



![Diversidad de tisanópteros en crisantemo [Dendranthema grandiflorum (Ramat.) Kitamura] var. Harman en Texcoco, estado de México](/img/pt/prev.gif)







 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
En México, la principal leguminosa de importancia agrícola es el frijol (Phaseolus vulgaris L.), la cual es la principal fuente de proteínas; sin embargo, su producción en los últimos años ha disminuido por ser un cultivo susceptible a condiciones extremas de sequía y alta humedad, así como al ataque de plagas y enfermedades, con lo que se afecta el rendimiento, el cual no supera en promedio la tonelada por hectárea (Servicio de Información Agroalimentaria y Pesquera 2014). Aunado a estos problemas, se presenta el ataque de insectos en granos almacenados, tal es el caso de los gorgojos Acanthoscelides obtectus (Say) y Zabrotes subfasciatus (Boheman), que tienen como hospedero principal a P. vulgaris, en su fase de semilla; aunque también pueden alimentarse de otras especies de leguminosas en condiciones de aislamiento como es el caso de Vigna unguiculata (L.) Walp (National Research Council 2006, Savković et al. 2012, De la Cruz et al. 2013). Los gorgojos de las semillas disminuyen el valor nutricional y la fertilidad de éstas, para lo cual los investigadores han realizado estudios en busca de variedades resistentes de frijol para desarrollar programas de control en este tipo de insectos (Díaz et al. 1996, Sánchez et al. 1997, Ramírez et al. 2003).
Como alternativa para satisfacer la carencia de P. vulgaris, se han realizado estudios en nuestro país para evaluar la fenología, rendimiento y biomasa de V. unguiculata, (conocida como caupí, vigna, frijol chino, chícharo de vaca, yorimuni, yorimón o cowpea en inglés) en condiciones de campo, así como buscar variedades o genotipos con mayor producción que permitan satisfacer las necesidades de proteína vegetal en la alimentación de la población humana (Murillo et al. 1997, Ávila et al. 2005, Wang et al. 2006, Apáez et al. 2014).
Vigna unguiculata es originaria de África, actualmente se siembra en el Continente Americano, principalmente en Brasil, Estados Unidos (California, Texas, Arkansas y estados del sureste) y México (Tamaulipas, norte de Sinaloa, sur de Sonora y Guerrero) (Apáez et al. 2014, National Research Council 2006). En estos estados de México existe el potencial para contribuir en la producción de esta leguminosa que permita favorecer la alimentación de la población, ya que cuenta con mayor rendimiento y valor nutritivo en semilla, ejote y forraje, además posee un alto contenido de proteínas, carbohidratos y fibra, lo cual lo hace un alimento óptimo para el ganado, además de ser un importante regenerador de suelos. Su desarrollo es adecuado en las zonas áridas y semiáridas, donde la escasez de agua y la baja fertilidad de los suelos no favorecen la producción agrícola (Murillo et al. 1997, National Research Council 2006, Olowe et al. 2006, Apáez et al. 2011, Apáez et al. 2014). Sin embargo, algunas plagas en campo provocan la disminución del rendimiento al alimentarse del área foliar y otras ocasionan pérdidas considerables en postcosecha, como es el caso de los brúquidos. El tratamiento químico en almacén es una manera de reducir la alta reproducción de estos insectos y se evitan pérdidas económicas considerables; aunque este tipo de tratamientos puede tener efectos negativos en la salud de los humanos, ante ésto se propone el uso de insecticidas de origen vegetal que disminuyen el riesgo y favorecen el control de los mismos (Southgate 1979, National Research Council 2006, Valenciaga et al. 2007, Abdoul et al. 2014).
Los brúquidos se encuentran distribuidos en casi todos los continentes. El mayor número de especies vive en las regiones tropicales de Asia, África, América Central y Sudamérica. Se alimentan principalmente de semillas de leguminosas y de otras familias de plantas, de las cuales algunas son de importancia económica (Southgate 1979, Salas et al. 2001, Ramírez et al. 2013).
