











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
Las aves acuáticas conforman uno de los grupos de especies más abundantes a nivel continental y son objeto de gran interés en términos ecológicos, económicos y de conservación. Dada su movilidad y sus hábitos migratorios, su manejo resulta complejo. La abundancia de sus poblaciones depende tanto de factores globales como locales y de las condiciones particulares a lo largo de sus rutas de desplazamiento (Pérez-Arteaga & Gaston, 2004). El efecto de estos factores ocurre de forma secuencial, a diferentes escalas y de manera no lineal a través de variaciones ambientales (Almaraz et al., 2012), de tal forma que la construcción de modelos predictivos que faciliten el manejo de estas poblaciones pueden ser materia de investigación en estudios futuros.
Las poblaciones globales de aves acuáticas pueden responder a fenómenos de gran escala como las teleconexiones climáticas (Almaraz & Amat, 2004; Jaksic & Fariña, 2010; Lehikoinen & Jaatinen, 2012; Almaraz et al., 2012; Roach & Griffith, 2015). Diversos estudios han reportado la existencia de índices que han sido utilizados para evaluar la calidad del hábitat (Faaborg et al., 2010; Barker et al., 2014), sin embargo, los objetivos del trabajo y la disponibilidad de la información dadas las condiciones propias del área de estudio limita su implementación. Como una medida confiable se ha asumido que la dinámica de los cuerpos de agua y sus atributos ayuda a explicar la estructura del paisaje (Barker et al., 2014a b).
En tal sentido, estudios realizados sobre grupos de aves durante su estancia invernal o en poblaciones no migratorias, han aportado evidencia de que algunos factores locales son elementos importantes para explicar su distribución y abundancia (Clemente-Sánchez et al., 2014). Por ejemplo Ward et al., (2010), reportaron una reducción en el tamaño poblacional de aves asociada a la modificación de la vegetación en humedales cercanos a zonas en proceso de urbanización. Así mismo, la manipulación de los humedales mediante la reducción o aumento de los niveles de inundación, favorece la presencia de solo ciertos grupos de aves (Parsons, 2002; Bashuk et al., 2012). Por su parte, Austin et al., (2001) encontraron una mayor abundancia de parejas reproductivas de patos de superficie en humedales con una baja cobertura de zonas agrícolas.
Se han reportado incrementos en el tamaño de las poblaciones con respecto a la extensión de los humedales. Por ejemplo, Webb et al., (2010) encontraron una asociación positiva entre el tamaño poblacional de patos de superficie y la disponibilidad de cuerpos de agua semipermanentes adyacentes, mientras que Rosselli y Stiles (2012) documentaron respuestas positivas en la abundancia de rálidos y patos a estas mismas variables.
Santiaguillo es un humedal que representa una oportunidad ideal para estudiar la dinámica de las poblaciones en función de la disponibilidad de hábitats acuáticos. Su ubicación en la parte meridional de las rutas migratorias de anseriformes, su estacionalidad y oscilaciones interanuales, permiten el estudio de las poblaciones en invernación, sujetas a condiciones variables a lo largo del tiempo.
El objetivo de este trabajo fue determinar la respuesta de las poblaciones de aves acuáticas migratorias en estancia invernal a la estructura del paisaje durante la temporada invernal en la Laguna de Santiaguillo, Durango. Por otro lado, su reciente adhesión al convenio de Ramsar desde el año 2012 (RSIS, 2016), requiere de estudios que aporten elementos para el manejo de sus hábitats y poblaciones silvestres. Se prueba la hipótesis de que el tamaño de las poblaciones de aves acuáticas durante la estancia invernal (identificadas por los monitoreos oficiales del USFWS y SEMARNAT) depende de la estructura del paisaje como la forma, la extensión y la distribución que fueron consideradas en la zona de humedales del Valle de Santiaguillo.
