











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
Está ampliamente documentado que México posee una gran diversidad biológica, la cual enfrenta hoy en día graves problemas de conservación (Núñez et al., 2003; Martínez-Meyer et al., 2014). El análisis de la biodiversidad de un área o región contribuye a entender los patrones y procesos, así como el papel que tiene cada especie en los ecosistemas (Thomas, 2013; Vellend, 2017). Su estudio implica registrar, caracterizar, comprender, cuantificar, cualificar y preservar organismos de vida silvestre (Martínez-Meyer et al., 2014). Uno de los grupos más estudiado y numeroso del mundo es el de los insectos. Según McGavin (2002), el 55.3% de los organismos que habitan en el planeta son insectos, con 29 órdenes, 949 familias y más de un millón de especies descritas. Los coleópteros son el grupo más diverso del Orden Insecta (Zhang et al., 2018), cuenta con 392,415 especies descritas, incluyendo 2,928 especies extintas (Zhang, 2013). Los coleópteros han sido sumamente exitosos colonizando diferentes tipos de hábitat, desde áreas terrestres hasta cuerpos de agua dulce (Triplehorn & Johnson, 2005), lo que hace de ellos un grupo de importancia biológica y ecológica (McGavin, 2002), y fundamental para evaluar la calidad de los ecosistemas y su conservación (Ribera & Foster, 1997; Fontúrbel, 2002; Zhang, 2013; Guzmán-Mendoza et al., 2016).
La vegetación y sus características pueden influir en la diversidad de los coleópteros (Escobar, 2000; Sanabria et al., 2008; Jiménez-Sánchez et al., 2013; Sánchez-Reyes et al., 2015; Sánchez-Hernández et al., 2018), por eso la importancia de muestrear varios tipos de vegetación ya que con monitoreos extensos podremos obtener una visión amplia de los organismos que se encuentran en nuestros ecosistemas y así contribuir a su conservación.
Para Chihuahua, el conocimiento de la fauna de coleópteros es escaso. Los primeros registros para el estado fueron realizados por Furth y Jolivet (2009), quienes reportaron 68 especies, 24 géneros y una familia (Chrysomelidae). Dos años después, García et al., (2011) listaron 15 especies, diez géneros y dos familias (Melolonthidae y Scarabaeidae). El Instituto de Biología (IBUNAM) en su Portal de Datos Abiertos UNAM (2014) muestra datos de especímenes recolectados que representan 17 especies, diez géneros y seis familias más (Cantharidae, Geotrupidae, Lamparydae, Lycidae, Meloidae, Telegeusidae). Finalmente, en las bases de datos de la Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO, 2014) se reportan cinco especies más: el pinacate (Eleodes obscurus LeConte, 1851), el botijón (Meloe sp.), la mariquita (Hippodamia convergens Guérin-Méneville, 1842), el escarabajo de cuadros cleritos (Necrobia spp.) y la doradilla (Diabrotica undecimpunctata Mannerheim, 1843). Actualmente, la fauna de coleópteros reportada en el estado de Chihuahua está integrada por 105 especies, pertenecientes a 51 géneros y 14 familias.
Para El Rancho Teseachi ubicado en la parte centro-sur del estado, entre los municipios de Namiquipa, Bachíniva y Guerrero, este trabajo constituye un aporte a la actualización del conocimiento de los Insecta: Coleoptera. Los objetivos de este trabajo fueron: 1) generar un listado de especies de coleópteros de El Rancho Teseachi y 2) comparar la diversidad de coleópteros en dos comunidades vegetales (pastizal de zacate navajita y bosque de encino-pino).
