











 nova página do texto(beta)
nova página do texto(beta) Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroduction
Spiders are animals with a wide array of adaptations that allowed them to colonize most of the terrestrial habitats and becoming a megadiverse order (Foelix, 2011). They rank in 7th place among the richest terrestrial animal groups (Coddington & Levi, 1991), with more than 48,000 species (World Spider Catalog, 2019) and an estimated diversity of about 122,000 species (Agnarsson et al., 2013). Spiders have been evidenced as indicators of different types of environmental characteristics or alterations to the habitat that could be helpful to conservation purposes (Pozzi et al., 1998; Doran et al., 1999; Horváth et al., 2009; Buchholz, 2010; Cristofoli et al., 2010; Magura et al., 2010; Gerlach et al., 2013). However, the spider fauna is still poorly known for several regions of the world (Miller et al., 2014), and this is due in part to the reduced number of taxonomists, collections and to the relative difficulty to explore certain habitats (Coddington & Levi, 1991; Jiménez, 1991). Indeed, the studies of spider fauna are very uneven among different habitats, as an example, the spider faunas of the tropical mountain cloud forests (TMCFs) had been studied only in a few countries, Peru (Silva, 1992), Indonesia (Russell-Smith & Stork, 1994), Tanzania (Sorensen et al., 2002; Sorensen, 2003, 2004), Costa Rica (Yanoviak et al., 2003) and Mexico (Ibarra-Núñez et al., 2011; Maya-Morales et al., 2012; Méndez-Castro & Rao, 2014; Campuzano et al., 2016).
In addition to the canopy, understory and soil levels of many forests, the TMCFs have a great abundance and diversity of epiphytes, in a high humidity environment, due to persistent presence of clouds, mist and high precipitation rates. TMCFs are known by harboring high levels of biodiversity in reduced areas (with high levels of endemism) and by the ecological services they offer to surrounding human populations. At the same time, TMCFs are among the most endangered habitats by direct human influence or by climatic change (Bubb et al., 2004, Sanchez-Ramos & Dirzo, 2014). To preserve the diversity and ecological services of TMCFs, their species composition and their ecological dynamics are two basic subjects to study in view to stablish adequate management programs for this kind of habitats (Bubb et al., 2004).
North America is one of the best studied regions for spider diversity in the world (Coddington & Levi, 1991; Ubick et al., 2017), however, knowledge about spiders is biased to the north (Canada, USA and north of Mexico) and to some vegetation types (Jiménez, 1991). The last published data about the number of spider species recorded from Mexico is about 2,158 (Ibarra-Núñez, 2013), a low value for a country known to be among the ten most biodiverse countries (Martínez-Meyer et al., 2014). Spiders from the Mexican TMCF have been barely explored, with the published studies including only a few localities: Tacaná volcano (Ibarra-Núñez et al., 2011; Maya-Morales et al., 2012), Xalapa (Méndez-Castro & Rao, 2014) and El Triunfo (Campuzano et al., 2016). However, most of these publications do not include information about species composition or just give it partially and only one of these provide a complete list of the spiders found for one of the localities (Ibarra-Núñez et al., 2011). That publication showed that Linyphiidae was much more diverse (second in species richness) in the TMCF of Tacaná volcano than in other tropical sites (Silva, 1996; Silva & Coddington, 1996; Hofer & Brescovit, 2001). The Tacaná volcano study also included a high proportion of undescribed species from a variety of spider families (45.0% of a total 151), suggesting that site as a possible biodiversity hotspot. Similar to those results, in the ground and understory of a TMCF in Tanzania, Sorensen et al. (2002) found 170 spp, of which about 80% were undescribed, with Linyphiidae being third in species richness. As both studies were made on TMCFs, those data are indications that other sites with this vegetation type (as those in the Sierra Madre de Chiapas, Mexico) could have spider assemblages with similar patterns of composition, richness and high proportion of undescribed species.
This study aims to determine the spider fauna from the ground and understory (herbs, shrubs and small trees) of the TMCF in El Triunfo Biosphere Reserve (REBITRI for its Spanish acronym), Chiapas, Mexico, and to analyze if their spider species composition (i.e., species identities), richness and proportion of undescribed species are similar to those found in other TMCFs, including particularly a nearby site in the same mountain range, the Tacaná volcano (Ibarra-Núñez et al., 2011; Maya-Morales et al., 2012). In addition, new species records found at El Triunfo are included with their corresponding images. This study contributes to the knowledge of the biodiversity of a poorly studied and severely threatened habitat. The results found from these spiders’ communities also could be useful in designing conservation strategies for the TMCFs of Mexico and Central America.
Materials and methods
Study area. The study site was located in El Triunfo Biosphere Reserve, Chiapas, Mexico (15º 39’ 48.4’’ to 15º 38’ 54.3 N, 92º 48’ 16.9’’ to 92º 48’ 38.5’’ W, 1,998-2,115 m elevation, Fig. 1); this area comprises the largest and continuous cloud forest in Mexico (Pérez et al., 2010; Gual-Díaz & Rendón-Correa, 2014). Climate in this region is temperate humid with a mean annual precipitation of 3,305 mm and a mean annual temperature of 14.7ºC (CONAGUA, 2014). Dominant tree genera include species of Quercus, Matudea, Dendropanax, Ocotea, Conostegia, Amphitecna, Symplococarpon and Hedyosmum (Long & Heath, 1991; Williams-Linera, 1991).