MATERIALES Y MÉTODOS
El área de estudio se muestra en la Figura 1 (entre coordenadas 24o 28’ y 24o 60’ N y 104o 36’ y 105o 00’ O). Se encuentra en el centro del estado de Durango, México, e incluye la región conocida como Valle de Santiaguillo, que comprende territorios de los municipios de Nuevo Ideal y Canatlán. Se trata de un sistema de humedales adherido al convenio de Ramsar y considerado como una región prioritaria por la Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. Por su extensión, los principales cuerpos de agua son las lagunas norte y sur de Santiaguillo y la presa San Bartolo, con superficies máximas de 93, 155 y 21 km2, respectivamente. Santiaguillo es un cuerpo de agua de origen natural y acumula los escurrimientos de la cuenca endorreica del mismo nombre; sin embargo, los volúmenes de la laguna norte son retenidos por una cortina de contención establecida desde 1912, causando una inundación persistente (Fig. 1) y limitando la circulación hacia la laguna sur. Por otro lado, diversos cuerpos de agua de menor tamaño, se encuentran alrededor de los vasos principales. Las precipitaciones anuales para el período de estudio, ascienden a 482 mm en promedio y 80 % de ellas ocurren entre junio y septiembre. Por otra parte, la temperatura media anual es de 16.01 oC (CLICOM, 2014). Una síntesis de la precipitación y temperatura media por año se muestra en la Figura 2 en forma de índice de aridez de De Martonne (De Martonne, 1926).
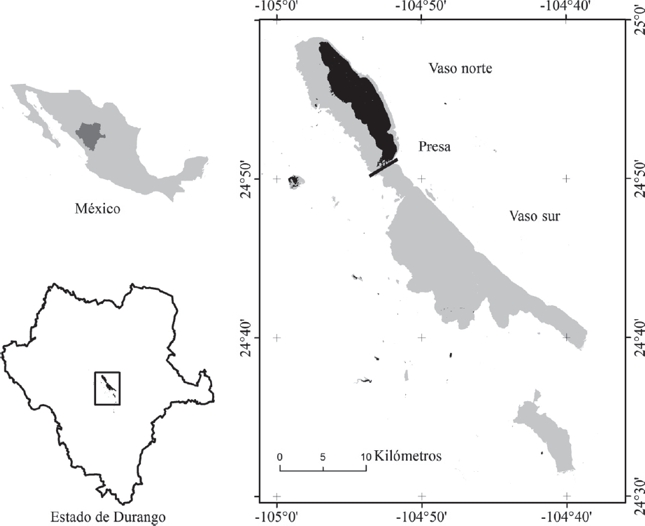
Figura 1 Ubicación de la zona de estudio. Los sombreados negro y gris muestran, respectivamente las extensiones mínima y máxima de los cuerpos de agua, registradas en el período de 1979 a 2014.
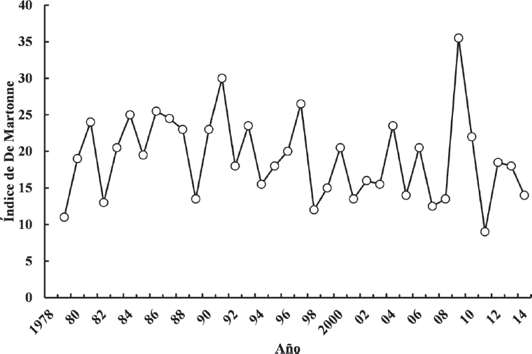
Figura 2 Índice anual de aridez de De Martonne (precipitación media / [temperatura media + 10]) calculado a partir de la información registrada en las estaciones meteorológicas de la zona de estudio.
La extensión y distribución de los cuerpos de agua se determinaron a partir de imágenes LANDSAT (USGS, 2014). Para el período de 1979 a 1986, la información se tomó de imágenes LANDSAT MSS 2 a 5. En el período de 1990 a 1997 y de 2003 a 2011, las imágenes se tomaron del sensor LANDSAT TM 4 y 5. Para el período entre 1999 y 2002 se usaron imágenes ETM+ 7, mientras que en los años 2013 y 2014, se seleccionaron imágenes OLI-TIRS 8. Se usaron preferentemente escenas tomadas en los meses de otoño, coincidiendo con el fin de la temporada de lluvias en la región y con el inicio de la estancia invernal. La presencia de nubes en el sitio de estudio, limitan el uso de imágenes durante el invierno. Consecuentemente se asumió que las condiciones registradas en otoño, son un indicador de las condiciones esperadas en enero.