Materiales y Métodos
Área de estudio
El Centro de Investigación y Transferencia de Tecnología Teseachi “CEITT Teseachic” se localiza en el estado de Chihuahua entre los municipios de Namiquipa, Bachíniva y Guerrero (Fig. 1). Teseachi está ubicado geográficamente entre las coordenadas 28° 47’ 42.24’’ N; 107° 26’ 06.79’’ O, con una altitud media de 2,083 m; tiene un clima seco-árido (Bs1kw), con una temperatura media anual de 13°C, la cual puede variar de -5.9°C a 31.6°C; la precipitación pluvial promedio anual es de 580 mm (Trevizo, 2016) y la mayor parte de esta se precipita en los meses de junio-septiembre. Abarca una superficie total de 12,300 ha de las cuales alrededor de 300 ha tienen potencial agrícola y las restantes se dividen en 15 secciones (Alamillo, Área agrícola, Bajío Ancho, Casco, El Estudiante, Fistulados, La Selva, Lajas, Las Coloradas, Palmitas, Potrero Nuevo, Rancho Viejo, Remuda, Savory y Varitas; Cuadro 1), con la finalidad de facilitar el manejo del rancho. La vegetación se caracteriza por la presencia de pastizales de zacate navajita (Bouteloua gracilis (Kunth) Lag. Ex Steud), bosque de encino (Quercus sp.) y bosque de encino-pino (Quercus-Pinus) (UACH, 2011).

Figura 1 Ubicación geográfica del área de estudio en el estado de Chihuahua, México. Los puntos señalan los cinco potreros muestreados.
Cuadro 1 Localización (coordenadas geográficas y altitud) de los sitios muestreados en dos comunidades (pastizal natural y encino-pino) de El Rancho Teseachi, Chihuahua, México.
| LOCALIDADES | LATITUD | LONGITUD | ALTITUD |
|---|---|---|---|
| Bosque de encino-pino | |||
| Lajas | 28°46’24.39’’N | 107°27’33.36’’O | 2,198 m |
| Bajío Ancho | 28°48’26.41’’N | 107°26’37.85’’O | 2,164 m |
| La Selva | 28°47’23.15’’N | 107°26’59.81’’O | 2,209 m |
| El Casco | 28°53’24.48’’N | 107°27’14.98’’O | 1,990 m |
| Alamillo | 28°47’24.42’’N | 107°26’55.29’’O | 2,208 m |
| Pastizal de zacate navajita | |||
| Lajas | 28°46’51.19’’N | 107°27’49.56’’O | 2,164 m |
| Bajío Ancho | 28°48’59’76’’N | 107°26’45.60’’O | 2,080 m |
| La Selva | 28°47’56.30’’N | 107°27’39.62’’O | 2,089 m |
| El Casco | 28°53’00.32’’N | 107°26’56.87’’O | 1,952 m |
| Alamillo | 28°47’14.64’’N | 107°26’24.52’’O | 2,140 m |
Para realizar el muestreo se seleccionaron cinco secciones: Alamillo, Bajío Ancho, El Casco, La Selva y Lajas, con una separación aproximada de 0.5-1.0 km. En cada sección se encontraba un sitio de pastizal de zacate navajita y otro de bosque de encino-pino (Fig.1 ; Cuadro 1), estas secciones en conjunto representan el 59% de la superficie del Rancho Teseachi (CONABIO, 2018). Cabe destacar que debido a que una parte del muestreo se realizó en temporada de lluvias, se coincidió con la época de floración de varias especies de plantas compuestas, especialmente en los pastizales, lo que favoreció la presencia de insectos polinizadores.
