











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
La fauna edáfica integra complejas redes tróficas contribuyendo con un amplio rango de servicios esenciales para el funcionamiento sustentable de los ecosistemas. Su composición y diversidad se relaciona con el tipo, edad, diversidad, estructura y manejo de los cultivos (Flores et al., 2008). Debido a su rápida respuesta a los cambios en el hábitat, a sus ciclos de vida relativamente cortos, a su facilidad de muestreo y a su alta diversidad en sistemas agrícolas, los organismos edáficos han cobrado importancia como herramienta para evaluar el funcionamiento y sustentabilidad del suelo (Kremen et al., 1993; Gibbs & Stanton, 2001).
Entre la mesofauna, los ácaros oribátidos y los colémbolos son los artrópodos más abundantes y de mayor diversidad específica en el suelo. A través de su actividad fragmentan y transforman los restos orgánicos en residuos disponibles para los mineralizadores, dispersan bacterias y hongos, y afectan el crecimiento y la composición específica de estos últimos por consumo selectivo (Coleman et al., 2004). Distintos autores han señalado su sensibilidad a las prácticas agrícolas, que se manifiesta en la reducción de su abundancia, de su diversidad y en la simplificación de su estructura comunitaria (Bedano et al., 2006, 2011; Socarrás, 2013).
Entre la macrofauna, las arañas son depredadoras generalistas importantes por su abundancia, biomasa y riqueza de especies (Wise, 2006). Ocupan distintos microhábitats, consumen diferentes presas, tanto de la meso como de la macrofauna, al mismo tiempo, presentan estrategias variadas de caza, resisten la falta de alimento y la desecación; reducen y mantienen a las poblaciones de insectos que consumen los cultivos por debajo de los umbrales de daño económico (Pedigo, 2001; Lawrence & Wise, 2000). Por su parte, los insectos coleópteros presentan una elevada riqueza de especies y abundancia, muchos participan, directa o indirectamente, en el control de especies fitófagas, en la descomposición, en la regulación del ciclo de nutrientes y en la polinización (Symondson et al., 2002; Van Driesche et al., 2008). La macrofauna conecta las redes hipogeas y epigeas por su amplio nicho espacial, las arañas por su alta capacidad de dispersión y amplitud trófica; y los coleópteros debido a los cambios de hábitat asociados a sus distintas etapas de desarrollo. Ambos grupos, son sensibles a las variaciones en la cobertura y estructura vegetal, así como a cambios de la cantidad de materia orgánica, agregación y otras propiedades físicas y químicas del suelo (Cole et al., 2005).
En el Cinturón Hortícola Platense (CHP), el tomate (Lycopersicum esculentum Mill.) se encuentra entre las hortalizas más cultivadas. Convencionalmente, su producción está asociada a la práctica de laboreos intensivos y al manejo sanitario por medio de la aplicación de distintas sustancias químicas de síntesis, como fertilizantes, herbicidas, fungicidas y plaguicidas (Hole et al., 2005). El objetivo general de este trabajo fue estudiar a la comunidad edáfica, epigea e hipogea de un cultivo convencional de tomate, durante un ciclo agrícola, para establecer un modelo estructural y funcional, que sirva como punto de comparación para evaluar los posibles beneficios de la aplicación de prácticas agroecológicas. Los objetivos específicos fueron: detectar a los grupos de especies más representativos, a nivel taxonómico y funcional de oribátidos, colémbolos, arañas y coleópteros; y proponer un modelo que integre a las taxocenosis de oribátidos, colémbolos, arañas y coleópteros, a nivel estructural y funcional, durante el ciclo del cultivo del tomate.