Las imágenes satelitales fueron sometidas a corrección atmosférica y topográfica mediante el paquete informático ATCOR-3 para ERDAS IMAGINE (Intergraph Corporation). Los cuerpos de agua fueron delimitados mediante el cálculo del índice diferencial normalizado de vegetación (NDVI). Para aumentar el contraste de los ambientes acuáticos con respecto a los terrestres, se usaron las bandas roja e infrarroja media (Salinas et al., 2002). La clasificación de las imágenes correspondientes al año 2014 fue validada con información de campo.
Para el período de 1979 a 2006 se utilizó información poblacional de la base de datos “Mexican Mid-winter Waterfowl Survey” del U.S. Fish and Wildlife Service (USFWS, 2014a), para la región de la Mesa del Norte y área de Santiaguillo (sitio nh-21). Los conteos incluyen información de la abundancia para especies de los géneros Anser, Chen, Anas, Aythya, Bucephala y Oxyura. Los datos por especie de las poblaciones migratorias existentes fueron agrupados en las categorías de gansos (Anser y Chen), patos de superficie (Anas) y patos buceadores (Aythya, Bucephala y Oxyura). Los conteos del USFWS fueron ejecutados durante el mes de enero, a intervalos anuales o trianuales mediante sobrevuelos y avistamientos por tierra. Adicionalmente, para los años 2011 y 2013, se utilizó la información colectada por la Delegación Federal en Durango de la Secretaría de Medio Ambiente y Recursos Naturales (SEMARNAT, 2014) utilizando técnicas y estacionalidades similares a las del USFWS. Una limitación de los datos de poblaciones es que están agrupados para toda la región, lo que reduce las posibilidades de mejorar la resolución espacial.
Las capas resultantes del cálculo del Índice Diferencial Normalizado de Vegetación (NDVI) fueron reclasificadas en las categorías “agua” y “tierra. La categoría “tierra” se consideró como la matriz en el paisaje del área de estudio. Los índices se calcularon a nivel de clase para la categoría “agua” mediante el paquete informático FRAGSTATS 4.2 (McGarigal & Ene, 2014). Para cada año de la serie temporal se calcularon los índices de área total, área promedio, fragmento mayor, número de fragmentos, borde total, densidad de bordes, forma media y proximidad media como indicadores de la complejidad ambiental, con potencial para aportar ventajas al hábitat de las aves acuáticas. El área total corresponde a la suma del área de todos los fragmentos acuáticos, expresada en hectáreas y es un indicador directo de la extensión territorial ocupada por el agua. El área promedio es la media de la extensión de todos los fragmentos y se considera un indicador del tamaño esperado de los cuerpos de agua. El índice de fragmento mayor equivale al área del cuerpo de agua mayor expresada como un porcentaje de toda el área de estudio, considerada como una medida de la dominancia de los ambientes acuáticos sobre los terrestres. El número de fragmentos es la cantidad de cuerpos de agua que pueden distinguirse como entidades individuales y se usó como un indicador del grado de fragmentación de la clase agua. El borde total es la longitud total del límite entre los ambientes acuáticos y terrestres. En el caso del área de Santiaguillo, una mayor longitud de bordes significa una mayor oportunidad para el desarrollo de la vegetación propia de las zonas de transición agua-tierra. La densidad de bordes es la longitud de las zonas de transición agua-tierra por unidad de área, lo que facilita su comparación entre años. La forma media estimada como dimensión fractal, mide la complejidad en la forma de los cuerpos de agua. Su valor oscila entre 1 y 2, para formas simples y formas complejas no euclideanas, respectivamente. De manera similar a la longitud de bordes, las formas irregulares se asocian con la presencia de una mayor diversidad de hábitats. La proximidad media mide la cercanía entre los cuerpos de agua. Su valor es positivo y se incrementa con la cercanía, o bien, tiende a cero cuando no existen vecinos. En el contexto de este estudio, la proximidad puede asociarse con el gasto energético de las aves o el riesgo de depredación durante su desplazamiento. De esta forma la estructura del paisaje resulta un enfoque estratégico para mejorar el entendimiento de la abundancia de las aves, en función de parámetros de configuración espacial, disponibilidad de energía, diversidad de condiciones de hábitat y riesgos de depredación (Barker et al., 2014b).
La respuesta de las poblaciones a la disponibilidad y complejidad de los ambientes acuáticos, se evaluó mediante modelos de la forma X = β0 + β1 X1 + …βn Xn, usando análisis de regresión múltiple Stepwise entre la abundancia de cada grupo de especies (variable dependiente) y los índices de paisaje (variable independiente).
