1997; Van et al., 2008).
Es importante identificar la influencia del uso del suelo sobre las comunidades animales, ya que puede ayudar a identificar zonas necesarias para la conservación de especies residentes y migratorias. El objetivo general de este estudio fue analizar la influencia del uso de suelo sobre las comunidades de aves en el ejido Nuevo Conhuas, en la región de Calakmul. Los objetivos particulares fueron: determinar la cobertura de suelo de cada sitio, como reflejo de su uso; analizar la diversidad de la comunidad de aves en cada sitio con distinto uso de suelo; analizar la influencia del uso de suelo sobre la abundancia de especies entre sitios y mes de muestreo, así como la abundancia de las aves migratorias neotropicales y residentes; analizar la influencia del uso de suelo sobre gremios alimenticios; identificar aquellas especies que se obtuvieron en una sola ocasión o aquellas obtenidas en dos ocasiones a lo largo del muestreo; e identificar la importancia de cada uso de suelo sobre distintas especies y familias de aves.
Materiales y métodos
El ejido Nuevo Conhuas se localiza al sureste del estado de Campeche, (89° 55' 22" N, 18° 32' 26 " O) en el municipio de Calakmul, y forma parte de la Reserva de la Biósfera de Calakmul, la cual limita al este con el estado de Quintana Roo y al sur con la República de Guatemala (Fig. 1). De acuerdo con el sistema de clasificación de Köppen modificado por García (1973), el clima de la región es cálido subhúmedo (Aw) (García, 1973; SEMARNAP, 1999), la temperatura promedio anual es de 24.6°C, y las precipitaciones pueden alcanzar hasta 873.3 mm (SEMARNAP, 1999). La vegetación principal son selvas secas inmersas en una mezcla de selvas tropicales, subperennifolias y subcaducifolias secas pudiendo ser altas, medianas, bajas o inundables (Miranda, 1958; SEMARNAP, 1999).

Selección de sitios de muestreo. El muestreo fue realizado en el ejido Nuevo Conhuas (18° 32' 26 " N, 89° 55' 22" O) donde se seleccionaron cuatro sitios con base en el uso actual de suelo. Para esto, realizamos entrevistas semiestructuradas a tres de los pobladores más viejos del ejido, quienes fueron de los primeros habitantes del mismo y conocieron las condiciones originales de la región de estudio y los cambios que ha habido en el uso del suelo. El primer sitio se localizó dentro de la REBICA (18° 18' 52.19" N, 89° 51' 23.40" O), esta área está sujeta a protección y no se permite el aprovechamiento extractivo desde la declaración de la reserva en 1989.
La vegetación presente en este sitio es una mezcla de selva baja (también llamados bajos inundables), selva mediana y alta. Los otros sitios se ubicaron en la zona de amortiguamiento de la REBICA; en estos sitios se lleva a cabo aprovechamiento de recursos forestales y actividades como la agricultura y ganadería. El segundo sitio es un acahual de manejo forestal con más de 20 años de abandono (18° 31' 19.61" N, 89° 47' 31.92" O) en el que se realizó la explotación de los recursos forestales, la vegetación es selva mediana. El tercer sitio es un acahual de agricultura con vegetación secundaria entre 5 y 10 años desde su abandono (18° 32' 9.24" N, 89° 53' 12.95" O), era sembrado con maíz, chile y calabaza (chihua). El tipo de agricultura que se realizó en este sitio fue roza, tumba y quema; la vegetación presente actualmente es selva baja y selva mediana. El cuarto sitio es de manejo ganadero (18° 31' 22.97" N, 89° 54' 47.53" O) en el que se mantiene ganado bovino en pastizales de Muhlenbergia spp. Es importante mencionar que los sitios en la zona de amortiguamiento se encuentran inmersos en parches en los que se han realizado distintas actividades de aprovechamiento, tales como ganadería, agricultura, aprovechamiento forestal y apicultura, entre otras. Algunas actividades se siguen realizando, como la agricultura a pequeña escala y apicultura.
