











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El aguacate es un frutal perenne originario de regiones tropicales y subtropicales de Centroamérica, perteneciente a la familia Lauraceae (Ish-Am et al., 1999; Castañeda-Vildózola et al., 1999; Peña, 2003; Alcaraz & Hormaza, 2009; Pérez-Balam et al., 2012). Existen tres razas: la antillana (P. americana var. americana Mill.), la guatemalteca (P. americana var. guatemalensis L. Wms.) y la mexicana [P. americana var. drymifolia (Schlecht. y Cham.) Blake], las cuales presentan diferencias en la forma, color, tamaño, textura de los frutos y tolerancia al frío y la humedad (Bernal & Díaz, 2008; Alcaraz & Hormaza, 2009). Han dado origen a cultivares con diferentes restricciones en la fecundación; algunas son autoestériles y otras autofértiles (Wysoki et al., 2002; Bernal & Díaz, 2008; Alcaraz & Hormaza, 2009).
En el aguacate para evitar la autopolinización se presentan grupos complementarios de acuerdo con el patrón de apertura floral, denominados A y B. Este mecanismo se denomina protoginia dicogamia sincronizada. Las flores tipo A en la primera apertura están en fase femenina, tienen el estigma receptivo y las anteras dehiscentes, cierran al medio día y abren al día siguiente en la tarde en fase masculina. Las flores tipo B abren en fase femenina en la tarde, luego cierran y abren nuevamente a la mañana siguiente en fase masculina (Davenport, 1986; Ish-Am, 2004; Bernal & Díaz, 2008; Alcaraz & Hormaza, 2009).
En algunos cultivares se presenta asincronía en la apertura floral, lo cual causa traslape entre las fases florales, así, no se requiere sembrar variedades de apertura floral complementaria en los huertos de aguacate (Wysoki et al., 2002; Alcaraz & Hormaza, 2009). El cultivar Hass es un híbrido resultante del cruce de las variedades guatemalteca y mexicana. Este cultivar presenta receptividad estigmática en la segunda apertura floral, por lo cual puede autopolinizarse (Alcaraz & Hormaza, 2009).
En el cultivo de aguacate, se presentan tres mecanismos de polinización de acuerdo con la procedencia del polen que ingrese a la flor, denominados: autogamia, geitonogamia y polinización cruzada. Siendo esta última la de mayor aporte para la producción de frutos (Davenport, 1986; Wysoki et al., 2002). Las flores atraen grupos específicos de polinizadores, de acuerdo con la morfología floral y los recursos nutricionales que ofrecen, estas interacciones se han catalogado como síndromes de polinización (Rosas-Guerrero et al., 2014). Las flores de P. americana no presentan interacciones especializadas; la disposición de las estructuras secretoras de néctar y reproductivas facilita el acceso a insectos de distintos órdenes (Ish-Am et al., 1999).
La importancia de los insectos en la polinización del cultivo de aguacate ha sido discutida (Wysoki et al., 2002; Cabezas & Cuevas, 2007; Ish-Am & Lavah, 2011) y se estima que en algunas regiones la visita de los insectos no es indispensable para la polinización de este frutal (Davenport et al., 1994; Davenport, 2003). En cultivos del sur de Florida (EEUU) la deposición de polen en la segunda apertura floral es llevada a cabo por el viento (Davenport et al., 1994). En cambio, en Israel y el sureste de España es necesario el transporte de polen por insectos, es decir la polinización cruzada prevalece sobre la autopolinización (Ish-Am & Eisikowitch, 1998; Cabezas & Cuevas, 2007). La especie presenta polinización anemófila, pero predomina una entomófila generalista, debido a la morfología y el comportamiento de apertura floral (Wysoki et al., 2002; Ish-Am & Lavah, 2011; Carabali et al., 2017).
En Colombia se reconoce la importancia de los insectos como moscas, avispas, hormigas, abejas Trigona y especies introducidas como A. mellifera para la polinización del cultivo de aguacate (Vásquez et al., 2011). Incluso, que el rendimiento se incrementa entre un 21-96% con la actividad de las abejas (Vásquez et al., 2011). Sin embargo, se requiere profundizar en el aporte de las especies en la producción y los efectos que podría tener la ausencia de polinizadores en los huertos. El objetivo de esta investigación fue determinar el efecto de la exclusión de insectos asociados a las inflorescencias de Persea americana cv. Hass y correlacionar su presencia/ausencia con la fructificación de las inflorescencias.