RESULTADOS
La superficie ocupada por los cuerpos de agua, calculada a partir del análisis de las imágenes satelitales, se muestra en la Figura 3. Se observa una pronunciada variación en su extensión, con oscilaciones variables, con un mínimo de 4,000 ha en el año 2001 y un máximo de 29,000 ha en el año 1992, a consecuencia de la dinámica climática e hidrológica de la región. El evento más reciente de máxima acumulación de agua, ocurrió en el año 2008. Los eventos de desecación y expansión de los principales cuerpos de agua, ocurrieron a velocidades y temporalidades distintas (Fig. 3), lo que produjo diferentes patrones de distribución y de extensión, creando distintas condiciones de paisaje cada año. Por ejemplo, el vaso norte conservó persistentemente una extensión de agua importante, mientras que el vaso sur desarrolló independientemente superficies y formas variables e incluso se desecó totalmente en algunos años (Fig. 4).
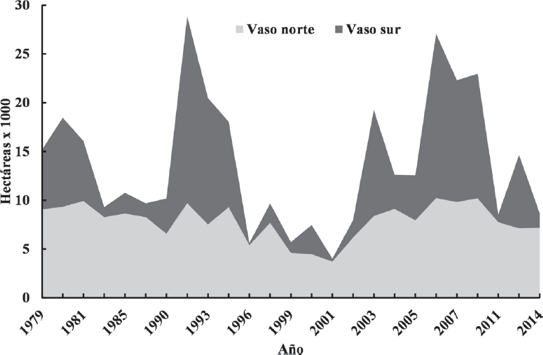
Figura 3 Serie temporal de la superficie ocupada por agua en los humedales de la región de Santiaguillo, Durango. Nota: Los valores se muestran apilados.
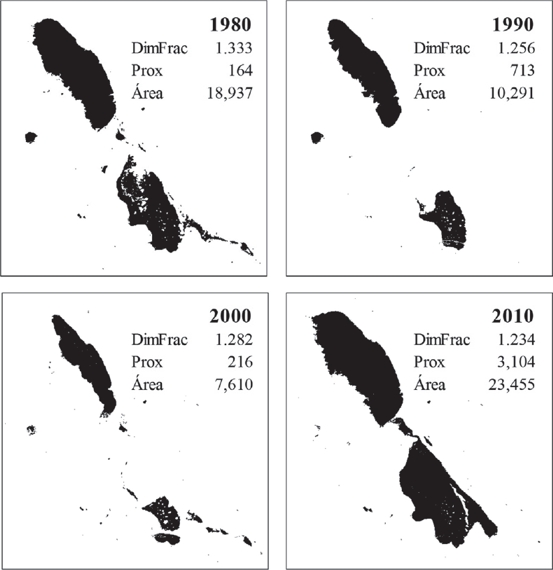
Figura 4 Extensión del humedal en cuatro años diferentes mostrando sus valores de Dimensión fractal (DimFrac), Proximidad (Prox) y Área total (Área). DimFrac tiende a 2 en las formas más complejas, Prox es adimensional y se incrementa con la cercanía, mientras que el Área se expresa en hectáreas.
Cada grupo de aves mostró patrones de abundancia particulares (Fig. 5). Los patos buceadores tuvieron una presencia poco importante en la región, atribuible a la baja profundidad y a la turbidez del agua, propia de esta región y poco adecuada para estas especies; no obstante, entre 1982 y 1986 este grupo registró un incremento poblacional. Los patos de superficie mostraron un patrón similar al grupo anterior, pero con cifras marcadamente superiores entre 1982 y 1986, alcanzando un máximo de 52,000 individuos. Ambos grupos siguieron tendencias históricas distintas a las reportadas por el U.S. Fish and Wildlife Service (2013) para las especies de patos en general, durante la primavera, sugiriendo que estas variaciones podrían obedecer a factores locales.