Debido a la importancia de V. unguiculata es necesario estudiar los problemas relacionados con las plagas que afectan a este cultivo, en especial a las especies de brúquidos en postcosecha. De acuerdo con información previa (Jesús Romero Nápoles, comunicación personal), en V. unguiculata se han registrado los siguientes brúquidos: A. obtectus, Callosobruchus maculatus (Fabricius), Decellebruchus atrolineatus (Pic) y Z. subfasciatus, de los cuales no hay datos relacionados con tablas de vida en esta leguminosa, ya que las tablas de vida son útiles para conocer las probabilidades que tiene un individuo de una población determinada de vivir un número X de unidades de tiempo (Méndez et al. 1990) y entender su dinámica poblacional.
El objetivo del presente estudio fue estimar los parámetros poblacionales (tasa neta de reproducción, tasa intrínseca de incremento natural, tasa finita de crecimiento y tiempo de generación) obtenidos de tablas de vida de A. obtectus, C. maculatus (proveniente de dos colonias) y Z. subfasciatus utilizando como alimento semillas de V. unguiculata, para conocer si esta leguminosa es resistente al ataque de estos gorgojos.
Materiales y métodos
La investigación se realizó en el Laboratorio de Ecología de Insectos del Colegio de Postgraduados, Campus Montecillo, Estado de México, en los meses de julio y agosto de 2014. Se emplearon cámaras de cría con condiciones de temperatura de 28 ± 2 °C y 78 ± 2% de humedad relativa, que permitieron la reproducción de las especies A. obtectus, C. maculatus y Z. subfasciatus.
Con relación a C. maculatus se evaluaron los parámetros poblacionales de dos colonias provenientes de diferentes hospederos, la primera se obtuvo de semillas de V. unguiculata (C. maculatus-vigna) colectada en campo en El Fuerte, Sinaloa y la segunda (C. maculatus-cicer), del Laboratorio de Control Biológico del Programa de Entomología y Acarología del Colegio de Postgraduados, Montecillo, Estado de México, que fueron criados en semillas de garbanzo (Cicer arietinum L.). En tanto que A. obtectus y Z. subfasciatus se adquirieron de colonias alimentadas con frijol de la variedad Peruano en el Laboratorio de Ecología de Insectos del Colegio de Postgraduados, con las cuales se establecieron las colonias madre de cada especie y todas se alimentaron con V. unguiculata.
El establecimiento de las colonias madre se realizó dos meses antes del experimento, el cual consistió en limpiar (eliminar granos dañados, rocas y trozos de vainas secas) las semillas de V. unguiculata que sirvieron como alimento de A. obtectus, C. maculatus en sus dos modalidades y Z. subfasciatus. Posteriormente se colocaron en bolsas de plástico y se dejaron por tres días en el congelador, para reducir el daño por insectos distintos a los de interés, ácaros o patógenos. Una vez transcurrido el tiempo de enfriamiento se secaron a temperatura ambiente para posteriormente utilizarlos como alimento de las colonias de los insectos en estudio, los cuales se colocaron en frascos de cristal de 500 mL de capacidad, donde se les incorporaron los adultos de cada especie de brúquido para incrementar la población de los insectos y contar con suficientes especímenes para el experimento.