Muestreo
La recolección de los individuos se realizó a través de seis muestreos mensuales durante cuatro meses en la temporada de lluvias (junio-septiembre) y dos en la de transición (octubre y noviembre) de 2018. Se emplearon dos métodos de captura de acuerdo con Márquez-Luna (2005): 1) transectos de colecta manual, 2) trampas de caída o “pitfall”. Las trampas de caída consistieron en un recipiente de plástico de un litro de capacidad enterradas a ras del suelo, con 200 ml de alcohol al 70% y estiércol de perro como atrayente (Cárdenas & Hidalgo, 2006). Las trampas se colocaron sobre un transecto lineal de 300 m de longitud distanciadas 50 m una de otra. Las trampas estuvieron activas 24 h y se revisaron dos veces durante ese periodo de tiempo. Se emplearon dos transectos de colecta manual de 50 m de longitud, uno para el pastizal y otro para el bosque de encino-pino dentro de cada sección. En estos transectos se realizó la búsqueda directa de los organismos en las dos comunidades vegetales de cada sección. Una vez localizados los organismos se capturaron con pinzas entomológicas y se depositaron en frascos con alcohol etílico al 70%. Para evitar una subestimación de los resultados, los dos transectos de búsqueda manual establecidos en cada sección estuvieron separados 200 m y se hicieron de manera paralela al sitio establecido para las trampas de caída. A cada transecto se le dedicaron cuatro horas, en un horario establecido entre las 8:00 y 15:00 horas durante el tiempo en que las trampas de caída estuvieron activas.
La identificación taxonómica se realizó bajo un microscopio estereoscópico VE-153G, utilizando las claves de Borror y Delongs (Triplehorn & Johnson, 2005), American Beetles Volumen I y II (Arnett & Thomas, 2000; Arnett, et al., 2002) y con la ayuda de personal del Instituto de Biología, UNAM. Los ejemplares fueron depositados en la Facultad de Zootecnia y Ecología de la UACH en el Laboratorio de Recursos Naturales.
Análisis de datos
La riqueza de especies se definió como el número de especies encontradas en cada comunidad vegetal. La diversidad de especies para cada comunidad vegetal se evaluó a partir del uso del número de especies efectivas propuesto por Jost (2006). Para realizar este análisis se utilizó la diversidad de orden 1( 1 D), que es el exponencial de la entropía del índice de Shannon (diversidad verdadera). El orden ( 1 D) permite incluir todas las especies proporcionalmente a su abundancia en la comunidad (Moreno et al., 2011; Gotelli & Chao, 2013). La abundancia relativa de las especies se estimó como el número de individuos de cada especie presente en cada sitio entre el total de individuos de todas las especies, la cual fue contabilizada por sección y comunidad vegetal. Se usaron curvas de rango-abundancia para comparar la distribución de las abundancias de las especies entre comunidades (Moreno et al., 2011). Para comparar la composición de especies entre comunidades se utilizaron las abundancias relativas y realizó un escalamiento multidimensional no-métrico (NMDS, McCune & Grace, 2002) y un análisis de similitud (ANOSIM). El NMDS mide a través de la función de estrés, escalada entre 0 y 1, qué tan lejanas son las distancias en el espacio multidimensional de las originales en una matriz de asociación (Borcard et al., 2011).
Resultados y Discusión
Riqueza de especies
Se registraron 209 individuos, de los cuales 197 se utilizaron para los análisis, estos están agrupados en 12 familias, 29 géneros y 42 especies y morfoespecies. La familia Scarabaeidae tuvo la mayor riqueza (diez especies) y abundancia (104 individuos), en cambio, Coccinellidae, Curculionidae, Lampyridae, Melyridae y Tenebrionidae fueron las familias con la menor riqueza y abundancia (Cuadro 2). Estos contrastes podrían deberse tanto a las características de los sitios como al tipo de muestreo y los cebos utilizados, ya que para familias como Curculionidae y Lampyridae se necesita establecer trampas o métodos de colecta específicos para lograr su recolección (Sánchez-Soto, 2011; Zaragoza-Caballero, 2015; Pérez-De la Cruz et al., 2016). La familia Erotylidae, los géneros Acmaeodera, Calligrapha, Gibbifer, Harpalus, Odontota, Panagaeus y Pasimachus y las especies Acmaeodera mudgei Westcott, 2002, Calligrapha serpentina Rogers, 1856, Cyclocephala barrerai Martinez, 1969, Gibbifer californicus LaCordaire, 1842, Harpalus rufipes Degeer, 1774, Odontota signaticollis Baly, 1886, Panagaeus sallei Chaudoir, 1862, Pasimachus viridans LeConte, 1858, Chauliognathus misellus Horn, 1885, Chauliognathus profundus LeConte, 1858 y Photinus anisodrilus Zaragoza, 2007 representan nuevos registros para el estado de Chihuahua.