Materiales y métodos
Área de estudio. El estudio se realizó en dos huertos en monocultivo de aguacate Hass, de cinco años, con producción anual estable, ubicados en Popayán, Cauca, Colombia. Finca “Jireh”, N02° 27’ 21,9” O76° 34’ 09,9”; 1886 metros sobre el nivel del mar (msnm) y finca “Dios es Amor” N02° 27’ 47,2” O76° 34’ 05,0”; 1873 msnm, la temperatura anual promedio es de 19°C y precipitación de 1941 mm/año. La zona es de reconocida presencia de visitantes florales y polinizadores (Carabalí et al., 2017). Al inicio del experimento las fincas estaban en el periodo de floración principal, con inflorescencias en el estado, que se caracteriza porque las flores están completamente diferenciadas pero cerradas, de acuerdo con la escala fenológica propuesta por Salazar-García et al. (1998). El área circundante a los huertos incluye varias hectáreas de bosque de eucalipto, cultivos de café y potreros. Durante la floración no se aplicó insecticidas en las fincas para evitar afectación de los polinizadores.
Diseño experimental. Se utilizó un diseño completamente al azar, con tres tratamientos y 10 repeticiones por tratamiento. En cada huerto se seleccionaron aleatoriamente 30 árboles para la comparación de tres tratamientos de polinización, se asignaron 10 árboles al azar a cada uno los siguientes tratamientos:
T1- Tratamiento con inflorescencias descubiertas: Marcaje de 20 inflorescencias por árbol, las inflorescencias no se cubrieron para permitir así la visita de insectos a las flores (control).
T2- Tratamiento con inflorescencias cubiertas: Marcaje de 20 inflorescencias por árbol y cubrimiento de las inflorescencias con bolsas de tela tipo tul (30x30 cm, con una apertura menor a 0,1 mm) con el fin de evitar el acceso de los insectos a las flores.
T3- Tratamiento con inflorescencias cubiertas + insecticida: Marcaje de 20 inflorescencias por árbol, cubrimiento de las inflorescencias con bolsas de tela tipo tul (30x30 cm, con una apertura menor a 0,1 mm) y aplicación de insecticida de contacto e ingestión (Spinetoram®; dosis 0,5 cc/L) no tóxico para abejas (una vez seco el producto en la lámina foliar) una sola vez, con el fin de eliminar microinsectos presentes en las flores.
En la aleatorización se excluyeron los árboles ubicados en el borde del cultivo para minimizar este efecto en las variables a evaluar. Se seleccionaron las inflorescencias de la zona baja y media del árbol. Las inflorescencias se embolsaron en días diferentes entre las 7:00h hasta las 18:00 h.
Se registraron las siguientes variables: número inicial de flores, el número de frutos cuajados (fructificación), la abscisión de frutos por inflorescencia. El conteo de frutos cuajados se llevó a cabo a los 20 días de realizar la implementación de los tratamientos hasta finalizada la floración.
El porcentaje de frutos formados se calculó de la siguiente manera:
Porcentaje (%) de frutos cuajados= (N° de frutos cuajados/ N° de flores inicial) x100.
Una vez que se obtuvo el número total de frutos en cada inflorescencia, las bolsas fueron removidas para permitir el desarrollo de los frutos.
Análisis de datos. El número de frutos producidos fue comparado mediante un análisis de varianza (ANOVA). Se realizó una prueba de comparación de medias (Tukey, P ≤ 0,05) para detectar diferencias entre los tratamientos. Se evaluó si se presentaban diferencias estadísticamente significativas entre las fincas mediante una prueba T-Student con un nivel de significancia del 0,05. Los análisis se realizaron en el programa MINITAB versión 16 (Minitab, State College, PA, EEUU).
Resultados
En la finca “Jireh” el número de frutos cuajados difirió significativamente entre tratamientos (F=230,27; g.l.= 2; P=0,000). Una mayor formación de frutos (cuajado) se presentó en el tratamiento donde los insectos no fueron excluidos, comparados con aquellos, donde hubo una exclusión física y química de los mismos. En el tratamiento con inflorescencias descubiertas se obtuvo un cuajado de fruto siete veces mayor al obtenido en los tratamientos de inflorescencias cubiertas y cubiertas + insecticida. El número de frutos cuajados en las inflorescencias en que se excluyeron los insectos (i.e. tratamientos de inflorescencias cubiertas y cubiertas + insecticida) no difirió significativamente entre ellos.