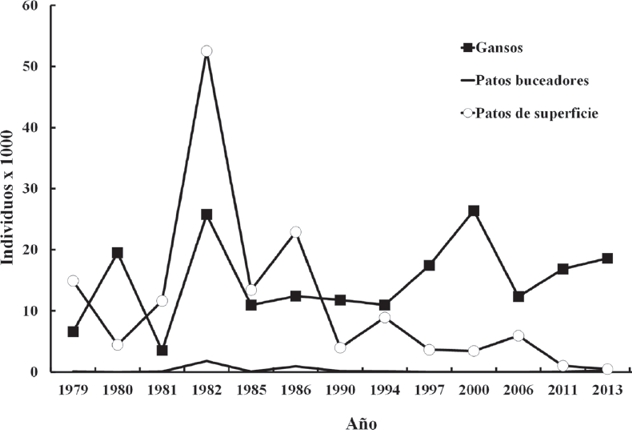
Figura 5 Serie temporal del tamaño poblacional para los grupos de aves acuáticas consideradas en el estudio. Gansos (Géneros Anser y Chen), patos de superficie (Género Anas) y patos buceadores (Género Aythya y similares).
El grupo de gansos incrementó su presencia en la región. Sus poblaciones tuvieron una tendencia histórica positiva y lineal, pasando de 6 mil a más de 20 mil animales entre 1979 y 2013. Este crecimiento coincidió con el fenómeno de incremento global, de tipo exponencial, mostrado especialmente por Anser caerulescens (Linnaeus, 1758) y Chen rossii (Cassini, 1861) (USFWS, 2014b).
Los índices de paisaje calculados mostraron valores independientes y no muestran la misma tendencia que se observa en la superficie ocupada por el agua (Cuadro 1). Esta falta de asociación es atribuible a la diversidad de configuraciones espaciales que el sistema de humedales puede tomar.

El vaso norte presentó inundaciones con extensiones variables, conservando una forma casi constante, gracias a la cortina de contención. Por su parte, el vaso sur mantuvo niveles de inundación variables, pero someras con una mayor exposición a la evaporación, con tendencia a las formas complejas e inestables y con formación de islotes con posibilidades de desarrollar vegetación acuática. Otro elemento que agregó heterogeneidad al paisaje, fueron los cuerpos de agua artificiales menores de uso agropecuario.
Se generó un modelo para el grupo de gansos y otro para los patos de superficie. La información poblacional de patos buceadores no fue suficiente para realizar un análisis apropiado, dada la baja disponibilidad de información poblacional (Cuadro 2).
Cuadro 2 Análisis de regresión múltiple Stepwise, entre la abundancia de cada grupo de especies y los índices de paisaje aceptados por modelo.

En el caso del grupo de gansos, el tamaño de los cuerpos de agua, medido como índice del fragmento mayor tuvo un efecto negativo significativo sobre el tamaño de las poblaciones (R2= 0.71; Pr>F=0.008), mientras que la complejidad de los ambientes acuáticos, medida como índice de la forma media, mostró una influencia positiva (R2= 0.51; Pr>F=0.008) (ver Figura 6).

Figura 6 Modelo gráfico de regresión múltiple de la población invernal de gansos en respuesta a los índices de forma y del fragmento mayor (IFM) (x = - 44479 - 24160 IFM + 62560 Forma).
Por su parte, el grupo de patos de superficie respondió positivamente a la complejidad del ambiente, medida en forma de índice de dimensión fractal basado en la relación perímetro vs área (ver cuadro 2, R2= 0.49; Pr>F=0.006), y negativamente a la proximidad entre cuerpos de agua, medida como la distancia al vecino más cercano (R2= 0.87; Pr>F=0.0003). Es decir, sus poblaciones tendieron a ser mayores cuando las formas de los cuerpos de agua fueron más complejas y las distancias entre los cuerpos de agua fueron más reducidas.
DISCUSIÓN
Notablemente ninguno de los dos grupos respondió positivamente a la superficie total ocupada por el agua. Este resultado fue inesperado y diferente a lo encontrado en otras regiones. Rosselli & Stiles (2012) reportaron mayores densidades de aves acuáticas en los humedales más extensos de la región andina en Colombia; sin embargo, en contraste con el presente trabajo que solo considera especies migratorias, estudiaron especies locales no migratorias.
La topografía del sitio de estudio permite que la profundidad del agua disminuya rápidamente y que una mayor superficie de playa quede expuesta, cuando los humedales se contraen. Esto representa una ventaja en términos de disponibilidad de alimento. Los gansos deben excavar para alcanzar raíces y rizomas, mientras que los patos de superficie sólo pueden alimentarse en zonas someras. Por otro lado, Santiaguillo es el mayor humedal natural de la región y puede mantener cierto nivel de agua durante todo el año, pero es probable que otros cuerpos de agua de menor tamaño se sequen a lo largo del invierno, forzando a las aves a moverse hacia esta zona.