Field work and taxonomic determinations. Six sampling events were carried out (one per month) in 2014 over the two main seasons in the region, dry (from February to April) and rainy (from June to August). Ground and understory spiders were caught at each sampling event, in ten plots (each one 40 x 16 m) previously established around El Triunfo Camp which were separated by at least 100 m each. Difference in elevation between the highest and lowest plots was less than 100 m. To access most of the spider fauna, we employed four collecting methods (modified from Coddington et al., 1991, Scharff et al., 2003 and Cardoso, 2009). For ground spiders, we used two sampling techniques per plot (Fig. 2): (1) eight pitfall-traps were distributed on both sides of the plot and recovered after 72 hours (each trap filled with 200 ml of propylene glycol at 50% as preservative), and (2) four litter samples (1 m2 each one) were collected from each plot, to be examined (a few hours later) for the spiders. Understory spiders were sampled using two techniques in four circular subplots (5 m diameter, Fig. 2): (1) direct collect in two of the circular subplots, by hand or using an aspirator, investing in each subplot 15 min for low vegetation (from ground surface to knee level) and 15 min for high vegetation (from knee level to arm maximum reach), and (2) sweeping/beating vegetation for the other two subplots, using a sweep net for low vegetation (30 passes) and a stick with a 1 m2 collecting tray for high vegetation (30 strokes). Direct search and sweeping/beating were alternated each month to use both techniques in each subplot. Samplings were performed by four collectors at each sampling event.

Figure 2 Sampling design implemented to collect ground and understory spiders associated to the tropical mountain cloud forest in El Triunfo Biosphere Reserve, Mexico. Ground spiders: pitfall-traps (gray circular dots) and litter ground samples (X marks). Understory spiders: direct search (2 circular subplots in one side of the plot) and sweeping/beating (2 circular subplots in the other side of the plot). See text for details.
Collected spiders were preserved in 96% ethanol and later deposited at the Colección de Arácnidos del Sureste de México (ECOTAAR) in El Colegio de la Frontera Sur, Tapachula, Chiapas, where they were identified to family, genus or species (or morphospecies) making use of taxonomic literature. Some too young immature specimens could not be identified below family rank, as their morphological characteristics did not allow us to do a good species assignment, these specimens were excluded from the species list, but not from the abundance results at family level. Other older immatures have enough characters to be determined to genus, and their association with species was made by comparing their somatic characters with those of adults, but when in doubt they also were excluded from the species list. All identified species and morphospecies were compared with the species recorded by Ibarra-Núñez et al. (2011), which are also deposited in ECOTAAR. Taxonomic classification follows the World Spider Catalog (2019) current arrangement, which also was the information source to detect the species not recorded previously in Mexico, these new records are supported in results with photographic images.
Data Analyses. Data analyses were carried out in R (R Development Core Team, 2018). Differences in species composition between strata (ground vs. understory) and localities (El Triunfo vs. Tacaná volcano) were evaluated using the Colwell and Coddington (1994) complementarity measure and Jaccard similarity index (Magurran, 2004). Colwell and Coddington measure (CC) varies from 0 (for an identical species composition) to 1 (for a completely different species composition), while the Jaccard index (SJ) varies inversely (0 for a completely different species composition and 1 for an identical species composition). In addition, Chao´s estimated proportional similarity (CES, with 95% confidence intervals) was calculated following Miller et al. (2014), who replace values in the Jaccard index with estimated values of Chao 1 shared species and ACE estimator values. SpadeR package was implemented to obtain these estimations (Chao et al., 2016).
Differences in species richness between strata and localities were tested by sample-size-based (i.e., standardized at the same number of individuals) and coverage-based (i.e., standardized at the same coverage) rarefaction/extrapolation curves (Chao et al., 2014). Sampling performance was evaluated by the coverage values obtained from such estimations. In addition, percentage of singletons, sampling intensity, completeness (i.e., observed species number as a percentage of the estimated species number based on the Chao 1 estimator; Magurran, 2004), percentage of undescribed species and rank of most species diverse and most abundant families were used to compare results from this study with results of Tacaná volcano (Chamé, 2011; Ibarra et al., 2011; Maya et al., 2012) and two other studies of spiders from cloud forests (Sorensen et al., 2002; Malumbres-Olarte et al., 2017).
Results
El Triunfo overall results and new records. A total of 7,432 spiders were collected, 1,994 (26.8%) and 5,438 (73.2%) individuals from ground and understory, respectively. From that total 172 too young juveniles could not be assigned to a distinct species/morphospecies, but only to a family, so they were excluded from the species count (but not from the abundances count at family level). The collected specimens represent 28 families, 78 genera and 111 species/morphospecies, from which 41 were determined to species level, 12 as morphospecies related to a described species, 49 to genus, three to family and six were placed in a genus with doubt (Table 1). The 12 morphospecies related to a described species plus 47 of the morphospecies determined only to genus and the six species placed in a genus with doubt (58.6% from total species) are considered undescribed species (65 in total).