En la finca “Jireh”, cuando las flores fueron visitadas por insectos, en la mayoría de las inflorescencias se produjo entre 10-30 frutos, con valores atípicos entre 58-77 frutos. En contraste, cuando se elimina la participación de los insectos, se obtiene de uno a tres frutos por inflorescencia, con un máximo de 22 frutos en el tratamiento con inflorescencias cubiertas o de 15 frutos con las inflorescencias cubiertas + insecticida. En los todos los tratamientos se presentaron inflorescencias en las que hubo una abscisión total de las flores (Fig. 1a).
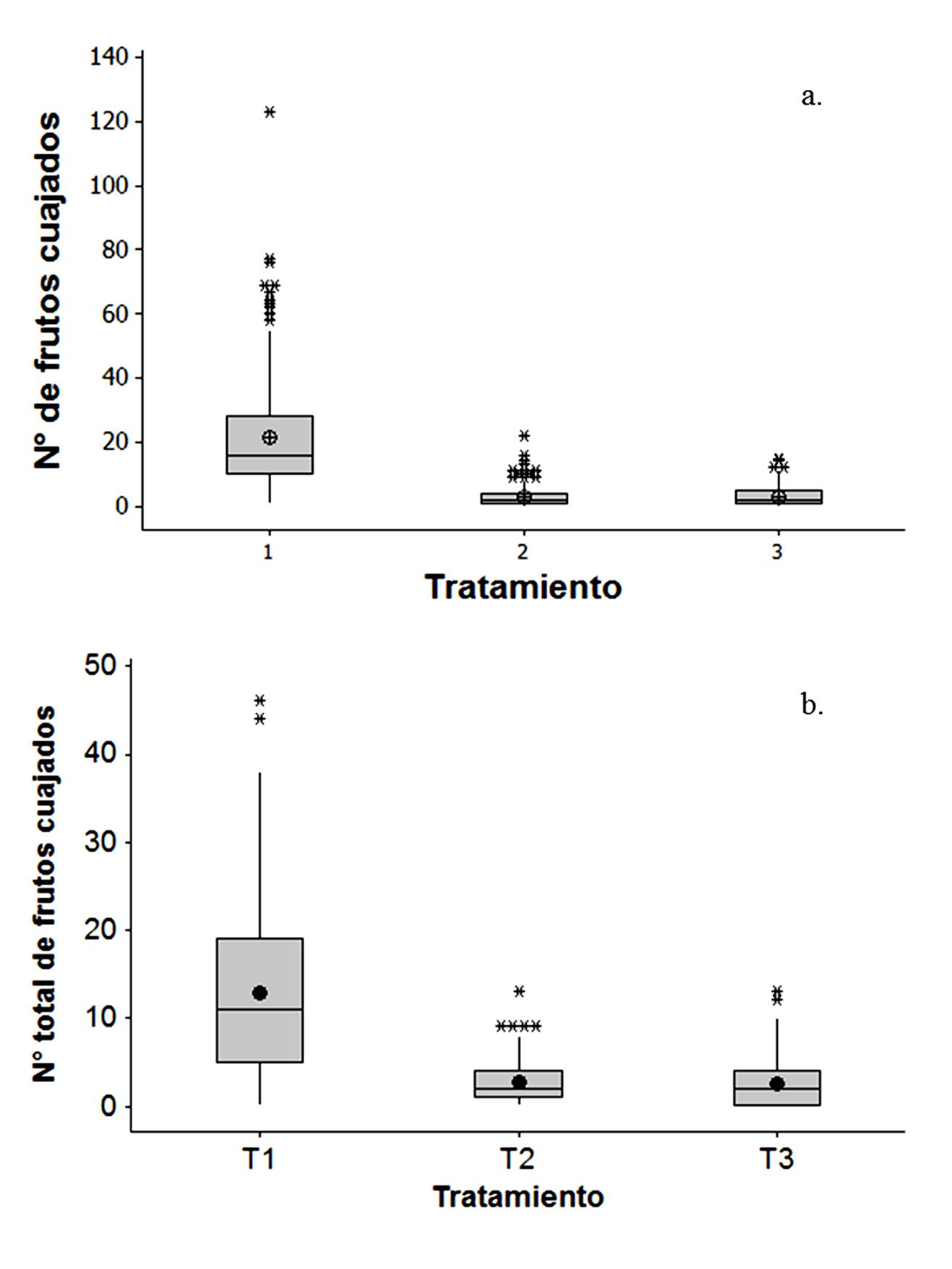
Figura 1 Variabilidad en el número de frutos cuajados de aguacate cv. Hass obtenido en: a) la finca “Jireh” y b) la finca “Dios es amor” usando tres tratamientos que influencian la polinización. (T1) tratamiento con inflorescencias descubiertas, (T2) tratamiento con inflorescencias cubiertas, (T3) tratamiento con inflorescencias cubiertas + insecticida. Se muestra un diagrama de caja. Los límites de la caja corresponden al rango intercuartil, la línea horizontal indica la mediana, el punto oscuro indica la media, los bigotes se extienden a los datos extremos y los asteriscos indican los valores atípicos.