Otros factores pueden explicar la respuesta a la extensión de los humedales. En este trabajo no se evaluó directamente el papel de las comunidades de plantas hidrófilas; sin embargo, en estudios como el de Houlahan et al., (2006), se encontró una mayor riqueza y abundancia de plantas acuáticas en los cuerpos de agua de menor extensión. Esta es una condición similar a lo observado en Santiaguillo, donde la vegetación emergente puede hacer más compleja y atractiva la configuración del paisaje para las aves, especialmente en el vaso sur. Bashuk et al., (2012), manipularon la profundidad de una serie de cuerpos de agua en Manitoba y reportaron una respuesta en el uso del sitio por parte de las poblaciones de patos. Mientras las especies buceadoras respondieron positivamente a un aumento en la profundidad del agua, los patos de superficie lo hicieron de forma negativa. Estas condiciones opuestas pueden estar presentes en Santiaguillo. Por un lado, las aguas profundas son comunes en el vaso norte gracias a una cortina de contención, mientras que ocurre lo contrario en el vaso sur. En este sentido, la permanencia prolongada del agua puede causar la pérdida de heterogeneidad ambiental y diversidad de especies en los humedales sujetos a períodos prolongados de inundación y un efecto inverso en el caso de inundaciones temporales (Parsons, 2002).
Nuestros resultados son concordantes con lo reportado por Medina-Torres et al., (2007), en un estudio sobre el pato mexicano (Anas diazi Ridgway, 1886) en la mesa central Mexicana, donde se detectó un marcado uso de los cuerpos de agua de menor extensión; mientras que los embalses mayores, como las presas, fueron evitados. En su estudio, el uso diferencial se atribuye a la presencia de vegetación acuática. En el caso de Santiaguillo, si bien no se evaluó la presencia de vegetación acuática, se sabe que ciertas especies emergentes, flotantes y sumergidas, son típicas de los cuerpos de agua menores, poco profundos, que ofrecen las condiciones adecuadas para los patos de superficie.
Pearse et al., (2012), encontraron una asociación positiva entre la distribución de patos de superficie y la complejidad del paisaje durante el invierno, en un área de humedales transformados en zonas agrícolas. Nuestro trabajo se llevó a cabo en una región donde la agricultura es también una actividad común; sin embargo, la configuración de los cuerpos de agua, fue suficiente para detectar una relación significativa entre la abundancia de aves acuáticas y la complejidad del paisaje. Ello guarda correspondencia con los objetivos del presente estudio. Sin restar importancia al efecto potencial de los cultivos, nuestro sitio de estudio es diferente en que las tierras de labranza no han invadido los cuerpos de agua, por lo que los ambientes acuáticos pueden diferenciarse aún de las zonas agrícolas.
En el caso de los patos de superficie, nuestros resultados son parcialmente similares a los de Sebastián-González et al. (2010), quienes coincidentemente, documentaron un efecto nulo del tamaño de los cuerpos de agua durante la temporada de invernación. Contrariamente, reportaron una asociación negativa entre el número de aves y la conectividad entre humedales durante la época reproductiva, mientras que en la presente investigación, encontramos evidencia de esta relación durante el invierno. Si bien, nuestros datos están limitados al período de estancia invernal, una explicación a estas diferencias es que solo una de las especies de patos de superficie (A. diazi) en nuestra región, completa su ciclo vital localmente, mientras que el resto son migratorias y se reproducen en regiones boreales. De esta forma, la acumulación de reservas se vuelve importante para la migración de primavera, lo cual puede lograrse con vuelos cortos entre cuerpos de agua cercanos, sin importar su tamaño.
Estos resultados son también concordantes con los reportados por Webb et al., (2010), quienes documentaron una respuesta positiva de las poblaciones de patos de superficie al área de los humedales individuales, a su proximidad y a la cobertura de la vegetación. La correlación entre la población de patos de superficie y el índice de proximidad, denota la importancia de los cuerpos de agua de tamaño reducido. Estos pueden contribuir a la continuidad y cercanía entre los fragmentos, aumentando los puntos potenciales de descanso y reduciendo las distancias de viaje. En otras regiones, las configuraciones paisajísticas de este tipo han demostrado ser importantes en el fomento de poblaciones reproductivas (Naugle et al., 2000).