Materiales y métodos
Área de estudio. El estudio se llevó a cabo en una parcela de 150 m2, a cielo abierto, sembrada con tomate variedad platense, en un predio perteneciente al CHP. A nivel regional, el clima es templado, con una temperatura media anual de 16.3 ºC, precipitaciones medias anuales de 1,023 mm y humedad relativa media anual del aire de 77.6%. El suelo es un argiudol vértico sin sustrato orgánico con horizonte Ap de 0,2 a 12 cm. El cultivo estudiado estuvo bajo manejo convencional de bajos insumos que se practica en quintas de menos de diez hectáreas de superficie, donde se cultivó de forma intensiva, con el uso de insumos químicos limitado, que incluyeron la aplicación por calendario de agroquímicos para el control de malezas (Dicloruro de 1,1'-dimetil-4,4'-bipiridilo y trifluralina), ácaros, pulgones, trips (clorfenapir y malatión) y hongos (penconazol) y fertirriego.
Se realizaron cuatro recolecciones, desde diciembre de 2012 a febrero de 2013, a partir del momento en que las plántulas tenían una altura aproximada de 30 cm y hasta que los tomates se encontraban maduros para cosechar.
Para estudiar la mesofauna, en cada recolección se tomaron 12 muestras de suelo de aproximadamente 300 cm3 a una profundidad de 5 cm. Los oribátidos y colémbolos se extrajeron de las muestras previamente pesadas, mediante embudos de Berlese, a temperatura ambiente y sin luz, durante 10 días. Los individuos fueron contados y separados a nivel de especie o morfoespecie y su abundancia se estimó a través del número promedio de individuos cada 100 g de suelo.
La recolección de arañas y coleópteros se realizó mediante 11 trampas de caída, ubicadas de manera equidistante cada 10 m, a lo largo de las hileras cultivadas. Cada trampa consistió en un recipiente de plástico de 800 ml, 12 cm de altura y 10 cm de diámetro superior. Se utilizó como preservante 350 ml de una solución no atrayente de NaCl saturada, agua y trazas de detergente, como tensoactivo. Los individuos capturados fueron separados e identificados a nivel de especie o morfoespecie. El valor de importancia específico considerado fue el número de individuos promedio por trampa.
Se estimó la riqueza específica, diversidad Shannon-Wiener (H´) y equitatividad de Pielou (J), mediante el programa PAST (ver.3.14) (Hammer et al., 2013).
La estructura de la comunidad de artrópodos estudiada fue definida con base en las especies que estuvieron presentes durante la mayor parte del desarrollo del cultivo, por considerarse más probable que establezcan interacciones. Luego fueron separadas en función de su abundancia estableciéndose cuatro categorías: a) presentes en las cuatro recolecciones reuniendo 10% o más de abundancia relativa promedio por recolección; b) presentes en las cuatro recolecciones y reuniendo menos del 10% de abundancia relativa promedio por recolección; c) presentes en tres recolecciones y reuniendo 10% o más de abundancia relativa promedio por recolección y d) presentes en tres recolecciones reuniendo menos del 10% de abundancia relativa promedio por recolección.
Las especies de coleópteros y de arañas fueron asignadas a grupos funcionales o gremios con base en la bibliografía disponible (según la terminología utilizada por los especialistas en cada grupo taxonómico (Cardoso et al., 2011; Martínez-Ramos, 2008). Los oribátidos y colémbolos fueron considerados en un solo grupo, como mesodetritívoros (se alimentan de restos de tamaño mediano) y complementariamente fueron separados en función de su posición en el perfil del suelo en epi, hemi y euedáficos (Rapoport, 1959). Los colémbolos Entomobriomorpha han sido señalados como característicos del estrato superficial del suelo mientras que los Poduromorpha son reconocidos como habitantes de poros y canalículos del suelo (Bernava-Laborde & Palacios-Vargas, 2000) por lo que la proporción Entomobryomorpha/Poduromorpha se usó como indicadora del estrato del suelo en que los colémbolos se encuentran activos. La proporción de individuos y de especies de oribátidos pertenecientes a las categorías Macropylina, Gymnonota y Poronota se usó como indicadora del nivel de intervención antrópica en el sistema (Aoki, 1983; Shimano, 2011).
Las arañas fueron agrupadas en gremios según la clasificación propuesta por Cardoso et al. (2011) con base en la estrategia de caza y su dieta. Los coleópteros se separaron en grupos funcionales teniendo en cuenta el tipo de alimentación primaria del adulto (Lawrence & Britton, 1994; Moore et al., 2004).