Cuadro 2 Listado de taxa de coleópteros y número de individuos colectados en El Rancho Teseachi, Chihuahua, México, de junio a noviembre de 2018. *Nuevos registros para el estado.
| Especies | Bosque de encino-pino | Pastizal de zacate navajita | Total |
|---|---|---|---|
| Carabidae | |||
| Pasimachus viridans Leconte, 1858* | 2 | 2 | |
| Harpalus rufipes Degeer, 1774* | 2 | 2 | 4 |
| Panagaeus sallei Chaudoir, 1862 * | 2 | 2 | |
| Brachinus sp. | 7 | 7 | |
| Chlaenius sp. | 1 | 1 | |
| Polpochila sp. | 3 | 3 | |
| Platynus sp. | 5 | 5 | |
| Platynus sp. 2 | 2 | 2 | |
| Scarabaeidae | |||
| Cyclocephala barrerai Martinez, 1969* | 1 | 2 | |
| Phanaeus quadridens Say, 1835 | 3 | 3 | |
| Phanaeus sp. | 1 | 1 | |
| Canthon humectus Say, 1832 | 91 | 91 | |
| Xyloryctes sp. | 2 | 2 | |
| Chrysina sp. | 1 | 1 | |
| Onthophagus sp. | 1 | 1 | |
| Euphoria montícola Bates, 1889 | 1 | 1 | |
| Phillophaga sp. | 1 | 1 | |
| Dichotomius sp. | 2 | 2 | |
| Chrysomelidae | |||
| Odontota signaticollis Baly, 1886* | 1 | 1 | |
| Calligrapha serpentina Rogers, 1856* | 1 | 1 | |
| Diabrotica undecimpunctata Mannerheim,1843 | 2 | 2 | |
| Leptinotarsa rubiginosa Rogers, 1856 | 2 | 2 | |
| Leptinotarsa sp.1 | 3 | 1 | 4 |
| Leptinotarsa sp. 2 | 2 | 1 | 3 |
| Lampyridae | |||
| Photinus anisodrilus Zaragoza, 2007* | 2 | 2 | |
| Buprestidae | |||
| Acmaeodera mudgei Westcott, 2002* | 1 | 2 | 3 |
| Acmaeodera sp. 1 | 5 | 5 | |
| Acmaeodera sp. 2 | 2 | 2 | |
| Acmaeodera sp. 3 | 5 | 5 | |
| Acmaeodera sp. 4 | 1 | 1 | |
| Erotylidae* | |||
| Gibbifer californicus LaCordaire, 1842* | 3 | 3 | |
| Cantharidae | |||
| Chauliognathus misellus Horn, 1885* | 1 | 1 | |
| Chauliognathus profundus LeConte, 1858* | 1 | 1 | |
| Chauliognathus sp. | 1 | 9 | 10 |
| Chauliognathus sp. 2 | 5 | 5 | |
| Meloidae | |||
| Nemognatha sp. | 1 | 1 | |
| Nemognatha sp. 2 | 2 | 2 | |
| Lytta sp. | 2 | 2 | |
| Melyridae | |||
| Collops sp. | 3 | 3 | |
| Tenebrionidae | |||
| Eleodes armatus LeConte, 1858 | 3 | 1 | 4 |
| Coccinellidae | |||
| Hippodamia convegens Guérin-Méneville, 1842 | 2 | 1 | 3 |
| Curculionidae | |||
| Sp. 2 | 1 | ||
| TOTAL | 151 | 46 | 197 |
La especie Canthon humectus Say, 1832 fue la más abundante con el 46% del total de los individuos capturados, mientras que Acmaeodera sp. 4, Calligrapha serpentina, Chlaenius sp., Chrysina sp., Euphoria monticola Bates, 1889, O. signaticollis, Onthophagus sp., Phanaeus sp., y Phyllophaga sp., representaron el 0.5% del total de la muestra (Cuadro 2). Halffter et al. (2015) afirman que C. humectus se encuentra en un proceso activo de expansión geográfica ya que es una especie común que se presenta en zonas en las que no se tienen registros previos. Esta alta abundancia de C. humectus coincide con lo encontrado en otros estudios en hábitats similares al presente estudio (Briseño, 2012; Zamora-Vuelvas et al., 2014; Halffter et al., 2015; Carrillo-Ruiz et al., 2017).