Table 1 Species list and abundances of spiders collected in the tropical mountain cloud forest from El Triunfo Biosphere Reserve, Mexico. G, ground stratum, U, understory stratum. SSt, sex-stage: ♂ adult male, ♀ adult female, I immature, € sex undescribed, + species shared with Tacaná volcano, * new species record for Mexico. Species numeration is consecutive to that of Ibarra et al. (2011).
| TAXA | G | U | SSt |
|---|---|---|---|
| AGELENIDAE | |||
| Eratigena sp. 1 aff. E. flexuosa | 1 | - | ♂ |
| Eratigena sp. 2 | 1 | - | ♂ |
| Novalena sp. 2 | 89 | 16 | ♂, ♀ |
| ANYPHAENIDAE | |||
| Anyphaena sp. 11 | - | 74 | ♀ |
| Anyphaena sp. 12 | - | 22 | ♂, ♀ |
| Anyphaena sp. 13 | - | 1 | ♀ |
| Anyphaena sp. 14 | - | 16 | ♀ |
| Anyphaena sp. 15 | - | 8 | ♀ |
| Anyphaena sp. 16 | - | 12 | ♂ |
| Anyphaena sp. 17 | - | 1 | ♀ |
| Arachosia? sp. 1 | 3 | 4 | ♀ |
| Wulfila sp. 1+ | - | 2 | ♀ |
| ARANEIDAE | |||
| Araneus baul Levi, 1991 | - | 20 | ♂€, ♀ |
| Araneus sp. 3 | - | 76 | ♂, ♀ |
| Araneus sp. 4 aff. A. guttatus | - | 1 | ♀ |
| Araneus sp. 5 | - | 8 | ♀ |
| Lewisepeira chichinautzin Levi, 1993+ | - | 21 | ♂, ♀ |
| Manogea? sp. 1+ | - | 31 | ♂, ♀ |
| Micrathena lenca Levi, 1985+ | 1 | 68 | ♂€, ♀ |
| Ocrepeira rufa (O. Pickard-Cambridge, 1889)+ | - | 38 | ♀ |
| CLUBIONIDAE | |||
| Elaver sp. 2 | 2 | - | ♂, ♀ |
| Elaver sp. 3 | 2 | - | ♀ |
| CORINNIDAE | |||
| Castianeira rica Reiskind, 1969 | - | 1 | ♂ |
| Megalostrata raptor (L. Koch, 1866)+ | - | 1 | ♀ |
| CTENIZIDAE | |||
| Ummidia sp. 1 aff. U. zebrina | 1 | - | ♂ |
| Ummidia sp. 2 | 1 | - | ♂ |
| DIPLURIDAE | |||
| Euagrus carlos Coyle, 1988 | 2 | - | ♀ |
| HAHNIIDAE | |||
| Amaloxenops? sp. 1 | 2 | - | ♂, ♀ |
| LINYPHIIDAE | |||
| Agyneta sp. 3 | 461 | 5 | ♂, ♀ |
| Agyneta sp. 4 | 12 | - | ♂ |
| Ceratinopsis sp. 2 | 158 | 2 | ♂, ♀ |
| Fissiscapus sp. 2 | 82 | 4 | ♂, ♀ |
| Gonatoraphis? sp. 1 | 6 | - | ♀ |
| Grammonota teresta Chickering, 1970+ | - | 4 | ♂, ♀ |
| Jalapyphantes cuernavaca Gertsch & Davis, 1946+ | 2 | 40 | ♂, ♀ |
| Jalapyphantes puebla Gertsch & Davis, 1946 | - | 29 | ♂€ |
| Linyphia duplicata (F. O. Pickard-Cambridge, 1902)+ | - | 6 | ♀€ |
| Linyphia linguatula (F. O. Pickard-Cambridge, 1902)+ | - | 1 | ♂€ |
| Linyphia sp. 1 + | - | 21 | ♂, ♀ |
| Linyphia trifalcata (F. O. Pickard-Cambridge, 1902)* | 1 | 144 | ♂, ♀€ |
| Lygarina sp. 1 | 35 | - | ♂, ♀ |
| Microctenonyx? sp. 1 | - | 3 | ♂ |
| Paraletes sp. 3 aff. P. pogo | 22 | - | ♂, ♀ |
| Selenyphantes longispinosus (O. Pickard-Cambridge, 1896)+ | - | 17 | ♂, ♀ |
| Sisis sp. 1 | 14 | 3 | ♂, ♀ |
| Toltecaria sp. 1 | 22 | - | ♂, ♀ |
| Walckenaeria sp. 1 aff. W. crocea+ | 521 | 12 | ♂, ♀ |
| LIOCRANIDAE | |||
| Agroeca? sp. 1 | - | 17 | I |
| MIMETIDAE | |||
| Mimetus sp. 1+ | - | 4 | ♂ |
| MYSMENIDAE | |||
| Mysmena incredula (Gertsch & Davis, 1936)* | 7 | 5 | ♂ |
| NESTICIDAE | |||
| Gaucelmus sp. 1 aff. G. strinatii | 1 | - | ♂ |
| OCHYROCERATIDAE | |||
| Ochyrocera machadoi (Gertsch, 1977) | 38 | - | ♂, ♀ |
| Ochyrocera sp. 2 | 6 | - | ♂, ♀ |
| OONOPIDAE | |||
| Costarina plena (O. Pickard-Cambridge, 1894)+ | 60 | - | ♂, ♀ |
| Oonopidae sp. 2 | 2 | - | ♂, ♀ |
| Pescennina ibarrai Platnick & Dupérré, 2011+ | 1 | 1 | ♀ |
| PARATROPIDIDAE | |||
| Paratropis sp. 1 | 4 | - | ♂ |
| PHOLCIDAE | |||
| Ixchela pecki (Gertsch, 1971) | 3 | 2 | ♂, ♀ |