Muestreo de aves. El muestreo de aves se realizó del 01 de febrero de 2017 al 08 de junio de 2017. Se colocaron seis redes de niebla en el sotobosque, tres de seis m y tres de 12 m con una separación de 90 y 120 m entre ellas (Ralph et al., 2001), las cuales permanecieron abiertas desde las 06:30 hr hasta las 11:00 hr durante tres días consecutivos en las fechas anteriormente mencionadas. Este procedimiento se realizó cinco veces en cada sitio con un intervalo de tiempo de tres semanas. El esfuerzo total de muestreo fue de 14,580 hr/red, las aves fueron identificadas a nivel de especie con guías de campo (Kaufman, 2005; Perlo, 2006; MacKinnon, 2013; Arizmendi & Berlanga, 2014; Sibley, 2014). Las aves fueron marcadas en ambos tarsos con barniz para uñas (Kendall et al., 2009) para identificar posibles recapturas. Posterior a esto las aves fueron liberadas en el lugar de captura.
Análisis del paisaje. Para determinar la cobertura del suelo de cada sitio se utilizó ArcGIS versión 10.2. y FRAGSTATS versión 4 (McGarigal et al., 2012). Fue utilizada una imagen satelital LANDSAT 8 OLI del 14 de febrero de 2017 (USGS, 2017); debido a que no se requería hacer la comparación en el tiempo de las coberturas y usos de suelo de la zona, no se realizó una corrección atmosférica y la imagen se trabajó utilizando los números digitales. Se llevó a cabo una clasificación automatizada no supervisada determinando cuatro clases para toda la imagen (con base en el conocimiento de la región de estudio por el primer autor). Las clases fueron clasificadas de la siguiente manera: vegetación conservada (VEC), acahuales maduros (ACM), vegetación secundaria y pastizales (VESPA), y caminos y construcciones (SINV). Fue calculada también la riqueza y número de parches, así como la densidad de borde de los sitios de estudio con FRAGSTATS v4 (McGarigal et al., 2012). Se incluyó también la distancia a la comunidad en kilómetros (DISC) como variable explicativa.
Se estableció una zona buffer de 70 ha a partir del centroide de cada sitio en la que fueron colocadas las redes, la cual fue determinada con base en datos de rango hogareño de tres especies capturadas: Hylocichla mustelina (Gmelin, 1789), Sittasomus griseicapillus (Vieillot, 1818) y Empidonax virescens (Vieillot, 1819). Se ha reportado que los ámbitos hogareños registrados para estas especies no exceden una superficie de 70 ha (Anders et al., 1998; Ausprey & Rodewald, 2013; Jirinec et al., 2016).
Análisis de datos. La diversidad de aves fue analizada con el programa estadístico R Studio 1.0.153 (RStudio Team, 2016) utilizando el Paquete INEXT para conocer los números efectivos de especies (Chao et al., 2014) en sus órdenes (q) 0, 1 y 2, con el fin de identificar las diferencias en la diversidad de especies en cada condición de uso de suelo. Los números efectivos de especies permiten comparar la magnitud de las diferencias en las comunidades (Moreno et al., 2010). Fue calculado el valor de equidad EF1,2= D2/D1 (EF= equidad; D2= orden dos de diversidad; D1= Orden uno de diversidad) con los números efectivos de especies; esta relación permite representar la proporción de especies dominantes de una comunidad (Jost, 2010). Se calculó la cobertura de muestra para la comunidad de aves, para realizar una comparación más acertada en relación con los valores de diversidad obtenidos por medio de los números efectivos de especies (Jost, 2010).