Para dar inicio al experimento, las tablas de vida de los brúquidos se elaboraron con huevos que se obtuvieron de las crías madre antes mencionadas. Para estimular la oviposición se introdujo un grupo de insectos recién emergidos de cada especie con 300 semillas de V. unguiculata en frascos de cristal con capacidad de 500 mL y tapa cubierta por malla metálica para permitir la ventilación; 24 horas después se retiraron los adultos con lo cual se tuvo la certeza de que los huevos fueron depositados el mismo día y que la proporción hembras: machos fuese aproximadamente 1:1. En un microscopio estereoscópico se seleccionaron al azar las semillas a las cuales se les dejó un huevo (relación 1:1) para evitar competencia intraespecífica por alimento y espacio, de esta manera se formaron cohortes de 200 huevos en cada especie. Este procedimiento se llevó a cabo en C. maculatus en sus dos modalidades y Z. subfasciatus. Para el caso de A. obtectus, el cual no adhiere los huevos a la semilla, se siguió el mismo procedimiento para estimular la oviposición y posteriormente se colectaron 200 huevos al azar que se distribuyeron en igual número de semillas. Las cohortes se dividieron en dos grupos de 100 semillas para facilitar su manipulación. A la semana se registró el número de huevos que eclosionaron y no eclosionaron de cada una de las cohortes, se esperó este tiempo debido a que las larvas emergen entre los cuatro o cinco días y así tener la certeza de que todos cubrieron esta etapa. Para identificarlos se observaron al microscopio estereoscópico contabilizando como no eclosionados a los que no formaron embrión, es decir fueron infértiles y a los que eclosionaron se observó el orificio en el cual las larvas penetraron a la semilla o la larva murió antes de introducirse a ésta. Después de la primera semana se dejó pasar un promedio de 24 días de iniciadas las cohortes hasta que los adultos emergieron de las semillas, debido a que los estados de larva y pupa ocurren en el interior de ésta. Una vez que inició la emergencia de los adultos se llevó el registro de oviposición y mortalidad diaria de cada especie.
La estimación de los parámetros poblacionales en A. obtectus, C. maculatus (en sus dos modalidades) y Z. subfasciatus se realizó con la técnica de tablas de vida de cohorte y la prueba no paramétrica de Log rank (p ≤ 0.05) (Méndez et al. 1990). Así se obtuvieron las curvas de supervivencia (nx ), esperanza media de vida (ex ), emergencia de los adultos, tasas netas de reproducción (R o) (p ≤ 0.05), tasas intrínsecas de incremento natural (rm ) (p ≤ 0.10) (Krebs, 1985), tasa finita de crecimiento (λ), para estimar cuánto se multiplica una población por unidad de tiempo (t + 1) y el tiempo de generación (T) de las especies antes mencionadas con la metodología de Sánchez et al. (1997) y Vera et al. (2002).
Las curvas de supervivencia se construyeron con los datos de las tablas de vida y se compararon con la prueba de Log rank, donde las frecuencias de muerte de cada población se utilizaron para evaluar las discrepancias de las frecuencias observadas (O) con las que se espera ocurran (E, frecuencias esperadas) cuando la hipótesis de nulidad es verdadera. Estas discrepancias se midieron con la X2 cal que se comparó con una X2 0.05 (2 g.l.) de tablas (Méndez et al. 1990). Con los datos de tablas de vida se estimó la esperanza media de vida de cada especie de insectos.
Las tablas de fecundidad (mx, que representa el número de hijas producidas por unidad de tiempo por hembra madre de edad X) y fertilidad se obtuvieron con el registro diario de los huevos depositados por las hembras provenientes de las cohortes de cada especie y de los huevos que fueron fértiles que llegaron al estado adulto. Para ésto, las hembras y machos recién emergidos se colocaron en frascos de cristal de 500 mL con 100 semillas de V. unguiculata (dos frascos por especie para facilitar su manipulación), para que las hembras ovipositaran y cada 24 horas se retiraron las semillas con huevos y se colocaron en cajas Petri de plástico (9 cm de radio) las cuales se reemplazaron con semillas limpias, de esta forma se registró el número de huevos ovipositados cada día hasta que murió el último insecto de las especies en estudio. Las tasas de incremento (rm) se compararon con la prueba de Traslape de Intervalos (p ≤ 0.10) de Vera & Sostres (1991) en cada una de las cohortes en las siguientes combinaciones: A. obtectus vs C. maculatus-cicer; A. obtectus vs C. maculatus-vigna; A. obtectus vs Z. subfasciatus; C. maculatus-cicer vs C. maculatus-vigna; C. maculatus-cicer vs Z. subfasciatus y C. maculatus-vigna vs Z. subfasciatus, para ello, se ajustó el valor de este parámetro a la ecuación de Lotka según Vera et al. (2002). En este caso, se estableció que si los intervalos no se traslapaban después de 33 días (t = 33 d), que es el periodo de tiempo promedio en que se multiplica la población de las especies en estudio, las curvas fueron consideradas estadísticamente diferentes.