Considerando la pertenencia de las especies a cada una de las categorías descritas arriba, se proponen interrelaciones entre ellas para confeccionar un modelo estructural y funcional.
Resultados y discusión
Mesofauna. Se recolectaron 136 individuos de oribátidos pertenecientes a seis familias y siete especies (Cuadro 1). Aproximadamente el 70% del total de individuos recolectados correspondieron a la especie Scheloribates praeincisus acuticlava Pérez Iñigo & Baggio, 1986, que resultó también la de mayor frecuencia temporal (Cuadro 4). De acuerdo con la proporción entre grupos de oribátidos, el conjunto que habita este suelo hortícola está integrado en su mayoría por especies del grupo Poronota (Balogh & Balogh, 1988), característico de sitios alterados por el hombre (Aoki, 1983). Oppiella nova (Oudemans, 1902), Tectocepheus velatus (Michael, 1880) y Oribatula (Zygoribatula) lata (Hammer, 1961) son especies cosmopolitas, de gran variabilidad morfológica, que han sido mencionadas como especies asociadas a suelos bajo intervención antrópica, comunes en agroecosistemas. Durante el desarrollo del cultivo, la abundancia de oribátidos se mantuvo en valores cercanos a 20 ind/100g de suelo, excepto en la segunda recolección, cuando aumenta el número de especies (Cuadro 2). La diversidad estimada no supero el valor 1 durante todo el desarrollo del cultivo debido al bajo número de especies, y al alto predominio específico deducible de los valores de equitatividad de Pielou (J) (Cuadro 2). Es destacable que los valores de diversidad observados en este cultivo son menores a los observados en parques y plazas de la ciudad de La Plata (Accattoli et al., 2012). Las especies dominantes mencionadas arriba, son comunes en otros sistemas bajo presión antrópica en la provincia de Buenos Aires (Accattoli & Salazar-Martínez 2008,; Salazar-Martínez & De Luca, 2015), aunque forman parte de comunidades de mayor diversidad específica.
Cuadro 1 Lista y ubicación taxonómica de las especies y morfoespecies de la meso y macrofauna recolectadas y su abundancia relativa (%) por grupo y por periodo en el cultivo de tomate.
| Familia | Especie/morfoespecie | Recolección | |||||
|---|---|---|---|---|---|---|---|
| I | II | III | IV | ||||
| Oribatida | Hemileiidae | Hemileius initialis | 8,7 | 4,4 | - | - | |
| Galumnidae | Erogalumna zeucta | 4,4 | - | - | - | ||
| Oppiidae | Brachioppia cajamarcensis | - | 7,3 | - | - | ||
| Oppiella nova | - | 11,6 | 19,1 | 8,7 | **** | ||
| Oribatulidae | Oribatula (Zygoribatula) lata | - | 2,9 | - | - | ||
| Scheloribatidae | Scheloribates praeincisus acuticlava | 82,6 | 72,5 | 80,9 | 82,6 | * | |
| Tectocephaeidae | Tectocepheus velatus | 4,4 | 1,6 | - | 8,7 | **** | |
| Collembola | Hypogastruridae | Morfo sp. 1 | - | - | 2,9 | - | |
| Poduromorpha | Tullbergidae | Morfo sp. 1 | 25,0 | 8,3 | 14,3 | 25,0 | * |
| Brachystomellidae | Morfo sp. 1 | 25,0 | - | - | - | ||
| Entomobryomorpha | Proisotoma minuta | - | 33,3 | 42,9 | 12,5 | ** | |
