











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Las lagartijas son reptiles que se caracterizan por su sensibilidad a los cambios de temperatura y estructura del hábitat, así como por ser relativamente fáciles de observar en ambientes específicos (Smart et al., 2005; Díaz de la Vega-Pérez et al., 2014). La variación en su diversidad es un tema central dentro del estudio de la ecología de comunidades (Cruz-Elizalde et al., 2014), donde la riqueza y composición de especies frecuentemente se asocian con el tipo de vegetación que les provee de microhábitats, sitios de percha y refugio (Huey et al., 1983; Dias & Rocha, 2004; Attum & Eason, 2006). Asimismo, la distancia entre hábitats promueve que su composición y diversidad taxonómica sea distinta, como resultado de la limitada dispersión de estos vertebrados (Cruz-Elizalde & Ramírez-Bautista, 2012; Fernández-López & Lavín-Murcio, 2016; Cruz-Elizalde et al., 2016). Así, algunos trabajos han encontrado que la cercanía entre distintos hábitats favorece una mayor representación de niveles taxonómicos supraespecíficos por el intercambio de especies (Cruz-Elizalde et al., 2014, 2016).
La Sierra El Cuale es un sistema montañoso compartido por los municipios de Puerto Vallarta y Talpa de Allende, Jalisco, en el occidente de México. Se eleva hasta los 2,400 m y presenta nueve tipos de vegetación donde, por su cobertura, sobresalen los bosques tropicales caducifolio y subcaducifolio, pino-encino, encino, pino y mesófilo. Por su riqueza de especies, la Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO) la considera como región prioritaria para la conservación por su potencial aporte a la biodiversidad, mantenimiento de procesos ecológicos, regulación del microclima, regulación de la cantidad y calidad de agua, entre otros (Cruz-Romero et al., 2013).
A pesar de la anterior afirmación, aún se desconocen aspectos sobre la variación de la estructura de los ensamblajes biológicos asociados al ecosistema Sierra El Cuale, en especial sobre el ensamble de lagartijas, cuyo estudio puede aportar elementos para intentar evaluar el impacto de su conservación o los efectos adversos de no hacerlo. Es así que en este trabajo se registran los cambios espaciales y temporales de la diversidad taxonómica de lagartijas en cuatro tipos de vegetación y dos estaciones climáticas (lluvias y seca) de la Sierra El Cuale. Además, se realizó un análisis comparativo de distintividad taxonómica promedio a nivel regional, contra las Reservas de la Biosfera de Chamela-Cuixmala y Sierra de Manantlán.
Materiales y métodos
Los muestreos se llevaron a cabo mensualmente de julio de 2015 a junio de 2016 en cuatro sitios dentro de la Sierra El Cuale, Jalisco, con distinto tipo de vegetación y altitud (expresada en promedio para cada sitio): bosque tropical subcaducifolio (BT), 20°34'14.40'' N-105°11'55.50'' O, altitud 218 m; bosque de encino (BE), 20°32'46.9'' N-105°6'46.61'' O, altitud 959 m; bosque de pino-encino (BPE), 20°30'40.99'' N-105°5'41.74'' O, altitud 1,320 m; y bosque de pino (BP), 20°30'28.35'' N-105°4'56.28'' O, altitud 1,509 m (Fig.1).