| Modisimus sp. 1 aff. M. propinquus | 10 | 7 | ♂, ♀ |
| PHRUROLITHIDAE | |||
| Phonotimpus sp. 2 | 87 | - | ♂, ♀ |
| Phonotimpus sp. 3 | 33 | - | ♂, ♀ |
| Phonotimpus sp. 4 | 8 | - | ♂ |
| PRODIDOMIDAE | |||
| Tivodrassus sp. 1 | 1 | - | ♂ |
| SALTICIDAE | |||
| Corythalia nigriventer (F. O. Pickard-Cambridge, 1901)* | - | 7 | ♀ |
| Cotinusa distincta (Pecham & Peckham, 1888) | - | 9 | ♂ |
| Lyssomanes mexicanus Longunov, 2014 | - | 8 | ♂, ♀€ |
| Mexigonus sp. 1+ | - | 127 | ♂, ♀ |
| Phanias sp. 1 | - | 12 | ♂, ♀ |
| Salticidae sp. 1 | 1 | - | ♀ |
| Sarinda sp. 1 | - | 2 | ♂ |
| Zygoballus sp. 1 aff. Z. minutus | 1 | 56 | ♂, ♀ |
| TETRAGNATHIDAE | |||
| Azilia guatemalensis O. Pickard-Cambridge, 1889 | - | 5 | I |
| Chrysometa rincon Levi, 1986 | 1 | 393 | ♂€, ♀ |
| Chrysometa sp. 1 aff. C. palenque+ | - | 68 | ♀ |
| Chrysometa sp. 3 | - | 5 | ♀ |
| Cyrtognatha sp. 1+ | - | 23 | ♂, ♀ |
| Glenognatha spherella Chamberlin & Ivie, 1936+ | 3 | 243 | ♂, ♀ |
| Leucauge simplex F. O. Pickard-Cambridge, 1903+ | - | 1767 | ♂, ♀ |
| Leucauge sp. 2 | - | 110 | ♂ |
| Tetragnatha sp. 1 | - | 3 | I |
| THERAPHOSIDAE | |||
| Theraphosidae sp. 1 | 2 | - | ♀ |
| THERIDIIDAE | |||
| Ameridion sp. 2 | - | 24 | ♂, ♀ |
| Ameridion sp. 3 aff. A. cobanum | - | 1 | ♀ |
| Ameridion sp. 4+ | - | 1 | ♀ |
| Chrosiothes sp. 1 aff. C. wagneri | - | 23 | ♂, ♀ |
| Cryptachaea sp. 1 | - | 1 | ♂ |
| Exalbidion dotanum (Banks, 1914)+ | - | 3 | ♀ |
| Exalbidion rufipunctum (Levi, 1959)* | - | 3 | ♂, ♀ |
| Hentziectypus florens (O. Pickard-Cambridge, 1896)+ | - | 1 | ♀ |
| Neopisinus bigibbosus (O. Pickard-Cambridge, 1896)* | - | 10 | ♂, ♀ |
| Phoroncidia triunfo Levi, 1964+ | - | 55 | ♂€, ♀ |
| Phycosoma altum (Keyserling, 1886)+ | - | 15 | ♂, ♀ |
| Phycosoma sp. 2 | - | 5 | ♂, ♀ |
| Rhomphaea projiciens O. Pickard-Cambridge, 1896 | - | 18 | ♂, ♀ |
| Spintharus flavidus Hentz, 1850+ | - | 13 | ♂, ♀ |
| Tekellina sp. 1 | - | 88 | ♂ |
| Theridion evexum Keyserling, 1884+ | - | 356 | ♂, ♀ |
| Theridula sp. 1 | - | 2 | I |
| Thymoites sp. 4 aff. T. marxi | - | 13 | ♂, ♀ |
| Thymoites sp. 5 | - | 61 | ♂, ♀ |
| Wirada mexicana Campuzano & Ibarra-Núñez, 2018+ | - | 1 | ♂ |
| THERIDIOSOMATIDAE | |||
| Epeirotypus sp. 1+ | - | 665 | ♂, ♀ |
| Theridiosoma davisi Archer, 1953+ | 20 | 372 | ♂€, ♀ |
| Wendilgarda mexicana Keyserling, 1886+ | - | 3 | ♀ |
| THOMISIDAE | |||
| Tmarus vitusus Chickering, 1956+ | - | 14 | ♀€ |
| TRACHELIDAE | |||
| Trachelas sp. 1+ | 1 | 7 | ♂, ♀ |
| ULOBORIDAE | |||
| Philoponella tingens (Chamberlin & Ivie, 1936)+ | - | 28 | ♂, ♀ |
| Uloborus segregatus Gertsch, 1936+ | - | 1 | ♂ |
| Uloboridae sp. 1 | - | 2 | I |
| ZODARIIDAE | |||
| Ishania sp. 2 | 135 | - | ♂, ♀ |
| Ishania sp. 3 | 22 | - | ♂, ♀ |
| Specimens not assigned to a morphospecies | 106 | 66 | I |
| TOTAL | 1994 | 5438 | 7432 |
Tetragnathidae was clearly the dominant family (2,666 individuals, 35.2%), followed by Linyphiidae (1,711 individuals, 23%), Theridiosomatidae (1,060 individuals, 14.3%) and Theridiidae (723 individuals, 9.7%) (Table 1, Fig. 3A). Theridiidae was the richest family (with 20 species, 18.2%), followed by Linyphiidae (19 species, 17.3%), Anyphaenidae and Tetragnathidae (nine species each, 8.2%) and Araneidae and Salticidae (eight species each, 7.3%), while the rest of families have no more than three species (Table 1, Fig. 3B).