Resultados
Las tablas de vida de cada especie se presentan en forma de curvas de supervivencia (Figura 1A). La prueba de Log rank detectó diferencia significativa (p ≤ 0.05) en las cohortes de A. obtectus en sus combinaciones con C. maculatus-cicer, C. maculatus-vigna y Z. subfasciatus (Cuadro 1); el resto de las combinaciones (C. maculatus-cicer vs C. maculatus-vigna, C. maculatus-cicer vs Z. subfasciatus y C. maculatus-vigna vs Z. subfasciatus) no mostró diferencias significativas. Esto se debió a que la mortalidad de los insectos fue mayor en estado de larva, principalmente en Z. subfasciatus donde se presentó el valor más alto (Cuadro 2).

Figura 1 A) Supervivencia B) Fecundidad C) Fertilidad de cohortes de brúquidos alimentados con Vigna uniguiculata.
Cuadro 1 Valores de X2 obtenidos mediante la prueba de Log rank al comparar curvas de supervivencia de los brúquidos.


El número de adultos emergidos (24 a 27 días de edad) fue mayor en C. maculatus en sus dos modalidades seguido de A. obtectus, en cambio Z. subfasciatus resultó con el menor número de emergencia, probablemente debido a que su hospedero primario es el frijol. Por otra parte, con relación a la longevidad máxima C. maculatus-cicer y A. obtectus fueron más longevos en comparación con C. maculatus-vigna y Z. subfasciatus que vivieron menor tiempo (Cuadro 3).

La mortalidad de los adultos de A. obtectus inició a los 29 días y en Z. subfasciatus a los 35 días; sin embargo, la supervivencia de ambas especies se extendió a los 53 y 48 días, respectivamente. Por otra parte, C. maculatus-cicer y C. maculatus-vigna que iniciaron la mortalidad a los 32 días, su ciclo duró 43 y 47 días respectivamente obteniendo una mortalidad menor.
La esperanza media de vida (ex ) de las especies en estudio desde huevo hasta adulto, en general fue similar en los valores obtenidos en A. obtectus y C. maculatus en sus dos modalidades y menor en Z. subfasciatus, específicamente en las primeras etapas de desarrollo (Cuadro 4).
Cuadro 4 Esperanza media de vida (ex)† de brúquidos asociados con Vigna unguiculata.
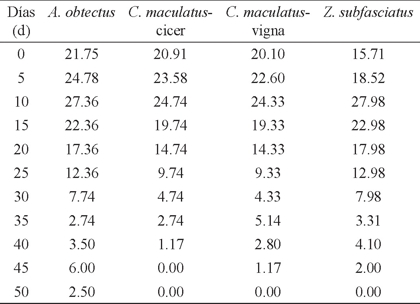
† Valores de cada 5 días.
Con los datos de fecundidad (huevos puestos durante la vida de las hembras) obtenidos (Figura 1B) se puede observar que este parámetro fue mayor en C. maculatus-cicer y menor en Z. subfasciatus. La primera especie fue más fecunda aunque menos longeva. En cambio, C. maculatus-cicer mostró tres ligeros incrementos (30, 33 y 38 días); similar comportamiento presentó C. maculatus-vigna con cuatro picos a los 29, 35, 38 y 42 días, aunque con menor fecundidad que C. maculatus-cicer, lo cual indicó que conforme iban emergiendo los adultos, su capacidad reproductiva fue mayor.
Para el caso de A. obtectus y Z. subfasciatus, mostraron un solo incremento a los 33 días, su periodo fértil culminó al mismo tiempo que las demás especies; cabe destacar, que en estas dos especies, algunas larvas salieron y puparon fuera de la semilla, debido posiblemente al tamaño de éstas, que es más pequeña en comparación con el frijol. La fecundidad de C. maculatus-cicer estuvo por encima de todas las especies; sin embargo, A. obtectus y C. maculatus-vigna presentaron similares valores en este parámetro, lo cual indicó que A. obtectus tiene alta capacidad reproductiva; pero se trata de una especie que no adhiere los huevos a la semilla.