Composición de especies
Las comunidades de coleópteros en los dos tipos de vegetación difieren en el número de especies y en la forma en la que se distribuye la abundancia de cada una de ellas (Cuadro 2). Las especies con la mayor abundancia relativa en el bosque de encino-pino fueron: C. humectus (0.46), Brachinus sp. (0.03) y Platynus sp. (0.02), mientras que en el pastizal de zacate navajita, las especies con mayor abundancia relativa fueron: Chauliognathus sp. (0.04), Acmaeodera sp. (0.02) y Acmaeodera sp3 (0.02) (Fig. 2).

Figura 2 Curva de rango-abundancia de dos comunidades de escarabajos muestreados en dos hábitats en El Rancho Teseachi, Chihuahua, México. Cahu= C. humectus, Brsp= Brachinus sp, Plat= Platynus sp, Chau2= Chauliognathus; Chasp= Chauliognathus sp, Acsp= Acmaeodera sp, Acsp3= Acmaeodera sp3, Posp= Polpochila sp.
La mayor riqueza de especies y abundancia (Cuadro 2, 3) se registraron en el bosque de encino-pino (151 individuos y 29 especies), lo cual representa el 77% del total de los ejemplares y el 69% del total de las especies. Este tipo de bosque también contiene el 92% de las familias reportadas en el trabajo. Por otra parte, en el pastizal de zacate navajita hubo menor riqueza (22 especies, 31% del total), menor abundancia (46 individuos, 23% del total) y menor número de familias (n = 9). Sin embargo, la diversidad verdadera encontrada en el bosque de encino-pino fue de 8.5 especies efectivas, mientras que en el pastizal aumentó a 16.6 especies efectivas (Cuadro 3). Es decir, que en el pastizal de zacate navajita se registró casi el doble de la diversidad de coleópteros comparada con el que se registró en el bosque de encino-pino (magnitud de cambio es de 7.6 especies). Cabe resaltar la influencia que tiene la presencia de una especie altamente dominante en el bosque de encino-pino. Si C. humectus fuera eliminado del análisis, la diversidad verdadera del bosque de encino-pino sería de 27.3 especies efectivas (H´= 3.31) y la magnitud de cambio pasaría de 7.6 especies a 11.2 especies, dejando al bosque de encino-pino como la comunidad más diversa.
Cuadro 3 Datos de diversidad de las diferentes comunidades forestales de El Rancho Teseachi, Chihuahua, México.