Figure 3 Abundance (A, n = 7,432) and species richness (B, n = 111) per family and stratum of spiders collected in the tropical mountain cloud forest from El Triunfo Biosphere Reserve, Mexico. Abundance is noted as Ln of the recorded abundance plus 1 to make visible those families with low abundance values.
One genus and five species were found as new records to Mexico. Lygarina Simon, 1894, new genus record, was cited previously only from South America (World Spider Catalog, 2018). The new species records were: Linyphia trifalcata (F. O. Pickard-Cambridge, 1902) (Figs. 4A-F) previously recorded from Guatemala, Mysmena incredula (Gertsch & Davis, 1936) (Figs. 4G-L) from Bahamas, Cuba, Panama and USA, Neopisinus bigibbosus (O. Pickard-Cambridge, 1896) (Figs. 5A-J) and Corythalia nigriventer (F. O. Pickard-Cambridge, 1901) (Figs. 5K-N) both from Panama, and Exalbidion rufipunctum (Levi, 1959) (Figs. 6A-J) from Ecuador and Panama (World Spider Catalog, 2019) (Table 1). In addition, for 11 species originally described from a single sex, we found specimens from the other sex, seven males and four females from seven families: Araneidae, Linyphiidae, Salticidae, Tetragnathidae, Theridiidae, Theridiosomatidae and Thomisidae (Table 1).
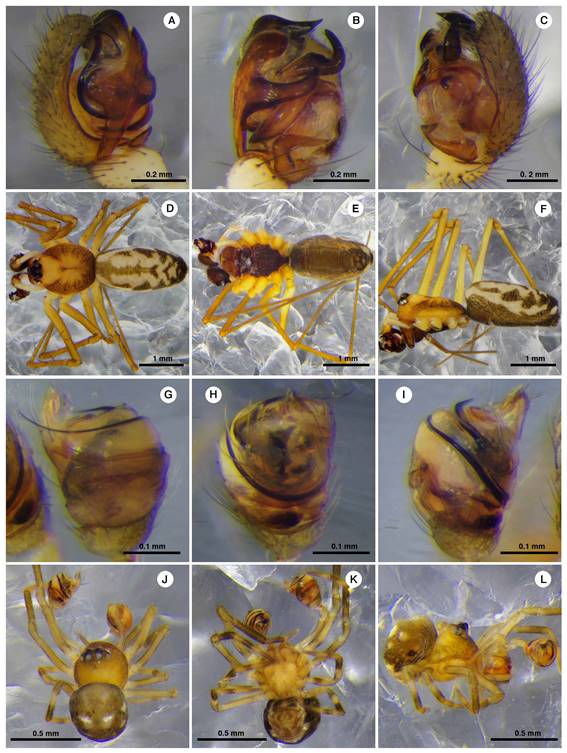
Figure 4 Males of Linyphia trifalcata (Linyphiidae) (A-F) and Mysmena incredula (Mysmenidae) (G-L). Left palpus in prolateral view (A, G), left palpus in ventral view (B, H), left palpus in retrolateral view (C, I), habitus dorsal (D, J), habitus ventral (E, K) and habitus lateral (F, L).
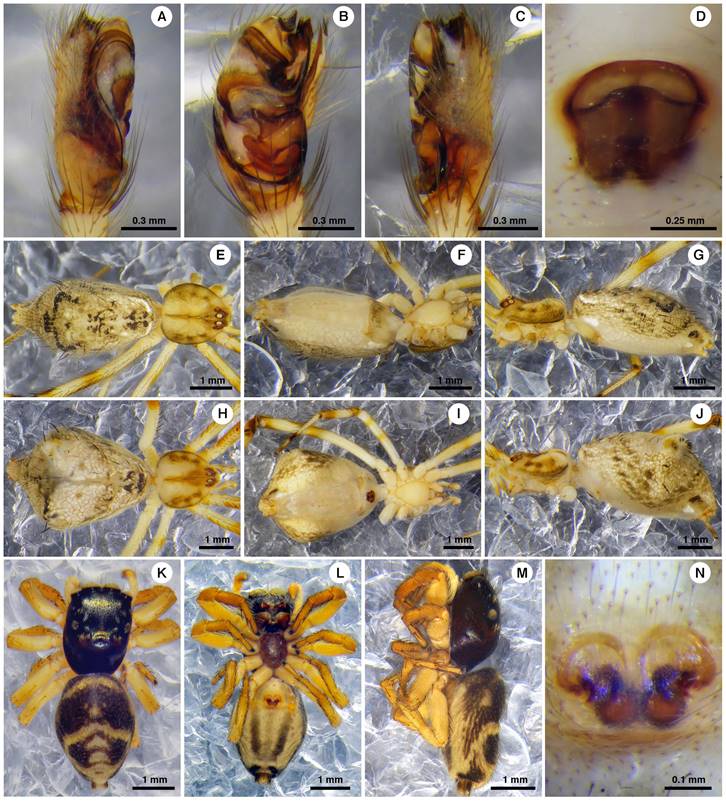
Figure 5 Neopisinus bigibbosus (Theridiidae) (A-J) and Corythalia nigriventer (Salticidae) (K-N). Male (A-C, E-G), female (D, H-N). Left palpus in prolateral view (A), left palpus in ventral view (B), left palpus in retrolateral view (C), epigynum in ventral view (D, N), habitus dorsal (E, H, K), habitus ventral (F, I, L), habitus lateral (G, J, M).