Con relación a la fertilidad (huevos que eclosionaron) C. maculatus-cicer obtuvo el valor más alto, A. obtectus y C. maculatus-vigna presentaron valores similares; sin embargo, Z. subfasciatus fue la menos fértil (Figura 1C).
Las tasas de reproducción de las especies se muestran en el Cuadro 5, donde la R o fue mayor en C. maculatus-cicer y menor en Z. subfasciatus, esto indicó que la primera población se multiplicó 21.7 veces cada 30.16 días, mientras que la segunda se multiplicó 5.36 veces cada 32.40 días. De acuerdo a la R o, el crecimiento poblacional de C. maculatus-cicer es cuatro veces mayor que la segunda, esto se atribuye a que Z. subfasciatus cuando se alimenta con V. unguiculata, por las características de la semilla tiende a tener poca preferencia en su alimentación, ya que su hospedero natural es el frijol; por lo que no ocasionaría un riesgo si se llegase a presentar en condiciones naturales con V. unguiculata.
Cuadro 5 Tasa neta de reproducción (Ro), tasa de incremento natural (rm ), tiempo por generación (T) y tasa finita de crecimiento (λ) de los brúquidos en Vigna unguiculata.
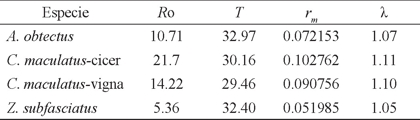
La tasa finita de incremento fue más alta en C. maculatus-cicer con excepción del tiempo por generación que resultó menor, esto es, con una rm = 0.102762 y λ = 1.11, indicó que por cada insecto presente en un momento dado (t), habrá 1.11 en la siguiente unidad de tiempo (t + 1); de igual forma para Z. subfasciatus que obtuvo el menor valor en este parámetro (rm = 0.051985, λ = 1.05) y mostró la tasa más baja de crecimiento, esto indicaría que su presencia en condiciones de campo o en almacén con V. unguiculata no sería tan problemática. En el caso de A. obtectus y C. maculatus-vigna, presentaron valores cercanos, lo cual indicó que tienen casi la misma capacidad para que se multiplique la población; sin embargo, la rm de A. obtectus fue ligeramente menor y con el mayor tiempo generacional, lo cual indica que V. unguiculata también resultó aceptable para esta especie de brúquido (Cuadro 5).
Se encontraron diferencias estadísticas significativas (p ≤ 0.10) de rm con la prueba de Traslape de Intervalos en las siguientes combinaciones: A. obtectus vs Z. subfasciatus, C. maculatus-cicer vs Z. subfasciatus y C. maculatus-vigna vs Z. subfasciatus. No así para A. obtectus vs C. maculatus-cicer, A. obtecus vs C. maculatus-vigna y C. maculatus-cicer vs C. maculatus-vigna. Para entender estas diferencias, teóricamente la población de Z. subfasciatus se incrementaría 5.36% en el transcurso de un día en comparación con C. maculatus-cicer que sería de 21.7 %; A. obtectus con 10.71 % y C. maculatus-vigna 14.22 %, lo cual indicó que teóricamente Z. subfasciatus y A. obtectus pueden alimentarse de granos de V. unguiculata en condiciones de almacenamiento y podrían convertirse en plagas de importancia económica.
Discusión
Con los datos de las tablas de vida se pudo observar la supervivencia de cada una de las especies, donde A. obtectus y Z. subfasciatus mostraron resultados similares a los obtenidos por Ramírez et al. (2003), Rentería et al. (1997) y Sánchez et al. (1997) quienes encontraron una duración similar del ciclo de vida en variedades de frijol y además una alta mortalidad de larvas, lo cual lo atribuyeron a la dureza de la testa y a tóxicos en esta estructura.