En la finca “Dios es Amor”, el número de frutos cuajados difirió significativamente entre tratamientos (F=194,55; g.l.= 2; P=0,000). Se encontraron significativas diferencias en el tratamiento con inflorescencias descubiertas con respecto a la exclusión física (tratamiento con inflorescencias cubiertas) y la exclusión física y química (tratamiento con inflorescencias cubiertas + insecticida). En estos últimos, con el cubrimiento de las flores y la aplicación de insecticida se produjo un cuajado de frutos entre 4-7 veces menor, siendo, significativamente similares entre ellos. En el tratamiento con inflorescencias descubiertas, la mayor parte de las inflorescencias produjo entre 5 y 19 frutos, con valores atípicos entre 44-46 frutos. En el tratamiento con inflorescencias cubiertas e inflorescencias cubiertas + insecticida se obtuvieron entre 1-4 frutos, con valores atípicos de 9-13 frutos (Fig. 1b).
Diferencias significativas se presentaron en los análisis del número de frutos cuajados, obtenidos en los tratamientos de polinización entre fincas (F= 375,69; g.l.= 1; P=0,000). Los tratamientos con inflorescencias cubiertas e inflorescencias cubiertas + insecticida tuvieron un porcentaje de cuajado de fruto similar entre fincas, en comparación a la alta variabilidad obtenida en el tratamiento con inflorescencias descubiertas. En la finca “Jireh” se obtuvo un promedio de 21 frutos cuajados/inflorescencia cuando los insectos tenían acceso a las flores, mientras que en la finca “Dios es Amor” se tuvieron 12 frutos/inflorescencia. La exclusión de los insectos mediante bolsas de tela generó un promedio de 2-3 frutos por inflorescencia (Cuadro 1). En los tratamientos con inflorescencias cubiertas e inflorescencias cubiertas + insecticida se presentaron frutos sanos y algunos con malformaciones.
Cuadro 1 Cuantificación del cuajado de fruto de aguacate cv. Hass obtenido en tres tratamientos que influencian la polinización. (T1) tratamiento con inflorescencias descubiertas, (T2) tratamiento con inflorescencias cubiertas, (T3) tratamiento con inflorescencias cubiertas + insecticida.
| Parcela | Trat. | Inflorescencias | Promedio de flores |
Total de frutos cuajados |
Promedio de Frutos |
|---|---|---|---|---|---|
| Finca “Jireh” | 1 | 200 | 93.13±2.44 | 4266 | 21.33 ± 1.24 a* |
| 2 | 200 | 109.43±2.78 | 596 | 2.98 3.64 | |
| 3 | 200 | 106.16± 2.79 | 619 | 3.095 ± 0.20 b | |
| Finca “Dios es Amor” | 1 | 200 | 153.72 ± 3.88 | 2546 | 12.73 ± 0.69 c* |
| 2 | 200 | 145.53 ± 3.20 | 525 | 2.625 ± 0.18 b | |
| 3 | 200 | 142.47 ± 3.91 | 483 | 2.415 ± 0.19 b |
Con la actividad de los insectos en las flores se produjo un mayor porcentaje de frutos cuajados en ambas parcelas, comparado con los tratamientos donde los insectos son excluidos por barreras físicas y físico-químicas. La exclusión de la visita de los insectos generó que se obtuviera un cuajado de fruto entre 4-9 veces menor que cuando los insectos tenían libre actividad en las flores (Cuadro 2). El porcentaje de cuajado post-antesis (i.e. inicial) en la Finca “Jireh” fue tres veces mayor al de la finca “Dios es amor”. La proporción de flores que se convierten en fruto y continúan su desarrollo a cosecha fue menor al 1%, de acuerdo con el porcentaje de cuajado final (Cuadro 2, Fig. 2) (se estimó con el número de frutos que continúan su desarrollo a los 120 días de la asignación de los tratamientos a las unidades experimentales). Estos resultados concuerdan con otras investigaciones sobre la alta abscisión natural de flores y frutos (Cuadro 3).