| Bosque encino-pino | Pastizal de zacate navajita | |
|---|---|---|
| Riqueza de especies | 29 | 22 |
| Abundancia | 151 | 46 |
| Shannon-Wiener (H´) | 2.14 | 2.78 |
| Diversidad verdadera | 8,5 | 16.6 |
| Equidad de Pielou (J´) | 0.60 | 0.90 |
La riqueza de especies y la abundancia de Coleoptera estuvieron repartidas de forma homogénea en los pastizales, lo que se refleja en una mayor equidad (Cuadro 3). Coincidiendo con lo encontrado por Mejía-Narváez (2018), quien concluye que la mayor diversidad de coleópteros se encuentra en sistemas silvopastoriles. Sin embargo, en el bosque de encino-pino se cuenta con gran abundancia, debido a la presencia de la especie dominante C. humectus (Scarabaeidae), que representa el 60% de los individuos capturados y se encuentra asociada exclusivamente a este tipo de vegetación. Su ausencia en el pastizal podría deberse a los componentes del ambiente local (temperatura, precipitación, estacionalidad y características del suelo), así como a la disponibilidad de alimento y zonas óptimas de crecimiento y apareamiento (Trevilla-Rebollar et al., 2010). Es oportuno mencionar que esta especie tiene hábitos coprófagos (Halffter et al., 2015) y puede estar siendo afectada negativamente por la ivermectina (C48H74O14), un desparasitante usado frecuentemente en el ganado vacuno en la región (Thorsten & Moon, 2019). La utilización de este químico tiene efectos negativos en la abundancia y diversidad de múltiples especies; por ejemplo, Iglesias et al. (2005) encontraron reducciones en la abundancia y diversidad de artrópodos coprófagos como resultado del efecto negativo que la ivermectina tuvo sobre la colonización natural de la materia fecal y la duración del proceso de degradación. Así mismo, Baena-Díaz et al. (2018) determinaron que el desparasitante ivermectina afectó a los individuos del escarabajo del estiércol (Euoniticellus intermedius) y su descendencia. Por lo tanto, es posible que también en nuestro sistema la ivermectina esté alterando la abundancia y diversidad de C. humectus y de otras especies.
Por otro lado, existen estudios que muestran que los bosques de encino-pino albergan la mayor riqueza de Scarabaeidae respecto a la selva mediana subperenifolia y el pastizal en el estado de Veracruz (Gómez-Beda, 2013), lo cual coincide con nuestros resultados. Esto se debe probablemente a que las condiciones de temperatura y humedad que caracterizan estos ecosistemas los hacen hábitats deseables para diveros organismos (Arizaga, 2009), ofreciendo así, refugio y alimento a los coleópteros.
El género Acmaeodera (Buprestidae) fue el más abundante en los pastizales. Su alta abundancia en la temporada muestreada puede estar relacionada con la presencia de plantas en floración durante esta época del año. Se sabe que las especies de este taxón se alimentan de polen y de los pétalos de las flores, principalmente (Romero-Nápoles & Westcott, 2017).
Similitud entre las comunidades
Se encontró una baja similitud entre las dos comunidades. La agrupación de los sitios coincidió con el tipo de comunidad presentándose dos grandes grupos (bosque encino-pino y pastizal) (Fig. 3). Los resultados del ANOSIM mostraron diferencias significativas entre las dos comunidades (ANOSIM R= 0.39, p < 0.03). Las diferencias entre las comunidades vegetales parecen explicarse por la presencia de especies exclusivas, por ejemplo, en el bosque de encino-pino: C. humectus, G. californicus, P. viridans y C. misellus; en el pastizal, Acmaeodera sp., Lytta sp., C. barrerai, Phillophaga sp. y C. serpentine. En cambio, siete especies (Acmaeodera mudgei, Chauliognathus sp., Eleodes armatus, Harpalus rufipes, Hippodamia convegens, Leptinotarsa sp.1 y Leptinotarsa sp.2) se compartieron en ambas comunidades (Fig. 3). Según Gómez et al. (2015), los sistemas sin estrato arbóreo (pastizales, entre otros) tienden a mantener una riqueza y abundancia de especies alta, pero la tasa de recambio de especies con otros hábitats tiende a ser baja, es decir poseen gran número de especies exclusivas. Por otro lado, Mejía-Narváez (2018) menciona que la similitud de especies entre bosques y pastizales es muy baja (40% de similitud), estos dos estudios coinciden con lo expuesto en este trabajo, ya que entre las dos comunidades vegetales que se muestrearon se comparten muy pocas especies.