Los adultos emergidos y la longevidad en cada una de las cohortes fue mayor en las dos modalidades de C. maculatus debido a que V. unguiculata al ser su hospedero primario no tiene dificultad para alimentarse de sus semillas. A. obtectus a pesar de no adherir los huevos presentó una emergencia similar a C. maculatus, lo cual indicó que tiene potencial para convertirse en plaga de granos almacenados, situación que no ocurrió con Z. subfasciatus, ya que esta especie se alimenta principalmente de frijol; sin embargo, aun con la baja emergencia puede convertirse en plaga local (Jarry & Bonet 1982, Savković et al. 2012).
Ahora bien, la mortalidad de larvas fue mayor en Z. subfasciatus, esto quizá se deba a las características físicas (dureza) y de toxicidad de la semilla, ya que como lo menciona Ramírez et al. (2003) en variedades resistentes de frijol encontró que estos insectos presentaron alta mortalidad.
Con relación a la esperanza media de vida se podría decir que quien tiene menor esperanza de vida sería el más resistente; sin embargo, este parámetro no necesariamente indicaría resistencia, ya que lo más confiable son los valores de las tasas de reproducción y el tiempo generacional de cada especie, donde se obtiene la población potencial de un insecto (Sánchez et al. 1997); sin embargo, A. obtectus y Z. subfasciatus obtuvieron tasas de reproducción bajas y el tiempo generacional similar en ambas especies; no así para C. maculatus en sus dos modalidades, que presentaron mayor tasa reproductiva y menor tiempo generacional, lo cual indicó que tanto A. obtectus como Z. subfasciatus tienen potencial para convertirse en plagas de V. unguiculata (Davis 1972, Jarry & Bonet 1982, Meik & Dobie 1986).
Respecto a la fecundidad, es importante mencionar que C. maculatus en sus dos modalidades distribuyó los huevos en las semillas casi de forma uniforme (se contabilizaron hasta 13 huevos por grano) presentando en promedio tres picos reproductivos, lo cual coincide con lo señalado por Thanthianga & Mitchell (1990) quienes encontraron que la fecundidad de C. maculatus se incrementó cuando emergen después de los 31 días y además pueden distribuir los huevos en las semillas de tal forma que un grano puede soportar hasta 15 o más larvas. Para el caso de A. obtectus y Z. subfasciatus, mostraron únicamente un incremento, el cual inició y terminó casi al mismo tiempo, donde los valores que alcanzó A. obtectus fueron similares a los presentados por C. maculatus-vigna, lo cual muestra su capacidad reproductiva. Sin embargo, a pesar de su buena fecundidad, su desventaja es la alta mortalidad de las larvas, ya que al no adherir los huevos a los granos mueren en la búsqueda de éstos o incluso se enfrentan a las características físicas (dureza) y de toxicidad de la semilla, lo cual controla el comportamiento de penetración de las larvas (Thiéry et al. 1994). Es importante señalar, que aun cuando las larvas de A. obtectus tienen la ventaja de elegir las semillas, éstas dejaron un 40% de granos sin invadir de los 200 que conformaron la cohorte, lo cual podría deberse a la dureza de la testa o tal vez a que algunas utilizaron el mismo punto de entrada a la semilla o quizá a la poca preferencia que tienen por esta leguminosa. Al respecto, Ramírez et al. (2003) señalaron que las larvas de primer estadio de A. obtectus presentan la mayor mortalidad al no penetrar la testa de la semilla de frijol en variedades resistentes.
En la fertilidad, se observó que a pesar de que C. maculatus en sus dos modalidades presentó la mayor fecundidad, su fertilidad disminuyó considerablemente debido a que hubo una gran cantidad de huevos infértiles, no así para A. obtectus que tanto la fecundidad como la fertilidad se mantuvieron con similar valor, lo cual indicó que su capacidad reproductiva es buena; sin embargo, la alta mortalidad de larvas afecta considerablemente su supervivencia y en el caso de Z. subfasciatus, presentó los valores más bajos lo cual podría atribuirse a la poca preferencia que tiene para alimentarse de las semillas de V. unguiculata.