Cuadro 2 Porcentaje de fruto cuajado inicial y final de aguacate cv. Hass obtenido en tres tratamientos que influencian la polinización. (T1) tratamiento con inflorescencias descubiertas, (T2) tratamiento con inflorescencias cubiertas, (T3) tratamiento con inflorescencias cubiertas + insecticida.
| Parcela | Tratamiento | Total de flores | Cuajado inicial de frutos (%) | Cuajado final de frutos (%) |
|---|---|---|---|---|
| Finca “Jireh” | 1 | 18625 | 24.2 | 0.349 |
| 2 | 21886 | 2.8 | 0.078 | |
| 3 | 20032 | 3.2 | 0.065 | |
| Finca “Dios es Amor” | 1 | 30744 | 8.2 | 0.088 |
| 2 | 29105 | 1.9 | 0 | |
| 3 | 28495 | 1.9 | 0.032 |
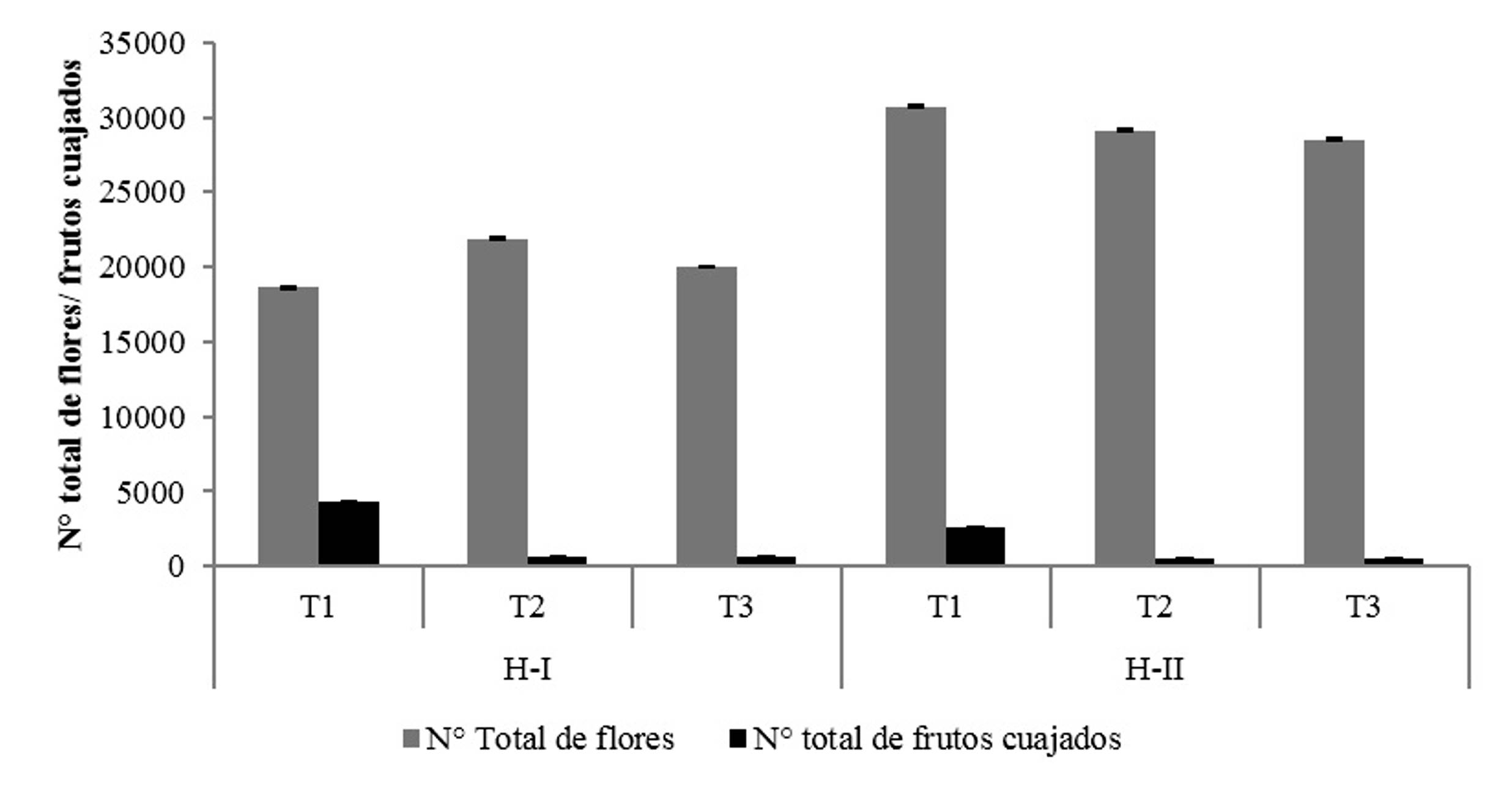
Figura 2 Número total de frutos cuajados en inflorescencias seleccionadas de tres tratamientos que influencian en la polinización de las fincas “Jireh” y “Dios es Amor” respecto al número de flores.
Cuadro 3 Número total de frutos cuajados en inflorescencias marcadas en el tiempo en aguacate cv. Hass usando tres tratamientos que influencian la polinización. (T1) tratamiento con inflorescencias descubiertas, (T2) tratamiento con inflorescencias cubiertas, (T3) tratamiento con inflorescencias cubiertas + insecticida.
| Parcela | Tratamiento | Frutos cuajados | |||||
|---|---|---|---|---|---|---|---|
| 20 días | 40 días | 60 días | 80 días | 100 días | 120 días | ||
| Finca “Jireh” | 1 | 3202 | 2205 | 353 | 184 | 65 | 65 |
| 2 | 508 | 151 | 34 | 29 | 17 | 17 | |
| 3 | 534 | 167 | 28 | 20 | 13 | 13 | |
| Finca “Dios es Amor” | 1 | 117 | 2236 | 831 | 215 | 215 | 27 |
| 2 | 490 | 48 | 9 | 7 | 7 | 0 | |
| 3 | 154 | 362 | 22 | 15 | 15 | 9 | |
Los análisis del seguimiento de frutos cuajados que continúan su formación hasta madurez fisiológica revelaron diferencias entre tratamientos. En la finca “Jireh”, luego de 120 días del marcaje o exclusión de la visita de insectos, de acuerdo con el tratamiento se obtuvo: en el tratamiento con inflorescencias descubiertas solo se conservaba el 1,52 % de los frutos, en las inflorescencias que se limitó la visita de insectos se conservaba entre el 