Figure 6 Exalbidion rufipunctum (Theridiidae), male (A-C, E-G), female (D, H-J). Left palpus in prolateral view (A), left palpus in ventral view (B), left palpus in retrolateral view (C), epigynum in ventral view (D), habitus dorsal (E, H), habitus ventral (F, I), habitus lateral (G, J).
Comparisons between ground and understory strata of El Triunfo. In the ground we found 22 families, 35 genera and 46 morphospecies, with Linyphiidae as the dominant family (1,405 individuals, 71.5%), followed by Zodariidae (157 individuals, 7.9%) and Phrurolithidae (128 individuals, 6.4%) (Table 1, Fig. 3A); the richest family was Linyphiidae (12 species, 10.9%) followed by Agelenidae, Oonopidae and Phrurolithidae (each with three species, 6.5%) (Table 1, Fig. 3B). In the understory we found 17 families, 62 genera and 84 morphospecies, with Tetragnathidae as the dominant (2,645 individuals, 48.6%), followed by Theridiosomatidae (1,040 individuals, 19.1%), Theridiidae (710 individuals, 13.1%) and Linyphiidae (306 individuals, 5.6%) (Table 1, Fig. 3A). The richest family was Theridiidae (20 species, 24.1%), followed by Linyphiidae (14 species, 16.9%), Anyphaenidae and Tetragnathidae (nine species each, 10.8%), Araneidae (eight species, 9.6%) and Salticidae (seven species, 8.4%) (Table 1, Fig. 3B). Both strata share 19 species, whereas 27 species were found only on the ground, and 65 species only on the understory (Table 1, Fig. 3B). The complementarity (CC) between these strata was 0.83, while the SJ and CES values were 0.17 and 0.2 (95% C. I. = lower 0.17, upper 0.43) respectively. Sample coverage were estimated at 99.3% and 99.8% for the ground and understory strata, respectively. Understory stratum was significantly more species rich than ground stratum, both up to a standardized sample size (1.45 times richer at 3,776 individuals), as up to a standardized coverage (1.54 times richer at 99.8% of coverage) (Fig. 7A-B).
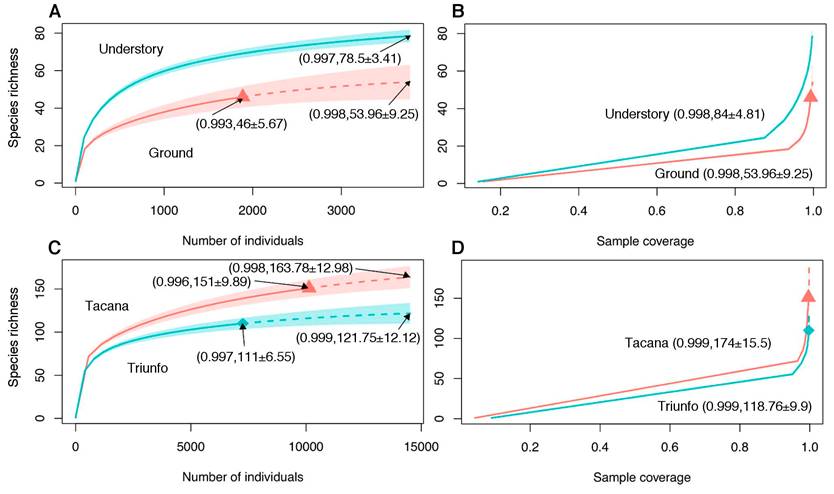
Figure 7 Spider species richness comparison between ground and understory strata of the tropical mountain cloud forest from El Triunfo Biosphere Reserve (A-B), and between El Triunfo and Tacaná volcano localities (C-D). Comparisons based on Chao et al. (2014) methodology using sample-size-based (A, C) and coverage-based (B, D) rarefaction (solid lines) and extrapolation (dashed lines) curves, with 95% confidence intervals (gray-shaded regions). Sample-size curves constructed up to the base sample size (i.e., twice the smaller sample). Coverage-based curves (B, D) calculated over the higher coverage percentage of each comparison. Reference samples (when displayed) are denoted by a different polygon for each curve in each comparison. Numbers in parentheses indicate the sample coverage with their respective richness values and confidence interval at each sample reference and /or comparison level.