Fue calculado el índice de diversidad Beta de Whittaker (βw), el cual es una relación del promedio de las diversidades alfa y la diversidad gamma (Calderón et al., 2012), así como el índice de similitud de Jaccard para conocer la similitud de las comunidades en cada sitio de muestreo. Fueron utilizadas pruebas de Kruskal-Wallis con la corrección de Bonferroni en R Studio 1.0.153 para analizar las diferencias en la abundancia entre los sitios muestreados y meses de muestreo para aves migratorias neotropicales y aves residentes, así como entre los tres gremios alimenticios más abundantes de la comunidad (insectívoros, nectarívoros y semilleros). La determinación de los gremios fue realizada con información disponible en los sitios web Naturalista (htp://www.naturalista.mx), Guía en línea del Laboratorio de aves de Cornell (http://neotropical.birds.cornell.edu) y guía en línea de Audubon para aves de Norteamérica (http://www.audubon.org/bird-guide), así como información en Neri (2007) y la guía de aves de Sibley (2014). Durante el muestreo basado en la abundancia de las especies, y con el objetivo de tener una aproximación sobre la presencia de especies raras, identificamos aquellas especies únicas o que se registraron en solo una unidad de muestreo (f1 o singletons) y aquellas especies que se capturaron en dos unidades muestrales durante todo el estudio (f2 o doubletons), con el programa estadístico EstimateS V.9 (Colwell, 2013).
Se realizó un análisis de correspondencia canónica (CCA) con el paquete Vegan (Oksanen et al., 2018) en R Studio, para determinar las asociaciones entre la comunidad de aves y las variables de tipos de vegetación de los sitios muestreados. Este método multivariado evalúa las relaciones de las especies con el ambiente, que están interactuando en un área determinada (Braak, 1987). Las variables utilizadas en el modelo fueron el logaritmo base 10 de la superficie de acahuales maduros (ACM), caminos y construcciones (SINV), y la distancia a la comunidad de Nuevo Conhuas (DISC). La abundancia de especies fue transformada al logaritmo natural más uno, la significancia del modelo y de las variables incluidas fueron evaluadas con pruebas ANOVA. Para la ordenación se consideró la escala de Hill tipo 2, (el promedio ponderado de las especies por sitio de registro). Esta escala permite interpretar espacial mente la relación entre las especies y las variables ambientales en los sitios donde éstas se registraron (McCune & Grace, 2002). Debido a que el CCA es sensible a la abundancia de las especies menos abundantes o raras, se realizó el tratamiento previo de los datos de abundancias de las especies de aves, transformando la variable considerando la distancia de Hellinger por ser recomendada para análisis de conglomerados o de ordenación cuando se analiza la composición de comunidades (Legendre & Gallagher, 2001).
Con las variables significativas del CCA se realizaron modelos lineales generalizados (GLM) con distribución binomial negativa para la comunidad de aves, con el objetivo de analizar la influencia del sitio y tipos de vegetación sobre la abundancia de las aves y familias; las familias representadas por un solo individuo fueron excluidas del modelo. Todos los modelos lineales generalizados se realizaron en R studio 1.0.153 con el paquete mvabund (Wang et al., 2012).
Resultados
La REBICA presentó la mayor superficie de vegetación conservada (52.56 ha) y la menor la tuvo el sitio ganadero (3.96 ha). Los sitios identificados como acahuales presentaron superficies similares de todos los tipos de vegetación. Fueron la REBICA y el sitio ganadero los que presentaron superficies de vegetación más contrastantes. El sitio dentro de la REBICA fue el más alejado de la comunidad, mientras que el sitio ganadero fue el más cercano (Fig. 2).
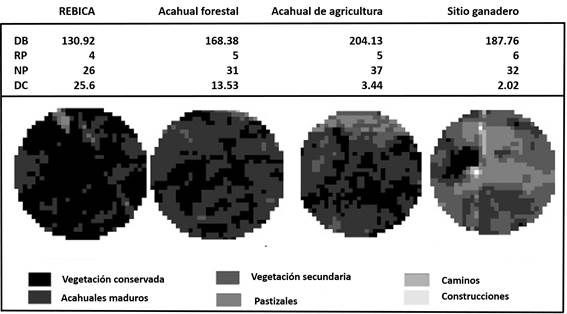
Figura 2 Resultados del análisis de paisaje y cobertura de suelo de cada sitio. Los círculos en escala de grises representan la superficie de cada sitio y cada tono es un tipo de cobertura. DB (Densidad de borde), RP (Riqueza de parches), NP (Número de parches), DC (Distancia a la comunidad en km).