| Entomobryidae | Entomobrya atrocineta | 25,0 | 8,3 | 2,9 | - | ||
| Morfo sp. 1 | - | 8,3 | - | - | **** | ||
| Morfo sp. 2 | - | - | 5,7 | - | |||
| Lepidocyrtus sp. | 25,0 | - | - | - | |||
| Cyphoderidae | Cyphoda limboxiphia | - | 41,7 | 31,4 | 62,5 | ** | |
| Araneae | Corinnidae | Castianeira sp. | - | 2,9 | - | - | |
| Meriola cetiformis | 5,6 | 4,3 | 3,5 | 1,6 | *** | ||
| Morfo sp. 1 | 3,7 | - | - | - | |||
| Linyphiidae | Agyneta sp. | 9,3 | 4,3 | 7,1 | 4,8 | *** | |
| Scolecura parilis | 5,6 | 4,3 | 1,8 | 6,4 | *** | ||
| Morfo sp. 1 | 5,56 | 20,0 | 6,19 | 3,17 | *** | ||
| Morfo sp. 2 | - | 2,9 | - | - | |||
| Morfo sp. 3 | 1,9 | 8,6 | - | 1,6 | **** | ||
| Morfo sp. 4 | 22,2 | 1,4 | 15,0 | 15,9 | * | ||
| Lycosidae | Hogna sp. | - | 1,4 | 1,8 | - | ||
| Morfo sp. 2 | 27,8 | 11,4 | 21,2 | 25,4 | * | ||
| Tetragnathidae | Glenognatha lacteovitatta | 18,5 | 38,6 | 43,4 | 41,3 | * | |
| Coleoptera | Anthicidae | Morfo sp. 1 | 5,0 | - | - | 3,7 | |
| Carabidae | Aspidoglossa sp. | - | 7,1 | - | - | ||
| Loxandrus sp. | - | - | - | 3,7 | |||
| Scarites sp. | 5,0 | - | - | - | |||
| Pachymorphus striatulus | 2,5 | - | - | - | |||
| Chrysomelidae | Morfo sp. 1 | 15,0 | 7,1 | 3,1 | 11,1 | *** | |
| Coccinelidae | Morfo sp. 1 | - | - | - | 3,7 | ||
| Curculionidae | Baridinae sp. 6 | 2,5 | - | - | - | ||
| Conotrachelus coelebs | - | - | 3,1 | - | |||
| Naupactus leucoloma | 2,5 | - | - | - | |||
| Naupactus xanthographus | - | 7,1 | 3,1 | - | |||
| Phyrdenus muriceus | 32,5 | 14,3 | 18,8 | 22,2 | * | ||
| Elateridae | Morfo sp. 1 | 12,5 | - | 3,1 | - | ||
| Morfo sp. 2 | 10,0 | - | 3,1 | 25,9 | |||
| Morfo sp. 3 | 2,5 | - | - | - | |||
| Morfo sp. 4 | 2,5 | - | 12,5 | - | |||
| Morfo sp. 5 | - | - | 3,1 | 7,4 | |||
| Morfo sp. 6 | - | 21,4 | 3,1 | - | |||
| Histeridae | Morfo sp. 1 | 2,5 | - | - | - | ||
| Lampyridae | Morfo sp. 1 | - | - | 6,3 | - | ||
| Lathridiidae | Morfo sp. 1 | - | - | 6,3 | - | ||
| Nitidulidae | Morfo sp. 1 | - | - | - | 7,4 | ||
| Scarabeidae | Morfo sp. 1 | - | 7,1 | 3,1 | 3,7 | **** | |
| Scydmaenidae | Morfo sp. 1 | - | - | 3,1 | - | ||
| Silvanidae | Morfo sp. 1 | 2,5 | 21,4 | - | - | ||
| Staphylinidae | Anothylus sp.1 | 2,5 | - | - | - | ||
| Apocellus obscurus | - | - | - | 3,7 | |||
| Apocellus opacus | - | - | 3,1 | - | |||
| Meronera sp. | - | - | - | 3,7 | |||
| Rugilus sp. | - | - | 9,4 | 3,7 | |||
| Scopaeus sp. | - | 7,1 | 3,1 | - | |||
| Tenebrionidae | Morfo sp. 1 | - | 7,1 | 12,5 | - | ||
*10% o más de abundancia relativa promedio y 100% frecuencia temporal
**10% o más de abundancia relativa promedio y 75% frecuencia temporal
***Menos del 10% de abundancia relativa promedio y 100% frecuencia temporal
****Menos del 10% de abundancia relativa promedio y 75% frecuencia temporal
Fecha y etapa fenológica I: 28-12-2012 Crecimiento vegetativo, II: 10-1-2013 Floración, III: 24-01- 2013 Fructificacion, IV. 7-2-2013 Maduración
Cuadro 2 Valores estimados de Índices de diversidad por fecha de recolección y grupo estudiado en el cultivo de tomate. S: riqueza específica.