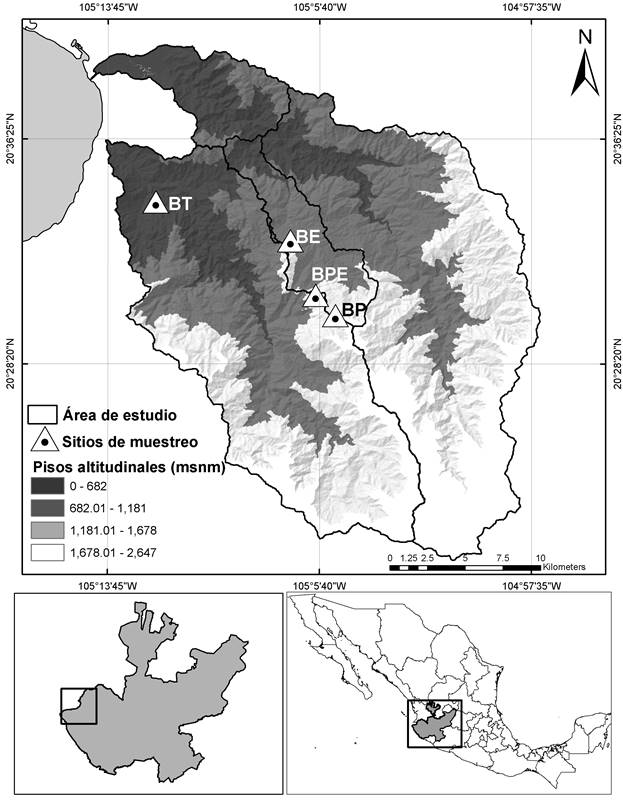
Figura 1 Demarcación de la Sierra El Cuale en el estado de Jalisco, México. Localización de los cuatro sitios de muestreo (triángulos con puntos): 1 = bosque tropical subcaducifolio (BT), 2 = bosque de encino (BE), 3 = bosque de pino-encino (BPE) y 4 = bosque de pino (BP).
En la Sierra El Cuale el clima es cálido subhúmedo (Aw) en los valles y llanuras, semicálido subhúmedo (ACw) en las partes bajas y templado (Cw) en las altas. Se presentan dos estaciones al año: lluvias, de mayo a octubre, y seca, de noviembre a abril (INEGI, 2000).
Para registrar las especies de lagartijas y su abundancia, en cada sitio se realizaron búsquedas visuales en transectos de banda por triplicado de 500 m de largo por 6 m de ancho (Lloyd et al., 1968). Las lagartijas se identificaron a nivel de especie con el apoyo de los trabajos de Ramírez-Bautista (1994), Flores-Villela et al. (1995) y García & Ceballos (1995).
El esfuerzo de muestreo y la riqueza esperada de especies por sitio, se evaluó con rarefacciones basadas en muestras y los estimadores no paramétricos Chao 2, Jackknife 1 y Jackknife 2 (Moreno, 2001). Por su parte, la rareza de especies se determinó con base en las especies únicas y duplicadas de cada sitio de muestreo (Colwell, 2013). Los cálculos se realizaron con el programa EstimateS V 9.1.0® (Colwell, 2013).
Para analizar si la variación del ensamble de lagartijas de la Sierra El Cuale responde al tipo de vegetación y estación del año por sitio, se utilizó un análisis de varianza multivariado basado en permutaciones (PERMANOVA) con dos factores cruzados (estaciones y tipos de vegetación) con efectos fijos (modelo tipo I). Este análisis se realizó con base en una matriz de similitud de Sorensen construida con los datos de presencia y ausencia de lagartijas. La significancia estadística se probó con 10,000 permutaciones y un valor de significancia de p≤0.05 (Anderson, 2005). Por otro lado, se aplicó un análisis de escalamiento multidimensional no métrico (NMDS), a partir de una matriz de similitud de Sorensen, para visualizar las relaciones de similitud de especies entre los tipos de vegetación a través de los periodos climáticos (Clarke & Gorley, 2006).
El grado en que las especies de lagartijas se relacionan taxonómicamente por sitio y estación, se estimó con el índice de distintividad taxonómica promedio (Δ+) (Clarke & Warwick, 1999). Para ello, se construyó una matriz de agregación con seis niveles taxonómicos (phylum, orden, suborden, familia, género y especie) relacionada con los datos de presencia y ausencia de las especies. Los niveles taxonómicos fueron ponderados siguiendo los criterios de Clarke & Gorley (2006), donde las especies dentro del mismo género tienen un peso de w1, especies dentro de la misma familia pero en diferente género tienen un peso de w2, especies dentro del mismo orden pero en diferente familia tienen un peso de w3 y así sucesivamente. Además, con base en 1,000 iteraciones aleatorias sin remplazo, se calcularon valores promedio e intervalos de confianza superior e inferior al 95% de certeza para la Δ+.