Fueron capturados 415 individuos (nueve de ellos recapturas) representados por siete órdenes, 22 familias y 82 especies. El 42.61% de las aves capturadas fueron migratorias neotropicales mientras que el 55.41% fueron aves residentes; el porcentaje restante fueron aves transitorias. La clasificación taxonómica, estacional, y por gremio alimenticio se muestra en el Apéndice 1. El sitio con mayor riqueza de especies fue el ganadero (D0= 36) y su comunidad estuvo dominada por una menor cantidad de especies (D2= 3.88) La mayor diversidad observada estuvo en el acahual de agricultura (D1= 26.59), mientras que la mayor diversidad estimada la tuvo la REBICA (D1= 36.38). Los valores de riqueza (D0), diversidad (D1) y dominancia (D2) de los cuatro sitios de muestreo se muestran en la figura 3.
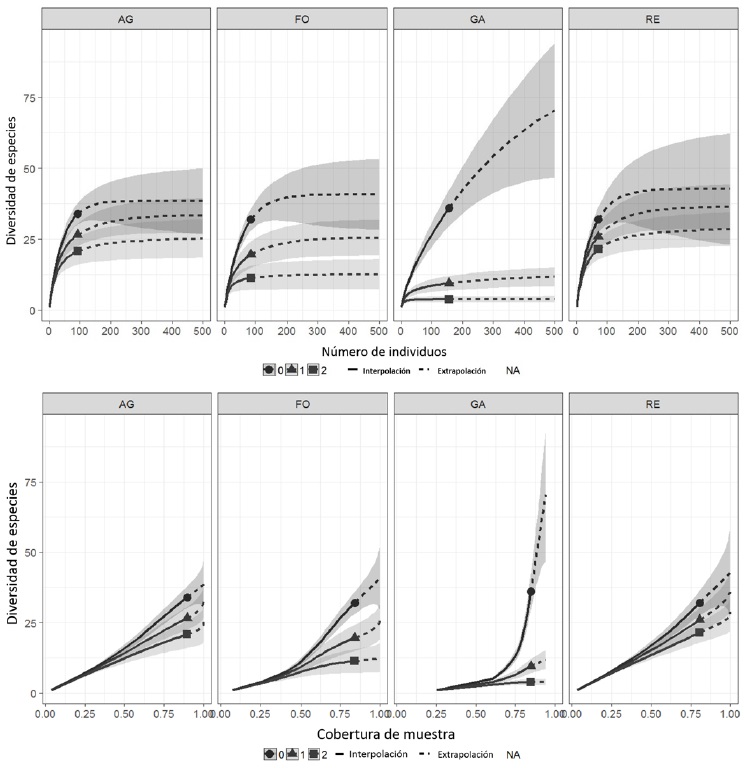
Figura 3 Riqueza (0), diversidad (1) y especies dominantes (2) en cada sitio de muestreo. La parte superior de la figura representa la diversidad de especies con relación al número de individuos capturados, mientras que la parte inferior representa la diversidad de especies con relación a la cobertura de muestra en cada sitio de muestreo. Se observa en la figura una diferencia en la magnitud de las comunidades de más del doble entre todos los sitios y el sitio ganadero, siendo los órdenes 0 y 1 (riqueza y diversidad, respectivamente) en los que se puede apreciar de manera más significativa esta diferencia, contrastantemente, se observa que la comunidad del sitio ganadero es dominada por una menor cantidad de especies (orden 2); AG (Acahual de agricultura); FO (Acahual forestal); GA (Sitio ganadero); RE (Reserva de la Biósfera de Calakmul).
Encontramos una mayor equidad en la comunidad de aves en la REBICA (0.82), seguida por el acahual de agricultura (0.78), el acahual forestal (0.59), y la menor la presentó el sitio ganadero (0.40). La diversidad beta de la comunidad fue βw= 1.44. Los sitios con mayor similitud respecto al índice de Jaccard fueron ambos acahuales (0.40) mientras que los menos similares fueron la REBICA y el sitio ganadero (0.15); la similitud fue relativamente media entre la REBICA y el acahual forestal (0.36) y de la REBICA con el acahual de agricultura (0.24).