| Recolección | |||||
|---|---|---|---|---|---|
| I | II | III | IV | ||
| Oribatida | S | 4 | 6 | 2 | 3 |
| Shannon-Wienner (H´) | 0,6 | 1,0 | 0,5 | 0,6 | |
| Equidad (J) | 0,5 | 0,5 | 0,7 | 0,5 | |
| Collembola | S | 4 | 5 | 6 | 3 |
| Shannon-Wienner (H´) | 1,4 | 1,4 | 1,4 | 0,9 | |
| Equidad (J) | 1 | 0,9 | 0,8 | 0,8 | |
| Araneae | S | 9 | 11 | 8 | 8 |
| Shannon-Wienner (H´) | 1,9 | 1,9 | 1,6 | 1,6 | |
| Equidad (J) | 0,9 | 0,8 | 0,8 | 0,8 | |
| Coleoptera | S | 14 | 9 | 17 | 12 |
| Shannon-Wienner (H´) | 2,2 | 2,1 | 2,6 | 2,2 | |
| Equidad (J) | 0,8 | 0,9 | 0,9 | 0,9 | |
Los colémbolos estuvieron representados por 59 individuos y nueve especies o morfoespecies que se ubican en cinco familias (Cuadro 1). Las especies con mayor abundancia relativa fueron Cyphoda limboxiphia (Börner, 1913) y Proisotoma minuta (Tullberg, 1871), que concentraron en conjunto aproximadamente el 60% del total de los colémbolos recolectados. Tullbergidae sp. 1 resultó ser también importante al reunir el 15% (Cuadro 1).
La especie C. limboxiphia ha sido reportada como comensal de hormigas alimentándose de sus deyecciones (Rapoport & de Izarra, 1962; Palacios-Vargas, 2014) y ha sido encontrada en agroecosistemas de la provincia de Buenos Aires donde ha sido descrita como regional y propia del suelo de la zona (de Izarra, 1981). Probablemente C. limboxiphia viva a expensas de deyecciones y hongos cultivados en los numerosos hormigueros presentes a lo largo de las melgas de tomate. En este cultivo, se observaron hormigas y pulgones asociados que representaron un 60% y un 10% respectivamente del total de individuos del resto de la macrofauna, excluyendo coleópteros y arañas (datos no publicados).
Proisotoma minuta se alimenta de las hifas de Fusarium oxysporum (Briones et al., 1998) que constituye una plaga del tomate (Misaico, 2015). Aunque no hemos detectado el hongo dañando al cultivo estudiado, éste es común en la zona (Polak & Mitidieri, 2005) y podría estar bajo el control de los agroquímicos utilizados.
La riqueza específica de los colémbolos fue similar a la de oribátidos, sin embargo, su abundancia promedio fue menor y con mayores fluctuaciones durante el período de estudio. Su mayor equitatividad se reflejó en mayores valores de diversidad durante todo el ciclo del cultivo (Cuadro 2).
La proporción Entomobryomorpha/Poduromorpha fue superior a uno en todas las recolecciones, lo que indica mayor actividad de los colémbolos en el estrato superficial del suelo.
La mesofauna tuvo escasa representatividad al comienzo y al final del cultivo con máximos en momentos intermedios, superando los colémbolos a los oribátidos en la tercera recolección.
El pobre registro de oribátidos y colémbolos realizado indica que la mesofauna no constituye un eslabón fuerte en la red trófica del suelo del cultivo de tomate, posiblemente debido al efecto negativo de las prácticas convencionales de manejo, que implican tareas frecuentes de labranza y remoción del suelo, a lo que se suma la aplicación de agroquímicos. Los colémbolos, aunque fueron más diversos podrían encontrarse más estrechamente relacionados con fitófagos como hormigas y pulgones de la red de herbivoría (Fig. 1).