Comparisons between El Triunfo and other TMCF sites. The Tacaná volcano study (Chamé, 2011; Ibarra et al., 2011; Maya et al., 2012) and the Sorensen et al. (2002) spiders’ inventory of a Tanzanian TMCF have the most analogous data to be compared with those of this work (Table 2). The Uchumachi (Bolivia) site (cited in Malumbres-Olarte et al., 2017) is partially comparable by having only some of the same data types shown by the other studies. The sampling intensity (for adult specimens) for Tanzania was about 2.6, 3.7 and 14.1 times of those of El Triunfo, Tacaná and Uchumachi, respectively, but the proportion of singletons and the completeness (empiric values) were similar for the Tanzanian and Mexican sites, whereas the Bolivian site had about twice of singletons percentage and the lowest completeness value. Concerning the proportion of undescribed species (empiric values), Tanzania had almost twice the value found in Tacaná and about 1.3 times the value for El Triunfo (Table 2). Theridiidae was the most species rich family in all three sites (there are no data for the Bolivian site), Linyphiidae occupied the second place in the Mexican sites and the third place in Tanzania. The dominant family was different in each site, but the subdominant was Linyphiidae in all three sites (there are no data for the Bolivian site) (Table 2).
Table 2 Data on spiders’ assemblages from studies of different Tropical Mountain Cloud Forest sites with comparable information. (A = Araneidae, C = Cyatholipidae, G = Tetragnathidae, L = Linyphiidae, M = Theridiosomatidae, P = Pholcidae, S = Salticidae, T = Theridiidae, Y = Anyphaenidae, NA = data not available). Source: 1) Malumbres-Olarte et al., 2017. 2) Sorensen et al. 2002. 3) Chamé, 2011; Ibarra-Núñez et al., 2011; Maya-Morales et al., 2012; Ibarra-Núñez data not published. 4) This study.
| Country / Site | Bolivia / Uchumachi | Tanzania / Uzungwa | Mexico / Tacaná | Mexico / Triunfo |
|---|---|---|---|---|
| Abundance: All / Adults | NA / 579 | NA / 9,096 | 9,532 / 2,175 | 7,260 / 2,273 |
| Sobs | 151 | 170 | 151 | 111 |
| Sampling intensity: All / Adults | NA / 3.8 | NA / 53.5 | 63.1 / 14.4 | 66.0 / 20.7 |
| Singletons (%) | 64 (42%) | 32 (18.8%) | 29 (19.2%) | 19 (17.3%) |
| Estimated richness and completeness (Chao1) | 214 (71%) | 197 (86%) | 186 (81.2%) | 125 (88.6%) |
| Undescribed species % | NA | 80% | 41.7% | 58.6% |
| Rank of families for species richness | NA | T > S > L > A (35, 17, 13, 8) | T > L > A = Y (29, 20, 14, 14) | T > L > Y = G (20, 19, 9, 9) |
| Rank of families for abundance | NA | P > L > T > C | T > L > Y > G | G > L > M > T |
| Source | 1 | 2 | 3 | 4 |
For the Mexican sites, our results show that 36 spider species are shared between both sites, with 74 (67.3%) exclusive species for El Triunfo and 115 (76.1%) exclusive species for Tacaná volcano (Table 1), then the complementarity (CC) between both sites was 0.84, while the SJ and CES values were 0.16 and 0.2 (95% C. I. = lower 0.16, upper 0.41), respectively. At higher taxonomic levels, both sites shared 57 genera (113 genera for both sites) and 24 families (38 families for both sites). The families with more shared species between both sites were Theridiidae (8 species), Linyphiidae (7 species), Araneidae and Tetragnathidae (4 species each). From all shared species between both sites, 11 were undescribed species, thus, the number of endemic species for El Triunfo was 54 (65 total undescribed species minus 11 shared undescribed species with Tacaná volcano) corresponding to 48.6%. of endemism. For the Tacaná volcano the number of endemic species is 52 (63 total undescribed species minus 11 shared undescribed species with El Triunfo) corresponding to 34.4% of endemism.
Sample coverages were estimated as 99.7% and 99.6% for El Triunfo and Tacaná volcano, respectively. Concerning the species richness, Tacaná volcano is significantly 1.34 times richer up to a standardized sample size (14,516 individuals) and for any estimated value of a sample size higher than 500 individuals (Fig. 7C). Similarly, Tacaná volcano is significantly 1.46 times richer up to a standardized coverage of 99.9%. Both coverage-based curves show no overlapping for any observed or estimated values (Fig. 7D).
Discussion
In El Triunfo there were clear differences between strata in species richness and composition, as confirmed by the rarefaction/extrapolation analyses (standardized values) and the low similarity values detected. Considering only exclusive species of each stratum (not standardized values), understory (65 species) was almost two times and a half as rich as the ground (27 species). If we consider all species, understory (84 species) was almost twice as rich as the ground (46 species). Differences in number of specimens between strata followed a similar pattern. Nevertheless, there was a slightly higher number of families on the ground, but only about half of genera than the understory. The distribution of the undescribed species between strata also follows this pattern, as 33 undescribed species were collected only on the understory, 22 only on the ground and 10 on both strata.
The higher species richness on the understory could be derived, in part from differences in sampling effort (number of specimens), but also from a higher number of available microhabitats on the understory, due to a more complex habitat (with high diversity of herbs, shrubs and epiphytes) that make possible the cohabitation of a greater number of species (Greenstone, 1984; Halaj et al., 2000). This pattern of higher species number on the understory vs. the ground agrees with other spider surveys in TMCF (Sorensen et al., 2002; Sorensen, 2003; Ibarra-Núñez et al., 2011). Differences in species composition between strata are shown in the high observed and estimated species turnover values. An intermediate complementarity value between ground and understory (CC = 0.58) was previously mentioned for another cloud forest from Tanzania (Sorensen, 2003), but the corresponding values found in our study (0.83) and in another locality in the same mountain range (0.76) by Ibarra-Núñez et al. (2011), suggest greater environmental and/or structural differences between these strata for the Mexican TMCFs, resulting in a noticeable vertical stratification.