La abundancia de las especies no presentó una distribución normal (W= 0.66, p<0.001). Se encontraron diferencias significativas en relación con la abundancia de aves migratorias neotropicales en los meses de enero y junio (Kruskal Wallis; X2= 13.88, df= 4, p= 0.007). También existieron diferencias entre los sitios con relación al gremio de los semilleros (X2= 15.22, df= 3, p= 0.001) entre el sitio ganadero y acahual forestal (p= 0.024), y sitio ganadero y la REBICA (p= 0.001); también entre los sitios con relación al gremio de los insectívoros (X2= 8.47, df= 3, p= 0.037) entre el acahual forestal y la REBICA. Fueron identificados 32 singletons, seis en la REBICA, cuatro en el acahual forestal, cinco en el acahual de agricultura y 17 en el sitio ganadero (Apéndice 1).
El CCA con las variables DISC+ACM+SINV presentó significancia estadística (p= 0.001) y explicó alrededor del 23% de la variación total. Las variables individualmente fueron también significativas, DISC (F= 1.74, p= 0.001), ACM (F= 1.57, p= 0.001) y SINV (F= 1.41, p= 0.019 (Fig. 4). Conforme a los GLM el uso de suelo de cada sitio influyó en la abundancia de las especies de aves (Cuadro 1).

Figura 4 Ordenación gráfica de los sitios muestreados. Se muestran las especies de aves más representativas en los sitios. Los acrónimos con números consecutivos del uno al cinco, pertenecen al número de repeticiones para cada una de las condiciones, en las cuales se puede apreciar la asociación de las especies a cada condición de uso; AG (Acahual de agricultura); FO (Acahual forestal); GA (Sitio ganadero); RE (Reserva de la Biósfera de Calakmul), DISC (Distancia en km a Nuevo Conhuas), SINV (Sin vegetación), ACM (Acahuales maduros).
Cuadro 1 Resultados de los coeficientes de los GLM para las especies de aves y las familias en los que se muestran los grados de libertad (Gl), Devianza (Dev), Significancia del modelo y de las variables utilizadas en cada modelo (p) y la suma de la información del criterio de Akaike (AIC) para cada modelo. Los valores de las variables de cada modelo fueron obtenidos por medio del análisis del paisaje. SP (Especies), VEC (Vegetación conservada), ACM (Acahuales maduros), SINV (Sin vegetación), FAM (Familias de aves).
| Modelo | Gl | Dev | p | AIC |
|---|---|---|---|---|
| GLM para especies de aves | ||||
| SP~VEC+ACM+SINV | 19 | 373.1 | 0.001 | 1923.124 |
| VEC | 18 | 186.20 | 0.002 | |
| ACM | 17 | 117.11 | 0.006 | |
| SINV | 16 | 69.83 | 0.01 | |
| GLM para familias de aves | ||||
| FAM~ VEC+ACM+SINV | 19 | 114.47 | 0.001 | 695.582 |
| VEC | 18 | 52.17 | 0.002 | |
| ACM | 17 | 40.46 | 0.005 | |
| SINV | 16 | 21.84 | 0.043 | |
Discusión
El uso de suelo tiene un efecto sobre la diversidad de las comunidades de aves, lo que fue evidente en los sitios muestreados de nuestro estudio. Nuestros resultados sugieren que los paisajes más heterogéneos y fragmentados favorecen la riqueza de aves, ya que el acahual de agricultura y el sitio ganadero tuvieron mayor heterogeneidad de cobertura, mayor fragmentación y presentaron una mayor riqueza de especies. La mayor riqueza de especies de aves observada fue en el sitio ganadero, lo que coincide con lo descrito por Ugalde et al. (2010) y Bojorges (2005) quienes encontraron resultados similares en un bosque templado en el Estado de México y una selva mediana en Veracruz, respectivamente. Sin embargo, muchas de las especies encontradas en el sitio ganadero, como Columbina passerina (Linnaeus, 1758), Columbina tlapacoti, Turdus grayi (Bonaparte, 1838), Volatinia jacarina (Linnaeus, 1766), Cyanocompsa parellina (Linnaeus, 1766) y Passerina cyanea (Linnaeus, 1766) son consideradas como especies generalistas. Esto coincide con lo reportado con Rangel-Salazar et al. (2009), quienes realizaron una comparación entre sitios bajo distintas condiciones de perturbación en un bosque templado en Chiapas y encontraron que el