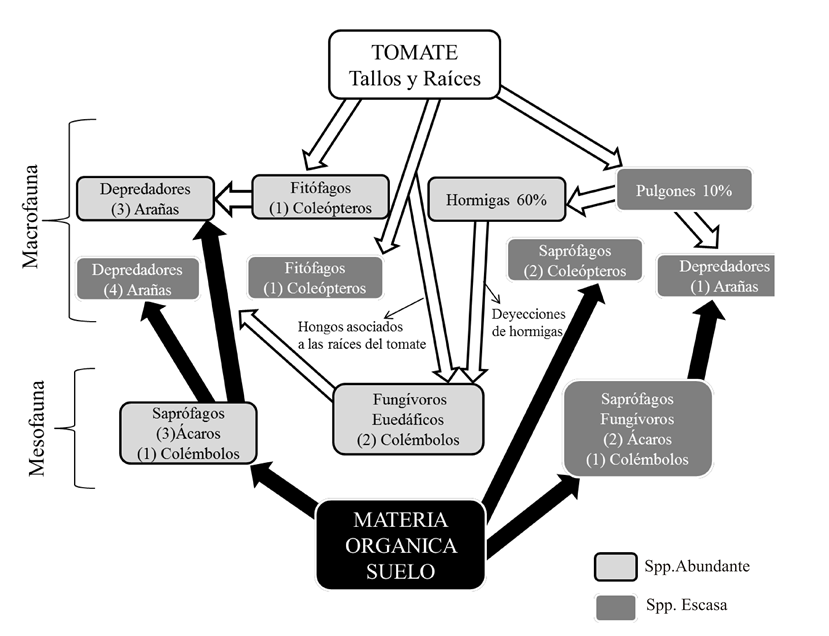
Figura 1 Modelo de la comunidad edáfica estudiada construido con las especies más frecuentes (presentes en al menos tres recolecciones) de arañas, coleópteros, ácaros, oribátidos y colémbolos. (Flechas negras: interrelaciones en la red de detritos. Flechas blancas: interrelaciones en la red de herbivoría. Abundantes: 10% o más del total de individuos recolectados. Escasas: menos del 10% del total recolectado).
Macrofauna: Araneae y Coleoptera. Se recolectó un total de 300 arañas, resultando más numerosas que coleópteros, debido posiblemente a que cuentan con la capacidad de trasladarse a zonas fuera del efecto de las tareas de manejo (refugios) y retornar al cultivo cuando las condiciones son más favorables. Tetragnathidae fue la familia dominante (36.72% del total de individuos recolectados), seguida de Linyphiidae (35.08%), Lycosidae (21.64%) y Corinnidae (4.92%). El número de familias encontradas representa el 25% aproximadamente del total de familias citadas para Argentina (World Spider Catalog, 2017).Esta tendencia a la reducción de las familias en agroecosistemas coincide con los datos obtenidos en estudios realizados en América del Norte (Symondson et al., 2002; Nyffeler & Sunderland, 2003), asociada a la disminución de la diversidad vegetal en los monocultivos. Algunas familias estuvieron representadas solo por una especie, tal es el caso de Glenognatha lacteovitatta (Mello-Leitão, 1944) (Araneae: Tetragnathidae), mientras que Linyphiidae fue la que presentó la mayor riqueza específica. Las especies más importantes fueron G. lacteovitatta, Lycosidae Morfo sp. 2 y Linyphiidae Morfo sp. 4 (Cuadro 1). Durante el desarrollo del cultivo, la diversidad específica mantuvo valores relativamente bajos, con baja dominancia (Cuadro 2). Desde el punto de vista temporal, los resultados obtenidos muestran un incremento en la abundancia del total de la comunidad de arañas a comienzos del cultivo y un descenso de los valores hacia el final del mismo.