Asimismo, se realizó una comparación de la distintividad taxonómica promedio (Δ+) y su variación (Λ+) de las lagartijas a nivel regional entre la Sierra El Cuale con las Reservas de la Biosfera de Chamela-Cuixmala, en la costa sur Jalisco, y Sierra de Manantlán, a 50 km de la costa de Pacífico y localizada en el sureste de Jalisco y noroeste de Colima. Chamela-Cuixmala tiene una amplitud altitudinal entre los 20 a 500 m y vegetación predominante de bosque tropical caducifolio y subcaducifolio (Domínguez-Castellanos et al., 2007); mientras que en Manantlán la altitud varía entre los 500 a 2,400 m, con vegetación de bosque tropical caducifolio, pino, encino y pino-encino (Oliveros-Jiménez, 2016). Para el análisis regional se construyó un modelo de Δ+ con una nueva matriz de agregación taxonómica que consideró a todas las especies de lagartijas presentes en las tres áreas (Clarke & Gorley, 2006). Las listas de especies para las dos Reservas (Cuadro 1) se obtuvieron de los trabajos de Suazo-Ortuño et al. (2008) y Oliveros-Jiménez (2016).
Cuadro 1 Lista de especies de lagartijas, ordenadas alfabéticamente, registradas en este trabajo para la Sierra El Cuale, así como para las Reservas de Chamela-Cuixmala (ChC) (Suazo-Ortuño et al., 2008) y Sierra de Manantlán (Man) (Oliveros-Jiménez, 2016). * = nombre válido para la especie (Meza-Lázaro & Nieto-Montes De Oca, 2015).
| Especies | Cuale | ChC | Man |
|---|---|---|---|
| Anolis nebulosus | x | x | x |
| Aspidoscelis communis | x | x | x |
| Aspidoscelis deppii | x | ||
| Aspidoscelis lineattissimus | x | x | x |
| Aspidoscelis sacki | x | ||
| Barisia imbricata | x | ||
| Coleonyx elegans | x | ||
| Ctenosaura pectinana | x | x | x |
| Elgaria kingii | x | ||
| Gerrhonotus liocephalus | x | x | x |
| Heloderma horridum | x | ||
| Holcosus sinister * | x | x | x |
| Iguana iguana | x | x | x |
| Marisora brachypoda | x | x | |
| Marisora unimarginata | x | ||
| Phrynosoma asio | x | ||
| Phyllodactylus davisi | x | ||
| Phyllodactylus lanei | x | x | |
| Plestiodon brevirostris | x | ||
| Plestiodon colimensis | x | ||
| Plestiodon parvalus | x | x | x |
| Sceloporus bulleri | x | x | |
| Sceloporus grammicus | x | ||
| Sceloporus horridus | x | x | |
| Sceloporus melanorhinus | x | x | x |
| Sceloporus pyrocephalus | x | ||
| Sceloporus scalaris | x | ||
| Sceloporus siniferus | x | ||
| Sceloporus utiformis | x | x | x |
| Scincella assatus | x | x | x |
| Urusaurus bicarinatus | x | x | |
| Número de especies | 12 | 18 | 28 |
| Número de géneros | 9 | 14 | 14 |
| Números de familias | 6 | 9 | 7 |
Los cálculos y análisis de NMDS, PERMANOVA y distintividad taxonómica se realizaron con el paquete PRIMER v6 + PERMANOVA® (Clarke & Gorley, 2006; Anderson et al., 2008).
Resultados
Se obtuvo una riqueza total de 12 especies de lagartijas dentro de siete familias. Siete especies son endémicas del país (Chávez-Avila et al., 2015) y, de acuerdo con la NOM-059-SEMARNAT-2010 (SEMARNAT, 2010), tres se encuentran sujetas a protección especial y una amenazada (Cuadro 2). La riqueza de especies fue mayor en el BE con nueve especies, seguida con ocho en el BT, así como de seis en el BPE y el BP. Por otra parte, todas las especies se registraron en la estación de lluvias y únicamente ocho en la estación seca. Las especies Anolis nebulosus, Sceloporus melanorhinus y S. utiformis se observaron en todos los tipos de vegetación. En cambio, Basiliscus vittatus, Gerrhonotus liocephalus y Scincella assatus se registraron sólo en BT, BPE y BE, respectivamente. De las seis especies restantes, cuatro ocurrieron en dos tipos de vegetación y dos en tres tipos (Cuadro 2). Las curvas de acumulación de especies (rarefacciones basadas en muestras) en cada tipo de vegetación, evidenciaron una representatividad del esfuerzo de muestreo entre el 72% y 91%, donde BP presentó la mayor riqueza esperada y BT la más baja (Fig. 2).