sitio más perturbado presentó mayor riqueza y menor diversidad de aves. Esto también se relaciona con lo descrito por Fahrig (2003, 2017) y De Camargo et al. (2018) quienes argumentan que la riqueza de aves se ve afectada por la pérdida de hábitat y no por la fragmentación. Por otro lado, en la REBICA, que tuvo la mayor superficie de vegetación conservada, menor presión por actividades humanas, y la mayor diversidad estimada, registramos especies como Onychorhynchus coronatus (Muller, 1776) (en peligro de extinción), Formicarius analis (d'Orbigny & Lafresnaye, 1837), Eucometis penicillata (Spix, 1825) (bajo protección especial), Amazilia candida y Chloroceryle aenea, que son especies que han sido reportadas como especialistas y se encuentran enlistadas bajo alguna categoría de protección (SEMARNAT, 2010), que además son consideradas como especies de interior de selva y para las que la perturbación puede ser un factor desfavorable. Hanski (2015) encuentra que sitios con menor perturbación y fragmentación tienden a favorecer la diversidad por lo que, aunque la fragmentación del hábitat puede parecer que incrementa la riqueza, se pierde diversidad y especies especialistas. La ausencia de las especies anteriormente mencionadas en los otros sitios puede estar relacionada con la intensificación de las actividades humanas, situación que puede afectar a cierta clase de aves (Smith et al., 2001). En este sentido, la estructura de la vegetación, como reflejo del uso de suelo, puede determinar la composición de las comunidades de aves (Ugalde et al., 2009).
La riqueza de especies de aves observada en este estudio equivale al 20.3% descrita por González et al. (2016), quienes reportan 403 especies para la REBICA. La diferencia en la magnitud de la diversidad de las comunidades de aves es muy grande entre los sitios, siendo esta magnitud más del doble en los otros sitios en comparación con el sitio ganadero. Los valores de equidad en las comunidades muestran el mismo patrón, siendo el sitio ganadero el que tiene el valor más bajo, situación que resulta de la alta abundancia de pocas especies en este sitio. Tal como encontramos en nuestro estudio, la comunidad de aves en el sitio ganadero tiende a estabilizarse más rápidamente al alcanzar un mismo nivel de cobertura de muestra.
La similitud entre los sitios de la REBICA y los acahuales, coincide con los hallazgos de Smith et al. (2001), quienes mencionan que especies de interior se pueden encontrar en sitios en regeneración; se ha reportado también para bosques de latifoliadas (Vázquez et al., 2017) dónde las áreas en regeneración se recuperan rápidamente y se pueden encontrar en estos sitios especies de aves de selvas maduras o de sitios con mayor estado de conservación. La similitud de las especies entre los acahuales y la REBICA puede significar una transición tendiente a la similitud de las comunidades de aves conforme madura la selva, lo que pudiera influir en la recuperación de las poblaciones de algunas especies. Por el contrario, la mayor diferencia fue entre la REBICA y el sitio ganadero, que actualmente tiene una presión permanente por actividades humanas, lo que explica esta disimilitud en la composición de especies.
Respecto a la abundancia de aves migratorias neotropicales, solamente encontramos diferencias con relación a los meses de muestreo y no en cuanto al uso de suelo. Lo que encontramos no coincide con lo descrito por Robbins et al. (1989) y González et al. (2016), quienes mencionan que la deforestación y el efecto acumulativo del uso de suelo pueden afectar al grupo de aves migratorias. Con base en nuestros resultados, proponemos que las aves migratorias utilizan sitios con distinto uso de suelo y estados de perturbación en los sitios donde pasan el invierno o usan durante la migración más al sur. Se requiere un mayor esfuerzo de muestreo para corroborar esta situación.