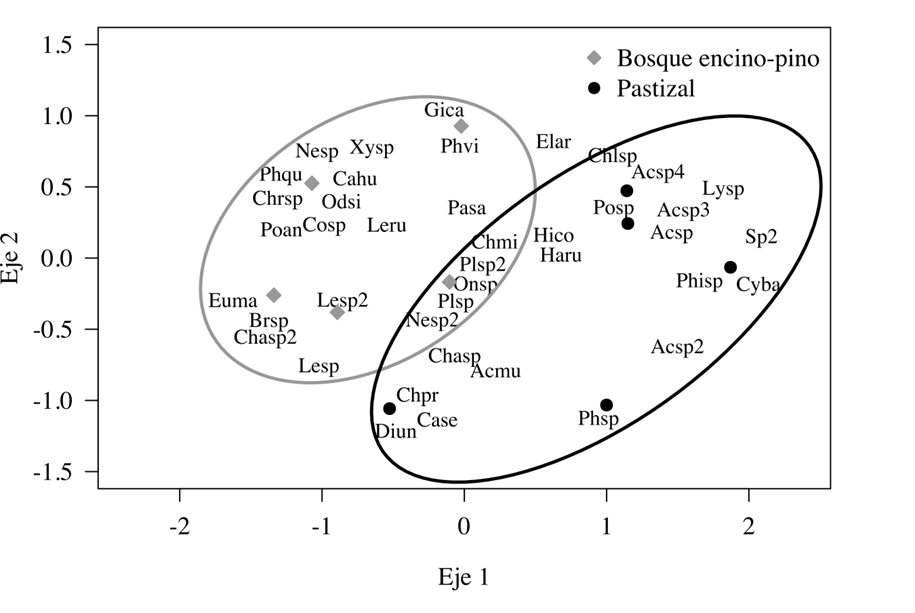
Figura 3 Escalamiento multidimensional no métrico (NMDS) utilizando el Índice de Jaccard. C. humectus= Cahu; Brachinus sp.= Brsp; Platynus sp.= Plsp; Chauliognathus sp2= Chasp2; P. quadridens= Phqu; Leptinotarsa sp.= Lesp; G. californicus= Gica; Collops sp.= Cosp; E. armatus= Elar; P. viridans= Phvi; H. rufipes= Haru; P. sallei= Pasa; Platynus sp2= Plsp2; Xyloryctes sp.= Xysp; L. rubiginosa= Leru; Leptinotarsa sp2= Lesp2; P. anisodrilus= Poan; Nemognatha sp2= Nesp2; H. convegens= Hico; Chrysina sp.= Chrsp; Onthophagus sp.= Onsp; E. montícola= Euma; O. signaticollis= Odsi; A. mudgei= Acmu; C. misellus= Chmi; Chauliognathus sp.= Chasp; Nemognatha sp.= Nesp; Acmaeodera sp.= Acsp; Acmaeodera sp3= Acsp3; Polpochila sp.= Posp; D. undecimpunctata= Diun; Acmaeodera sp2= Acsp2; Lytta sp.= Lysp; Chlaenius sp.= Chlsp; C. barrerai= Cyba; Phanaeus sp.= Phsp; Phillophaga sp.= Phisp; C. serpentina= Case; Acmaeodera sp4= Acsp4; C. profundus= Chpr; Sp2= Sp2.
Las familias presentes en ambas comunidades fueron: Carabidae, Chrysomelidae, Cantharidae y Tenebrionidae, en cambio, Lampyridae, Erotylidae y Tenebrionidae solo se encontraron asociadas al bosque de encino-pino y Curculionidae al pastizal.
Conclusiones
Se reportan diez especies, siete géneros y una familia como nuevos registros para el estado de Chihuahua, incrementando el estado del conocimiento de los coleópteros a 115 especies, 58 géneros y 15 familias. La mayor diversidad de coleópteros se encontró en los pastizales de zacate navajita.
La familia Scarabaeidae tuvo la mayor riqueza de especies y estuvo presente en las dos comunidades forestales. Canthon hummectus fue la especie más común en el bosque de encino-pino, por lo que puede ser una especie que indique la calidad y salud del ecosistema.
El bosque de encino-pino presentó el mayor número de especies exclusivas (20) en comparación con el pastizal (15). Siete especies se compartieron en ambas comunidades.
Se recomienda realizar análisis en los cuales se evalúe el porcentaje de ivermectina en el cuerpo de los escarabajos y los efectos sobre los individuos y su abundancia.