Cuadro 2 Especies de lagartijas registradas en los cuatro tipos de vegetación y por estación del año en la Sierra El Cuale. BT= bosque tropical subcaducifolio, BE= bosque de encino, BPE= bosque de pino-encino, BP= bosque de pino, Ll= lluvias y Se=secas. * = Endémica, + = Sujeta a protección especial y x = Amenazada (SEMARNAT, 2010; Chávez-Avila et al., 2015).
| Familia | Especie | Tipos de vegetación | Estación | ||||
|---|---|---|---|---|---|---|---|
| BT | BE | BPE | BP | Ll | Se | ||
| Anguidae | Gerrhonotus liocephalus + | x | x | ||||
| Corytophanidae | Basiliscus vittatus | x | x | ||||
| Dactyloidae | Anolis nebulosus* | x | x | x | x | x | x |
| Iguanidae | Ctenosaura pectinata*x | x | x | x | |||
| Phrynosomatidae | Sceloporus bulleri* | x | x | x | x | ||
| Sceloporus melanorhinus | x | x | x | x | x | x | |
| Sceloporus utiformis* | x | x | x | x | x | x | |
| Scincidae | Plestiodon parvulus* | x | x | x | x | x | |
| Scincella assatus | x | x | |||||
| Teiidae | Aspidoscelis communis*+ | x | x | x | x | x | |
| Aspidoscelis lineattissimus*+ | x | x | x | x | |||
| Holcosus sinister | x | x | x | x | |||
| Riqueza taxonómica | |||||||
| Número de especies | 8 | 9 | 6 | 6 | 12 | 8 | |
| Número de géneros | 6 | 7 | 4 | 4 | 9 | 5 | |
| Número de familias | 5 | 5 | 4 | 4 | 8 | 4 | |
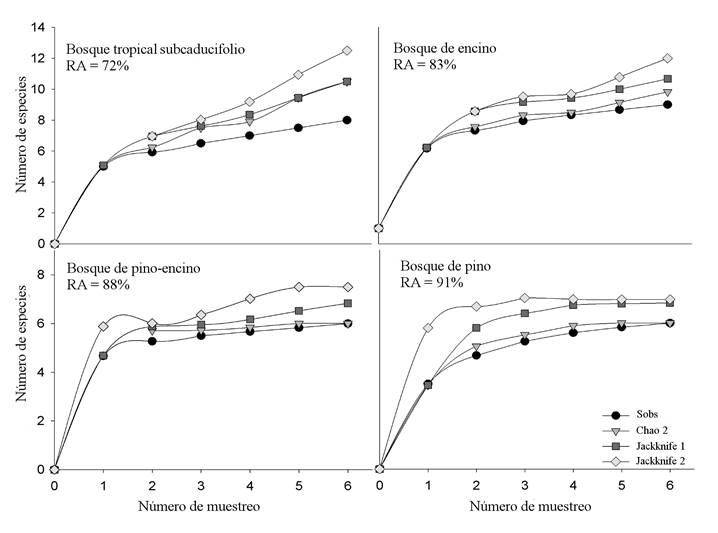
Figura 2 Curvas de acumulación de especies y valores de riqueza acumulada (RA) para los cuatro tipos de vegetación de la Sierra El Cuale, Jalisco. Sobs = riqueza observada.
En los análisis de similitud de especies, los resultados del PERMANOVA mostraron que existen diferencias significativas entre la composición de especies de lagartijas por los tipos de vegetación (p<0.0001), pero no se presentaron diferencias entre estaciones climáticas (p= 0.2636), ni entre la interacción de los tipos de vegetación con las estaciones climáticas (p= 0.255). Las pruebas pareadas no mostraron diferencias significativas en la composición de especies entre BPE y BP (p= 0.406), así como BT y BE (p= 0.052); pero sí entre BT y BPE (p= 0.001), BT y BP (p= 0.002), BE y BPE (p= 0.002) y BE y BP (p= 0.001). Por su parte, el análisis NMDS, con un valor de “stress” de 0.02, permitió identificar dos grupos: uno formado por BPE y BP y, el otro, por BT y BE (Fig. 3).