2,85% (Tratamiento con inflorescencias cubiertas) - 2,1% (Tratamiento con inflorescencias cubiertas + insecticida) de los frutos. En la finca “Dios es amor”, Se presentó mayor variabilidad en la proporción de frutos cuajados que continuaron su desarrollo; en el tratamiento 1 se conservaban el 1,06% de los frutos, en el tratamiento 2 hubo abscisión de todos los frutos y en el tratamiento 3 el 1,86% de los frutos continuaron su desarrollo.
Discusión
El efecto benéfico de los insectos como visitantes florales en la polinización del cultivo de P. americana ha sido ampliamente documentado (Wysoki et al., 2002). La morfología floral de esta especie presenta adaptación a la polinización por insectos debido a su tamaño, color y la disposición de los verticilos (Faegri & van der Pijl, 1979; Ish-Am et al., 1999; Peña, 2003). Los resultados del presente estudio corroboran la importancia de la visita de los insectos a las flores de P. americana cv. Hass en el incremento del cuajado de fruto. Por otra parte, se excluye la participación de insectos con tallas menores a 3 mm ya que se presentan valores similares al comparar el cuajado de fruto en inflorescencias cubiertas con o sin aplicación de insecticidas. En cultivos de aguacate Hass ubicados en Morales (Cauca), localidad cercana a la zona de estudio se encontró una alta diversidad de visitantes florales, 35 familias de insectos frecuentan las flores de este cultivo; principalmente dípteros de las familias Syrphidae, Tachinidae, Calliphoridae, Sarcophagidae e himenópteros de las familias Vespidae y Apidae (Carabalí et al., 2017). La abeja A. mellifera es el visitante más frecuente, pero también frecuentan el cultivo abejas silvestres como Tetragonisca angustula, Scaptotrigona barrocoloradensis, Partamona cf. aequatoriana, Trigona sp. (Carabalí et al., 2017). En México, la amplia entomofauna que visita las flores contribuye diferencialmente a la polinización debido a una alta variabilidad en la carga polínica, atribuida al tamaño corporal y movilidad en las flores (Ish-Am et al., 1999; Castañeda-Vildózola et al., 1999).