El gremio de mayor abundancia fue el de cazadoras (Tetragnathidae, Lycosidae y Corinnidae) y en menor proporción las tejedoras de telas sábana (Linyphiidae) (Cuadro 3). Las primeras presentan dietas menos restringidas que las tejedoras (Turnbull, 1973), capturando presas móviles e inmóviles, en relación con sus estrategias de caza al acecho (Nyffeler et al., 1994; Nyffeler, 1999).
Cuadro 3 Total de especies o morfoespecies recolectadas de cada grupo estudiado por grupo funcional y ubicación en el perfil edáfico. S: riqueza específica, Epi: epiedáfico, Hemi: hemiedáfico, Eu: Euedáfico.
| S | Saprófagos | Fitófagos | Fungívoros | Depredadores | Epi | Hemi | Eu | ||
|---|---|---|---|---|---|---|---|---|---|
| Cazadores | Tejedores | ||||||||
| Oribátidos | 7 | 5 | 2 | 7 | |||||
| Colémbolos | 9 | 8 | 1 | 5 | 4 | ||||
| Coleópteros | 32 | 11 | 6 | 1 | 14 | 32 | |||
| Arañas | 12 | 9 | 3 | 12 | |||||
| Total | 60 | 24 | 6 | 4 | 26 | 44 | 5 | 11 | |
Todos los gremios presentaron tendencias de variación temporal similares a lo largo del cultivo, aumentando progresivamente hacia la etapa final de cosecha. Esta misma tendencia fue observada en colémbolos, que constituyen sus presas habituales (35% de la dieta de algunos licósidos, Chen & Wise, 1999; Nyffeler & Sunderland, 2003). Otras presas comunes son los áfidos, cuyo número disminuye ante su presencia (Chiverton, 1986; Edwards et al., 1979), quienes, como se señaló anteriormente, estuvieron bien representados en las trampas de caída.
En relación con los coleópteros, se recolectaron 121 individuos, pertenecientes a 32 especies o morfoespecies en 15 familias. Curculionidae fue la familia dominante (28.3% promedio del total de individuos), seguida de Elateridae (27.4%), Chrysomelidae, (9.7%) y Staphylinidae (8.5%). El resto de las familias presentó abundancias menores al 5%. Elateridae, Staphylinidae y Curculionidae presentaron, además, la mayor riqueza específica (seis ó cinco especies cada una) (Cuadro 1). Las especies más importantes fueron Phyrdenus muriceus (Gemar, 1824) “el gorgojo del tomate” (22% de abundancia relativa promedio) y Elateridae Morfo sp. 2 (10%).
La cantidad de coleópteros fue menor que la de arañas durante todo el desarrollo del cultivo, sin embargo, su riqueza específica fue generalmente mayor. Durante el período de estudio la diversidad específica mantuvo los valores más altos entre los grupos estudiados, con alta equitatividad, y una alta tasa de recambio temporal de especies. Este resultado podría deberse al efecto de los plaguicidas que disminuyen su cantidad y aumentan la probabilidad de captura de especies diferentes en las trampas de caída (Cuadros 2 y 3).
Se registraron cuatro grupos funcionales (Cuadro 3), en orden de representatividad específica: depredadores (43.7% del total de especies), saprófagos (34.37%), fitófagos (18.75%) y fungívoros (3.1%). El predominio de los primeros marca la importancia del grupo como controlador biológico junto a las arañas. La mayoría de los depredadores fueron estafilínidos, coincidiendo con lo registrado en numerosos cultivos hortícolas (Sengonca et al., 2002). Los detritívoros y fungívoros presentes podrían relacionarse con los abonos adicionados que significan un aporte de materia orgánica. La mayoría de los fitófagos pertenecieron a P. muriceus, reconocida plaga del cultivo de tomate (Lanteri et al., 2002).
Considerando su abundancia, la razón fitófagos/depredadores fue mayor que 1, similar a lo registrado en otros cultivos con manejo convencional (Rouaux et al., 2013). Esto podría asociarse al uso de plaguicidas, que aumenta la abundancia de insectos fitófagos y disminuye la de entomófagos, debido probablemente a que los enemigos naturales son más sensibles a los agroquímicos que sus presas (Zalazar & Salvo, 2007; Nicholls-Estrada, 2008; Strassera, 2009; Fogel, 2012).