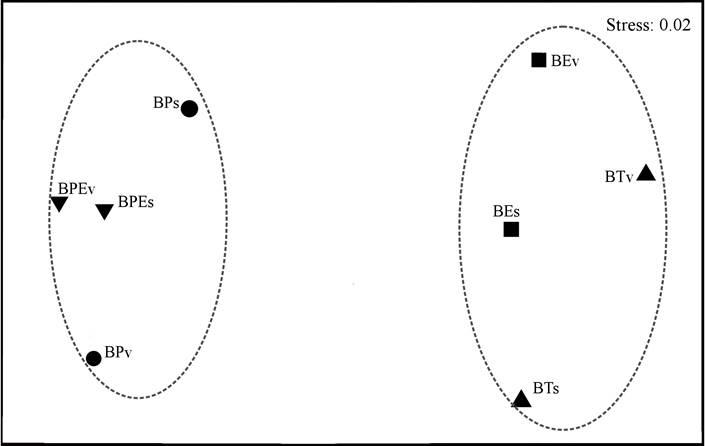
Figura 3 Gráfico del análisis multidimensional no métrico (NMDS) de la composición de especies de lagartijas para los cuatro tipos de vegetación y dos estaciones del año en la Sierra El Cuale, Jalisco. Simbología: Bosque tropical subcaducifolio lluvias (BTv) y seca (BTs), cuadrado = bosque de encino lluvias (BEv) y seca (BEs), triángulo inverso = bosque de pino-encino lluvias (BPEv) y seca (BPEs) y círculo = bosque de pino lluvia (BPv) y seca (BPs).
Por estaciones del año, el valor de distintividad taxonómica promedio fue mayor para lluvias (Δ+ = 26.5), donde se contabilizó la mayor riqueza de especies (12), próximo a la media esperada Δ+ = 26.5. Los valores para lluvias y seca se mantuvieron dentro de los intervalos de confianza (p>0.05) (Fig. 4A). Para los tipos de vegetación, el valor de Δ+ fue de 26.1 para BE y de Δ+ = 25.9 para BT. En ambos tipos de vegetación, además de registrase los mayores valores de Δ+ y próximos a la media esperada, se contabilizó el mayor número de especies, con nueve y ocho, respectivamente. Para BPE y BP, donde se registró la menor riqueza de especies (6), se calcularon valores de Δ+ de 23.8 que estuvieron por debajo de la media esperada. Los valores de Δ+ se observaron dentro de los intervalos de confianza para todos los tipos de vegetación (p>0.05) (Fig. 4B).
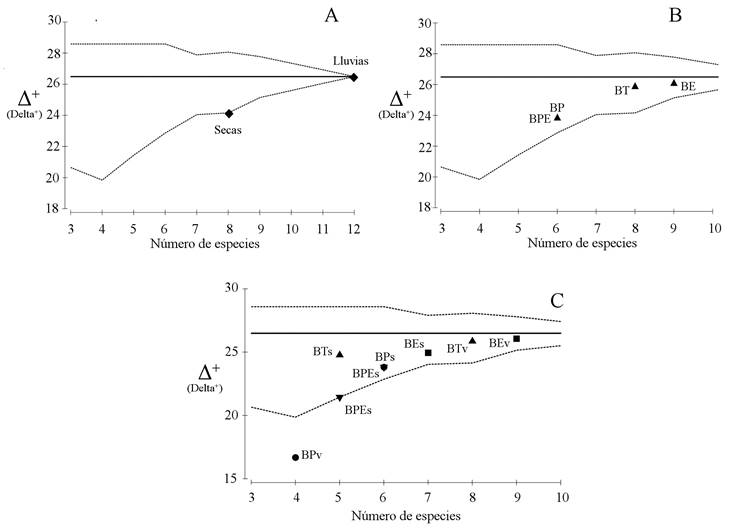
Figura 4 Distribución simulada de la distintividad taxonómica promedio Δ+ (línea discontinua) e intervalos de confianza del 95% (línea continua) para las especies de lagartijas de la Sierra El Cuale. A) Análisis temporal; B) Análisis espacial; C) Análisis espacio-temporal. Simbología Figura 4C: triángulo = Bosque tropical subcaducifolio lluvias (BTv) y seca (BTs), cuadrado = bosque de encino lluvias (BEv) y seca (BEs), triángulo inverso = bosque de pino-encino lluvias (BPEv) y seca (BPEs) y círculo = bosque de pino lluvias (BPv) y seca (BPs).