Tacaná volcano is located on the Sierra Madre de Chiapas, just 98 km away from El Triunfo (Ibarra-Núñez et al., 2011; Maya-Morales et al., 2012). This relatively short distance generated the expectation that the TMCFs on both sites could share a high proportion of spider species. Nevertheless, the complementarity/similarity values found between both sites shown low similarity at species level, but at genus and family level, the higher proportions of shared taxa (50.4% of total genera and 63.2% of total families) show a probably ancient community shared between these sites. The levels of endemism for both sites are comparable to other groups that inhabit the TMCFs of Mexico, like plants and mammals (Sánchez-Ramos & Dirzo, 2014), but lower to that found for reptiles and amphibians (Gual-Díaz & Mayer-Goyenechea, 2014, Mayer-Goyenechea & Gual-Díaz, 2014) and even for Mexican spiders in another habitat (Rivera-Quiroz et al., 2016). These values could be reduced if in future surveys some of these endemic species are found to be present in other sites. The high species turnover between sites seems due in great part to the observed relatively high proportions of endemics species.
Concerning the observed species richness, in both sites Theridiidae and Linyphiidae were the two most species rich families, followed by Anyphaenidae, Araneidae and Salticidae. The families Araneidae and Theridiidae are the two most diverse in many tropical habitats (Silva, 1996; Silva & Coddington, 1996; Ibarra-Núñez & García-Ballinas, 1998; Höfer & Brescovit, 2001; Ibarra-Núñez, 2013). Our results corroborate Theridiidae as highly diverse in both sites, but Araneidae was only moderately diverse, having the fifth/sixth place (shared with Salticidae). On the contrary, the occurrence of Linyphiidae among the richest families is common for temperate regions (Paquin & Dupérré, 2003; Ubick et al., 2017), but is atypical for tropical habitats, except for some tropical mountain faunas, where this family becomes more diverse (Silva, 1992; Sorensen, 2003; Ibarra-Núñez et al., 2011). It was notorious that Theridiosomatidae was the only family to share all its three species between both sites. Theridiosomatids are known to inhabit mostly in “humid, shaded forest habitats” (Coddington, 1986) as are these cloud forests, highlighting their potential as indicator species of this kind of habitats. Our results suggest that some Linyphiidae species (and perhaps also some Theridiosomatidae species) are taking the place of some Araneidae from tropical lowlands. Similar data were found by Russell and Stork (1994) and Sorensen (2004) in Indonesia and Tanzania, respectively. However, more studies about the distributional patterns for these families are necessary to support this hypothesis. Our results suggest that a high abundance of Theridiosomatidae and Linyphiidae, and a high species richness of Linyphiidae, are indicators of a good conservation level for the Mexican TMCFs. It is also interesting that several genera collected in both sites in Mexico (Fissiscapus, Paraletes, Wirada) or only in one of these sites (Sidymella, Josa, Lygarina) were found previously only in South America (World Spider Catalog, 2019), suggesting some affinities with the fauna from that subcontinent. A similar pattern has been recorded for some tree species in this type of forests (Rzedowski, 1996; Gual-Díaz & Rendón-Correa, 2014).
High proportions of undescribed spider species in tropical habitats have been cited previously (Coddington & Levi, 1991; Sorensen, 2003; Coddington et al., 2009; Miller et al., 2014). Data from different sites, with many undescribed species are not easily comparable in terms of taxonomic composition. Miller et al. (2014) have suggested as a possible solution what they call “Cyberdiversity”, an online community-based approach to compare results (specimens) from independent inventories with incomplete taxonomic knowledge by means of photographic images and/or DNA barcodes. Miller et al. (2014) and Rivera-Quiroz et al. (2016) consider that Cyberdiversity can overcome the taxonomic impediment and accelerate the comparison of specimens at distance, allowing to contrast faunal inventories (even without a name), making possible to know the distribution range of particular species, and enabling the detection of possible new species by taxonomists. We agree with this initiative, as we know it effectively enhance the taxonomic study and comparison of spider faunas. For a new species recently described by Campuzano and Ibarra-Núñez (2018) that was found first in four localities of Chiapas, the access to the images of the Diversity Inventory Pages by Álvarez-Padilla (2017) allowed the authors to know that this species was also found in San Luis Potosí (about 1,000 km away).
The proportions of undescribed species for each site (59% for El Triunfo, 54% for Tacana volcano) show a similar pattern between both sites, with more than half of observed species as new to science. The proportions of potential endemic species (new species for each site minus shared new species) is also high, 48% for El Triunfo, 47% for Tacana volcano. These data and those of Sorensen et al. (2002) showed a high proportion of undescribed species (potential endemic species), give support to the idea that TMCFs are hotspots of diversity for spiders. In spite of the high level in species turnover between the spiders’ assemblages of El Triunfo and Tacana volcano, both have similarities concerning the identity of the families with most species richness, in the families and genera composition, and in the proportions of undescribed species. The latter and the findings of the undescribed sex of several species, highlights the importance to carry out more studies on the spider fauna to better known their true diversity in this kind of habitats.