De acuerdo con la frecuencia temporal, la abundancia relativa de las especies y su posición en el perfil edáfico, se separó el núcleo de especies más características del suelo del cultivo estudiado, para proponer el modelo de interacciones que se tradujo en la Figura 1 (Cuadros 3 y 4).
Cuadro 4 Especies de mayor frecuencia temporal recolectadas, separadas de acuerdo a su abundancia relativa promedio.
| a | Scheloribates | b | Chrysomelidae Morfo sp. 1 |
| praeincisus | Meriola cetiformes | ||
| Tullbergidae Morfo sp. 1 | Agyneta sp. | ||
| 1 | Scolecura parilis | ||
| Phyrdeneus muriceus | Linyphiidae Morfo sp. 1 | ||
| Glenognatha | |||
| lacteovittata | |||
| Lycosidae Morfo sp. 2 | |||
| Linyphiidae Morfo sp. 4 | |||
| c | Proisotoma minuta | d | Oppiella nova |
| Cyphoda limboxiphia | Tectocepheus velatus | ||
| Entomobrya atrocincta | |||
| Scarabaeidae Morfo sp. 1 | |||
| LinyphiidaeMorfo sp. 3 |
a)100% frecuencia temporal y 10% o más de abundancia relativa promedio por recolección
b)100% frecuencia temporal y menos de 10% abundancia relativa promedio por recolección
c)75% de frecuencia temporal y 10% o más de abundancia relativa promedio por recolección
d)75% de frecuencia temporal y menos de 10% abundancia relativa promedio por recolección
En el modelo propuesto, siete de las doce especies de arañas registradas integran el núcleo central de la comunidad, (Cuadro 4, a y b) mientras que las dos únicas especies de coleópteros presentes son fitófagas, que se alimentan de las plantas de tomate. Las especies dominantes de la mesofauna que permanecen durante la totalidad del desarrollo del cultivo son dos, ambas euedáficas y saprófagas.
El segundo grupo de alta frecuencia temporal (Cuadro 4, c y d) está integrado por cinco especies de la mesofauna, dos de las cuales son colémbolos dominantes posiblemente relacionados con el sistema epigeo: P. minuta que ingiere hifas de hongos fitófagos del cultivo y C. limboxiphia relacionado con las deyecciones de hormigas fitófagas. Las especies menos abundantes del grupo fueron dos colémbolos, un oribátido y un coleóptero saprófago, que podrían relacionarse con la red de detritos iniciada en gran parte por la adición de restos orgánicos al cultivo.
En síntesis, el conjunto estable de la fauna edáfica del sistema estudiado está constituido en su mayoría por depredadores, saprófagos o fitófagos relacionados con el tomate. El modelo de interacciones posible entre estas especies, propuesto en la Figura 1, destaca que en el cultivo estudiado predominan las relaciones en el sistema epigeo respecto al suelo. Esto significa que la mayor parte de los artrópodos considerados se sostienen directa e indirectamente, en base a la energía del cultivo, que es justamente lo que se desea evitar con las prácticas de manejo aplicadas. Sin embargo, la adición de abonos orgánicos que forma parte de las prácticas sostiene el escaso desarrollo de la mesofauna edáfica registrada.
Conclusiones
El grupo más representativo de especies del cultivo estuvo formado por las especies Scheloribates praeincisus acuticlava (Acari: Oribatida) y Tullbergidae Morfo sp. 1 (Collembola) de la mesofauna, que concentraron más del 10% de los individuos y aparecieron en la mayoría de las recolecciones. Respecto a la macrofauna se destacaron Phyrdeneus muriceus (Coleoptera), Glenognatha lacteovittata, Lycosidae Morfo sp. 2 y Linyphiidae Morfo sp. 4 (Araneae).
El modelo de relaciones entre las especies más abundantes y frecuentes que habitan el suelo, evidencia que las especies de la meso y macrofauna integran un grupo que depende más de las plantas de tomate que de la materia orgánica del suelo.