La distintividad taxonómica promedio entre los tipos de vegetación a través de las estaciones del año fue más alta durante la estación de lluvias en BE (Δ+ = 26.1) y BT (Δ+ = 25.9), mientras que en la estación seca fue más baja con Δ+ = 24.9 y Δ+ = 24.8, respectivamente. Los valores de Δ+, tanto en BE como en BT, se observaron próximos al valor de la media esperada (Δ+ = 26.5) y dentro del canal de probabilidad (p>0.05) (Fig. 4C). El BPE presentó una Δ+ mayor en lluvias (Δ+ = 23.8) y menor en seca (Δ+ = 21.4). Para BP, el valor de Δ+ fue mayor en seca (Δ+ = 23.8) y menor en lluvias (Δ+ = 16.7). El BPE y BP presentaron valores de Δ+ distantes de la media esperada y, salvo para la estación de lluvias en BP donde el valor se ubicó fuera del canal de probabilidad (p≤0.05), el resto se observó dentro (p>0.05) (Fig. 4C).
Los valores de distintividad taxonómica a nivel regional, en orden descendente, entre Chamela-Cuixmala, Sierra del Cuale y Manantlán fueron de Δ+ igual a 26.6, 26.3 y 25.1, respectivamente. El valor de distintividad taxonómica para Manantlán fue el único que se ubicó por debajo de la media regional esperada (Δ+ = 25.1). Los valores para las tres regiones se ubicaron dentro del canal de probabilidad (p>0.05) (Fig. 5A). En cuanto a riqueza de especies de lagartijas, en Manantlán se contabilizaron 28 especies, 18 en Chamela-Cuixmala y 12 en este estudio dentro de la Sierra El Cuale. Por su parte, la mayor variación de la Λ+ en Manantlán (Λ+ = 61.0) indicó poca uniformidad en el arreglo taxonómico, lo que representa una cantidad importante de especies con diferentes categorías de agregación taxonómica. En cambio, se estimó menor variación en la Sierra El Cuale (Λ+ = 41.4) y Chamela-Cuixmala (Λ+ = 35.2), lo que manifiesta un arreglo taxonómico con más uniformidad. El valor de la variación de la distinción taxonómica para Manantlán fue el único por arriba del valor de la variación media-regional esperada (Λ+ = 60.0) (Fig. 5B).
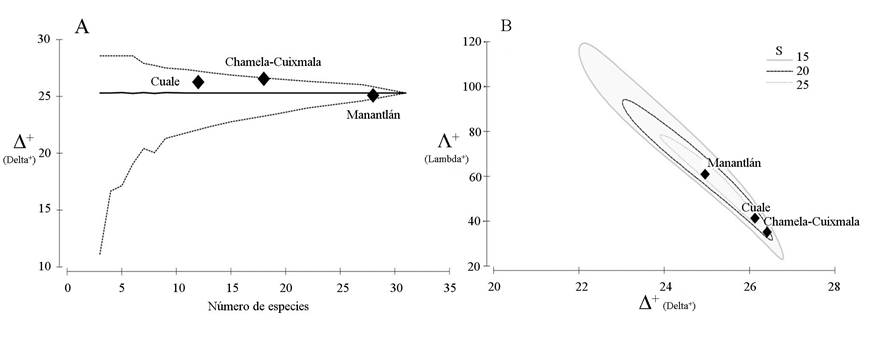
Figura 5 Distintividad taxonómica promedio (Δ+) simulada de la lista de especies de lagartijas (A) y la relación entre la distinción taxonómica promedio (Δ+) y su variación (Λ+) (B), para la Sierra El Cuale y las Reservas de la Biosfera de Chamela-Cuixmala y Sierra de Manantlán. En la figura 5B la S representa las líneas de la riqueza de especies en el modelo.
Discusión
El trabajo registró por primera vez 12 especies de lagartijas para la Sierra El Cuale, dentro de cuatro de los nueve tipos de vegetación presentes en el área. Se observó que la riqueza de especies en su conjunto para la zona (o riqueza regional), fue resultado de la contribución individual de cada tipo de vegetación (o riqueza local). En términos de conservación, la desaparición de alguno o varios tipos de vegetación, como por ejemplo BE y BPE, traería como consecuencia la disminución potencial en cinco especies, una de ellas sujeta a protección especial, de la riqueza regional de la Sierra El Cuale (ver Cuadro 2).