Referente a los parámetros poblacionales se encontró que C. maculatus-cicer en general obtuvo mayores valores en comparación con Z. subfasciatus que presentó los más bajos, esto se puede atribuir a su origen, debido a que se trata de plagas que vienen de regiones completamente aisladas y distintas, ya que la primera especie es nativa del Viejo Mundo y la segunda es originaria de América (Southgate 1979, Silva et al. 2004); sin descartar que Z. subfascitus en ausencia de frijol llega a alimentarse de especies de Vigna (Davis 1972, Meik & Dobie 1986). Para A. obtectus y C. maculatus-vigna su comportamiento fue similar, la primera es estenófaga y está asociada principalmente al frijol; sin embargo, se puede alimentar o invadir otros granos en condiciones controladas o de almacén (Jarry & Bonet 1982, Savković et al. 2012) y la segunda es polífaga y además V. unguiculata es su hospedero natural (Thanthianga & Mitchell 1990); sin embargo, tanto A. obtectus como Z. subfasciatus en ausencia de frijol pueden alimentarse de V. unguiculata, ya que son capaces de ocasionar pérdidas en postcosecha, lo cual las puede convertir en plagas de importancia económica como lo es C. maculatus (Davis 1972, Jarry & Bonet 1982, Meik & Dobie 1986).
En estudios realizados por Díaz et al. (1996), Sánchez et al. (1997) y Ramírez et al. (2003) relacionados con la supervivencia y fecundidad de A. obtectus y Z. subfasciatus en diferentes variedades de frijol, encontraron datos superiores en los parámetros de la R o (7.5 a 12.72) y rm (0.05351 a 0.0860) en comparación con los obtenidos en V. unguiculata; sin embargo, en el tiempo de generación (T) estos autores obtuvieron valores menores para Z. subfasciatus (28.93 a 29.56 días) y mayores para A. obtectus (36.8 a 38.6 días) en frijol, en cambio en V. unguiculata se presentó una duración similar del tiempo generacional con C. maculatus en sus dos modalidades con valores de 32.97 y 32.40, respectivamente (Cuadro 5). En la fecundidad en frijol, encontraron que A. obtectus, presentó un menor valor y Z. subfasciatus un mayor valor, a los obtenidos en V. unguiculata donde C. maculatus-cicer mostró el dato más alto en este parámetro, A. obtectus y C. maculatus-vigna presentaron similar comportamiento y en Z. subfasciatus la fecundidad más baja, lo que podría indicar que las semillas de V. unguiculata no son adecuadas para el óptimo desarrollo de este brúquido, quizá por la dureza de la testa; sin embargo, si llegan a alimentarse de ellas.
Con relación a C. maculatus, el daño que provoca a las semillas de garbanzo (C. arietinum L.) en condiciones de almacén es muy alto y podría equipararse al daño que este brúquido ocasiona también a V. unguiculata (Moreno et al. 2000, Erler et al. 2009, Panzarino et al. 2011); aunque faltaría realizar estudios complementarios relacionados con el comportamiento en campo, para evaluar las pérdidas que ocasionan estos insectos en V. unguiculata y con ello, generar un programa de manejo integrado.
La prueba de Traslapo mostró que teóricamente las poblaciones de C. maculatus en sus dos modalidades se incrementan con mayor rapidez en comparación con A. obtectus y Z. subfasciatus, lo que implica tomar medidas preventivas cuando se presenten estos insectos en condiciones de almacenamiento.
Conclusiones
Con los datos de supervivencia y los parámetros poblacionales se concluye que V. unguiculata es susceptible al ataque de C. maculatus en condiciones naturales y de almacenamiento, ya que al igual que C. arietinum son hospederos primarios de este brúquido.
Acanthoscelides obtectus y Z. subfasciatus al alimentarse de V. unguiculata en condiciones de almacenamiento, tienen potencial para convertirse en plagas de importancia económica en ausencia de frijol.