Para el grupo de gansos, el tamaño de los humedales es una variable determinante; no obstante, el indicador incluido en el modelo de regresión (índice del fragmento mayor), por definición depende únicamente de la extensión del cuerpo de agua más amplio. En este sentido, el grupo responde negativamente solo a los cambios en el tamaño de los cuerpos de agua más evidentes. En nuestro sitio de estudio, una disminución en el tamaño de los vasos principales, podría implicar una mayor superficie de pastizales asociados a sus bordes, donde los gansos pueden alimentarse. Esta forma de respuesta, es un patrón registrado en otros sitios para este grupo, por ejemplo Alopochen aegyptiaca (Linnaeus, 1766) en Sudáfrica (Raeside et al., 2007) y Anser albifrons (Scopoli, 1769) en California (Elphick, 2008).
Un rasgo importante es que el vaso norte de Santiaguillo adquiere siempre formas simples, estables y extensas. En contraparte, el vaso sur toma formas complejas, que varían con su extensión a medida que cambia su nivel de inundación. Esta condición puede contribuir al desarrollo de una mayor diversidad de plantas y hábitat de protección.
La complejidad del paisaje puede atribuirse a la división del humedal, causada por una cortina de contención. Bajo estas condiciones cada uno de los vasos recibe escurrimientos de dos cuencas hidrológicas de diferente extensión y con distintos volúmenes de precipitación. Esto produce tanto ambientes con inundaciones persistentes como áreas de inundación temporal somera que se modifican rápidamente por evaporación.
El contraste de condiciones causado por el manejo actual del sitio, puede producir hábitats apropiados durante el invierno para los grupos de especies evaluados. Adicionalmente, durante la primavera, el vaso norte podría compensar la pérdida de los ambientes someros propios del vaso sur.
No es posible probar el efecto de cada vaso, ya que nuestros datos poblacionales no pueden disgregarse; sin embargo, estudios como el de Kloskowski et al., (2009), han encontrado evidencias de que el manejo marcadamente diferencial en los niveles de agua de humedales contiguos (como sucede en el área de Santiaguillo), genera condiciones contrastantes que complementan los requerimientos de hábitat.
Otros factores no asociados al paisaje, tales como la presencia de cultivos, la calidad del agua, el clima (Chacón-de la Cruz & Pompa-García, 2015), la vegetación acuática y la cacería pueden también explicar la abundancia y distribución de las aves. Ante la falta de datos históricos para estas variables, se recomienda que los trabajos de monitoreo colecten esta información periódicamente. Aunque el uso de imágenes de satélites podría mejorar para análisis espaciales de gran escala (Barker et al., 2014a), en nuestro caso fueron limitados por la falta de correspondencia temporal en la fenología de los cultivos con la presencia de las aves.
CONCLUSIONES
La forma de los cuerpos de agua, su distribución y su extensión, son los factores más importantes para los grupos de aves estudiados en Santiaguillo. Una mayor complejidad de formas y la cercanía entre los cuerpos de agua, sugiere ser un factor importante por el cual parte de las especies de patos usan el sitio durante la estancia invernal. Por otro lado, la disminución en el tamaño de los cuerpos de agua más extensos y el aumento en la complejidad de sus formas favorecen la presencia de especies de gansos.
Los resultados permiten prever oportunidades de manejo, ya que la forma y distribución de los humedales son variables físicas, las cuales pueden manipularse con cierto grado de control, para producir efectos sobre las poblaciones a nivel local.
El historial obtenido mediante las imágenes LANDSAT indica que los humedales del sitio varían cíclicamente y que las variaciones más marcadas ocurren en el vaso sur. Al tratarse de un área suscrita recientemente a la Convención de Ramsar (desde el año 2012; RSIS, 2016), el conocimiento de las relaciones entre el paisaje y las poblaciones silvestres resulta de gran importancia para su manejo, ya sea con fines de conservación o de restauración. La información poblacional corresponde a datos anuales agregados para todo el sitio de estudio, por lo que serán necesarios estudios adicionales que registren la distribución diferencial de las aves sobre el humedal, la evaluación de las variables de hábitat a una escala más fina y el efecto de las actividades humanas.