Al comparar las abundancias de los gremios encontramos diferencias significativas, con lo que sugerimos que existe una preferencia de uso de los distintos sitios por parte de los gremios, lo cual ha sido descrito en otras investigaciones (Petit & Petit, 2003; Wilson et al., 1996). Pueden existir tendencias en la presencia de gremios en sitios con distinto uso de suelo y cobertura de vegetación, por ejemplo, los semilleros, que en este estudio estuvieron representados por especies generalistas, pueden ser favorecidos en áreas abiertas en las que las presiones antropogénicas son constantes, mientras que otros gremios la respuesta puede ser inversa.
En el sitio ganadero encontramos una mayor abundancia de especies que se registraron en una sola o en dos unidades de muestreo (f1 y f2); sin embargo, la mayoría de estas aves son consideradas generalistas, de espacios abiertos y que se caracterizan por estar presentes en sitios perturbados o bajo presión de actividades humanas. Wall et al. (2012), describieron que los paisajes agroforestales son importantes para este grupo de aves, situación que pudiera estar ocurriendo en este sitio.
De manera general, el uso de suelo determinó la composición de la comunidad de aves, y aunque el CCA explicó solamente el 23% de la variación total, el resultado refuerza lo encontrado con el índice de Jaccard. La distancia entre los acahuales y la presencia e identidad de las especies de aves en ellos muestra una tendencia en la composición de la comunidad. En este sentido, la estructura de la vegetación en los sitios puede determinar la composición de las comunidades de aves (Ugalde et al., 2009), y la variación no explicada por el CCA, puede estar relacionada a condiciones de hábitat y microhábitat, como la estructura, identidad o abundancia de las especies vegetales presentes en cada sitio.
De acuerdo con nuestros resultados en los modelos (GLM) las familias Cardinalidae, Columbidae y Thraupidae son favorecidas por sitios sin vegetación, mientras que la familia Furnariidae, principalmente representada por los trepatroncos, se vio influenciada por los acahuales y la vegetación conservada en la REBICA. Lo anterior remarca la importancia de los acahuales como sitios de transición hacia selvas maduras.
La familia Thraupidae, cuya única especie fue E. penicilata, resultó favorecida por la vegetación conservada y estuvo presente solamente en la REBICA, aunque tuvo una baja abundancia. A nivel de especie, Dendrocincla homochroa (Sclater, 1859), Habia fuscicauda (Cabanis, 1861) y Arremonops rufivirgatus (Lawrence, 1851), pertenecientes a las familias Furniriidae, Cardinalidae y Thraupidae, respectivamente, fueron fuertemente influenciadas por la vegetación, lo que coincide con los resultados encontrados para las familias. Un mayor esfuerzo de muestreo (tiempo y número de redes) podría incrementar la abundancia de algunas especies, lo cual ayudaría a realizar estimaciones más acertadas. Es necesario analizar la influencia de la cobertura de suelo y heterogeneidad de paisaje sobre especies particulares ya que distintas especies pueden responder de diferente manera a los cambios de cobertura, fragmentación y cantidad de hábitat disponible (De Camargo et al., 2018). En este sentido podemos pensar que no solamente las condiciones de uso de suelo favorecen o perjudican la diversidad de aves, sino pueden influenciar la composición de sus comunidades.
Conclusiones
La diversidad de las comunidades de aves fue favorecida por sitios con mayor cobertura de vegetación conservada y menos cobertura de suelo asociada a actividades antropogénicas. La similitud de la composición de aves en sitios con coberturas similares demuestra la importancia e influencia del uso de suelo y vegetación presente en la composición de las comunidades de aves.
El tipo de uso de suelo es un factor importante en la composición de las comunidades de aves y sitios con uso de suelo tendientes a la regeneración o conservación, pueden ser de importancia para la conservación de la diversidad de aves. Creemos que futuros estudios se deben de focalizar en el impacto particular y la influencia del uso de suelo sobre especies que han sido reportadas como amenazadas, tanto residentes como migratorias neotropicales. Ya que los acahuales presentaron una similitud mayor con la REBICA creemos que deben de realizarse esfuerzos para su manejo y conservación que sean tendientes a la transformación gradual en selvas maduras y sitios con mayor cobertura de vegetación conservada ya que esto favorece la diversidad de las comunidades de aves.