La polinización entomófila es esencial para la producción de aguacate en Israel, Sudáfrica, California y Chile, debido a que la autopolinización y la polinización por el viento son ineficientes (Peterson, 1955; Ish-Am & Eisikowitch, 1998; Ish-Am & Lavah, 2011). En el tratamiento donde se permitió la libre actividad de los visitantes florales se presentó un cuajado de fruto significativamente mayor, respecto a lo encontrado en las inflorescencias cubiertas, estos resultados concuerdan con investigaciones previas para el cultivar Hass. En el Sureste de España, la exclusión de los insectos con bolsas microperforadas limitó el desarrollo de los frutos, incluso se desarrollaron frutos partenocárpicos o deformes (Cabezas & Cuevas, 2007), como lo observado en los tratamientos con exclusión físico-química. En California, las variedades Regina, Hass, Clifton, Bacon y Ryan no presentan cuajado de fruto cuando se cubren las inflorescencias para excluir a la abeja Apis mellifera (Peterson, 1955).
El porcentaje de cuajado de fruto encontrado en el tratamiento que no se excluyeron los insectos es menor al obtenido en huertos de México. En aguacates criollos, La actividad de los insectos genera un cuajado de fruto superior al 60% respecto a las inflorescencias en que son excluidos (Can-Alonzo et al., 2005). Valores similares de porcentaje de cuajado fueron encontrados por Pérez-Balam y colegas (2012), cuando los insectos son excluidos ocurre un cuajado de fruto del 8,3%, este es inferior al obtenido por polinización cerrada y cruzada entre 55,3-58,9%. Sin embargo, estos estudios coinciden en que la actividad de los insectos incrementa el cuajado de fruto.
En las inflorescencias cubiertas, la agitación de las ramas por el viento podría facilitar la dispersión del polen. Los estudios sobre el aporte del viento en la polinización del cultivo de aguacate muestran resultados contrastantes, algunos indican que es un vector ineficiente (Peterson, 1955; Ish-Am & Eisikowitch, 1998; Ish-Am & Lavah, 2011) mientras que en otros muestran que hay una contribución a la producción de frutos (Davenport et al., 1994; Davenport, 2003; Can-Alonzo et al., 2005). Entre el 1-3% de la polinización de un cultivo es atribuible al viento en algunas regiones del mundo (Ish-Am & Lahav, 2011). Sin embargo, en el sur de California, árboles del cultivar Hass son polinizados durante la primera apertura floral (fase femenina) por la acción del viento, siendo este el principal vector de polen, a pesar de la presencia de A. mellifera (Davenport, 2003). Variedades como Simmonds y Hardee en la Florida se autopolinizan durante la segunda apertura floral; la fructificación puede ocurrir incluso en inflorescencias cubiertas, así que la presencia de polinizadores no es requerida (Davenport et al., 1994). En este estudio se encontró que inflorescencias cubiertas se autopolinizan, no obstante, el número de frutos cuajados es significativamente menor, por lo tanto, se requiere la actividad de los insectos para obtener cuajado de fruto.
Los resultados del presente estudio muestran que se puede obtener un mayor cuajado de fruto (siete veces superior) en inflorescencias de aguacate descubiertas, donde los visitantes florales tuvieron una mayor actividad, respecto a las inflorescencias cubiertas. Dado que estudios previos indican que el aguacate depende de la polinización entomófila para obtener rendimientos adecuados, los resultados encontrados sugieren que la actividad de polinizadores, visitantes florales y ocasionales es responsable del cuajado de fruto en las inflorescencias descubiertas. Se considera que el viento podría tener una menor contribución en la fructificación.