La heterogeneidad de especies de lagartijas por tipo de vegetación, significativamente diferente, puede deberse a la especificidad de uso que brindan sus hábitats disponibles (Pianka 1967; Tews et al., 2004) o a factores ambientales como temperatura o altitud (Lara-Reséndiz et al., 2014). Aunque no disponemos de datos de comportamiento, alimentación o ambientales confiables para establecer las relaciones entre la riqueza de lagartijas y tipo de vegetación, la frecuencia de registro de una especie en uno o más hábitats puede aportar información sobre si sus requerimientos son generalistas o especialistas (Martín-Regalado et al., 2011); así, para la Sierra El Cuale, se podrían considerar a B. vitattus, G. liocephalus y S. assatus como especialistas por registrarse en un tipo de vegetación y a A. nebulosus, S. melanorhinus y S. utiformis como generalistas por su presencia en todos los ambientes.
Aunque en otros estudios de reptiles, la mayor riqueza de especies se registra en la temporada de lluvias, cuando el recurso alimenticio es más abundante que en la estación seca (Garza-Castro et al., 2006; Vite-Silva et al., 2010), en este trabajo no fue posible establecer diferencias significativas entre ambas estaciones. En este caso, ciertas especies de lagartijas adoptan estrategias según la estación, que consisten en el aumento y disminución de las actividades de forrajeo, de comportamiento territorial y de desplazamiento (García et al., 2010).
Por su parte, la riqueza observada de lagartijas por tipo de vegetación, según los estimadores Chao 2, Jackknife 1 y Jackknife 2, está por debajo de la esperada entre una y cuatro especies. El trabajo de Chávez-Avila et al. (2015) proporciona información sobre la posible identidad de las especies de lagartijas restantes, a través de modelos de patrones de abundancia y distribución. De acuerdo con estos autores, las especies restantes que potencialmente se pueden observar en la Sierra El Cuale son: Aspidoscelis deppei, Heloderma horridum y S. heterolepis.
La diferenciación en los tipos de vegetación que se determinó con el análisis PERMANOVA y se observó en el NMDS, tal vez responde a la presencia de especies de lagartijas que hacen uso exclusivo de uno u otro grupo formado. De esta forma, se tienen a C. pectinata, A. communis y A. lineattissimus en el grupo BT y BE, mientras que S. bulleri se presentó exclusivamente en el grupo BPE y BP. Esta distribución diferenciada en las especies, puede estar relacionada con su capacidad de adaptación y dispersión, así como con la disponibilidad de recursos en los distintos tipos de vegetación (Chávez-Avila et al., 2015).
La distintividad taxonómica promedio (Δ+) fue mayor en la estación de lluvias por la presencia de más niveles taxonómicos con respecto a la seca. Para BT, BE y BPE se registraron dos géneros y dos familias más en lluvias que en seca. Solo BP tuvo dos familias y dos géneros más en la estación seca en relación con la de lluvias. Por tipo de vegetación, los mayores valores se observaron en BT y BE, ya que las especies correspondieron a diferentes géneros y familias; en cambio, BPE y BP, donde la Δ+ fue menor, la variedad de grupos taxonómicos fue pobre. Es decir, la riqueza taxonómica de la Sierra El Cuale (riqueza regional) depende del aporte individual de cada tipo de vegetación en las distintas estaciones del año (riqueza local).
En el estudio comparativo exploratorio entre la Sierra El Cuale y las Reservas de la Biosfera de Chamela-Cuixmala y Manantlán, las dos primeras presentaron un arreglo taxonómico más uniforme que la tercera. Es posible que la mayor o menor uniformidad registrada para los tres sitios, esté relacionada con la presencia de ambientes tropicales en El Cuale y Chamela Cuixamala, así como de bosques templados en Manantlán. Los ambientes tropicales han tenido más tiempo para diversificarse a diferencia de los templados, donde el tiempo de diversificación de las especies ha sido menor (Wiens & Donoghue, 2004). Asimismo, este resultado reconoce la importancia regional de la Sierra El Cuale en la diversidad de lagartijas de la región centro occidente de la costa del Pacífico.

