










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkActa zoológica mexicana
versión On-line ISSN 2448-8445versión impresa ISSN 0065-1737
Acta Zool. Mex vol.31 no.3 Xalapa dic. 2015
Artículos originales
Morfología del sistema reproductor de la hembra de Comadia redtenbacheri (Hammerschmidt) (Lepidoptera: Cossidae)
Morphology of the female reproductive system of Comadia redtenbacheri (Hammerschmidt) (Lepidoptera: Cossidae)
Arturo Ramírez-Cruz1 y Celina Llanderal-Cázares2
1 Instituto Politécnico Nacional. Centro de Educación Continua, Unidad Morelia. C.P. 58190. Morelia, Michoacán, México. <aramirezc@ipn.mx>
2 Colegio de Postgraduados. Entomología y Acarología, Campus Montecillo. C.P. 56230. Montecillo, Estado de México. <llcelina@colpos.mx>
Recibido: 22/04/2015.
Aceptado: 16/07/2015.
Resumen
Debido a su importancia económica y con la finalidad de profundizar en el conocimiento sobre la biología reproductiva del gusano rojo del maguey Comadia redtenbacheri (Hammerschmidt) (Lepidoptera: Cossidae), se determinó la morfología del sistema reproductor de la hembra de esta especie, el cual presenta en general las características básicas de otros Lepidoptera, con algunas particularidades específicas. La fecundidad potencial tuvo un valor promedio de 104 ovocitos por hembra. En la base de la bursa copulatrix se presentó un pequeño divertículo o lóbulo, y dentro de ella se encontró un solo espermatóforo. La espermateca presentó tanto un utrículo como una lagena. La porción glandular de las glándulas accesorias es inusualmente larga en comparación a otros Lepidoptera.
Palabras clave: gusano rojo, ovariolas, glándulas accesorias, bursa copulatrix, fecundidad potencial.
Abstract
Due to the economic significance of the agave red worm Comadia redtenbacheri (Hammerschmidt) (Lepidoptera: Cossidae), and with the objective of increasing the knowledge about its reproductive biology, the morphology of the female reproductive system was studied. In general, it is similar to that of other Lepidoptera, with a few differences. The average potential fecundity was 104 oocytes per female. A small diverticulum or lobe was present in the base of the bursa copulatrix; inside the bursa, a single spermatophore was found. The spermatheca has a utriculus and a lagena. The glandular portion of the accessory glands is unusually long compared with other Lepidoptera.
Key words: red worm, ovarioles, accessory glands, bursa copulatrix, potential fecundity.
INTRODUCCIÓN
Comadia redtenbacheri, conocido como gusano rojo del maguey, es un barrenador de agaves utilizado en su etapa larvaria como alimento en la gastronomía tradicional, por lo que constituye una fuente de ingreso económico temporal para habitantes de comunidades ubicadas en regiones semiáridas y áridas del centro de México, en donde sus poblaciones silvestres son intensivamente colectadas (Miranda-Perkins et al. 2013). Como sucede con otros insectos comestibles en diferentes países del mundo, a pesar de su importancia se conoce poco de tales especies, lo que dificulta que se consideren dentro de programas de manejo y aprovechamiento, para lo que se requiere del conocimiento de su distribución, biología, comportamiento, estacionalidad y su papel en los ecosistemas, entre otros aspectos, que permita la conservación tanto de los insectos como de sus hospederos (Yen 2009, 2012).
La información sobre aspectos reproductivos de Cossidae incluye la descripción de la estructura reproductiva externa principalmente de los machos, que es usada en Taxonomía, mientras que en los reportes relacionados con las hembras se hace mención principalmente a características del oviducto, de las papilas anales y de la bursa copulatrix, pero las hembras de varias especies son desconocidas (Dugdale 1974; Yakovlev 2006, 2008; Borth et al. 2011; Ivinskis et al. 2012). Otros estudios relacionados con la reproducción en esta familia se refieren al comportamiento de apareamiento y a la proporción sexual de Prionoxystus robiniae, un barrenador de especies forestales (Solomon & Neel 1973; Solomon 1976).
Con relación a la reproducción en C. redtenbacheri, Camacho et al. (2003) observaron adultos durante el cortejo y el apareamiento, y señalaron que las hembras ovipositan sobre cualquier sustrato disponible y mueren pocas horas después y que incluso las hembras que no se aparean pueden depositar huevos infértiles. Llanderal-Cázares et al. (2007) observaron que el apareamiento se inicia alrededor de las 19:00 horas y que las hembras que llegan a copular depositan los huevos en masa, unidos por la secreción de las glándulas accesorias, determinando que la fecundidad potencial es de 118 huevos por hembra. Miranda-Perkins & Llanderal-Cázares (2013) detectaron diferencias de fecundidad en cruzas con diferente proporción de sexos y concluyeron que en cruzas con igual proporción de hembras y machos se obtienen las mayores oviposturas, mientras que en aquellas con un macho y dos o tres hembras, la descendencia disminuye.
Debido a su importancia económica y con la finalidad de profundizar en el conocimiento sobre su biología reproductiva, este estudio tuvo como objetivo el determinar la estructura del sistema reproductivo de la hembra de C. redtenbacheri.
MATERIALES Y MÉTODOS
El estudio se inició con larvas extraídas del rizoma de agaves infestados, en septiembre de 2012 en los municipios de Tulancingo y Tizayuca, en el Estado de Hidalgo. Las larvas con peso mayor a 0.40 g fueron colocadas individualmente para la pupación en un sustrato de suelo y vermiculita a partes iguales, contenido en tubos de poliducto que se mantuvieron en oscuridad total a 20 ± 5 °C y 40-50% de humedad relativa, de acuerdo al procedimiento descrito por Miranda-Perkins et al. (2013). Las pupas se obtuvieron en el mes de marzo de 2013 y las hembras emergieron un mes después.
Para la caracterización morfológica del sistema reproductor de la hembra, se emplearon cuatro pupas próximas a eclosionar y nueve hembras adultas de dos días de edad, las cuales se disecaron en solución de Ringer para insectos para extraer completos los sistemas reproductores; se fijaron en líquido de Carnoy (Martínez 2002) y finalmente se enjuagaron en alcohol de 70%. Para la búsqueda de espermatóforos, se utilizaron más de 60 hembras adultas de uno a dos días de edad, tiempo suficiente para que ocurriera el apareamiento, que puede tener lugar pocas horas después de la emergencia; sus sistemas reproductores se disecaron, fijaron y enjuagaron como ya se mencionó y finalmente se transparentaron sumergiéndolos en una solución de glicerina en alcohol de 70% (1:1) hasta su utilización (Gurr 1956). Las observaciones de cada uno de los sistemas reproductores se realizaron en fresco con un estereomicroscopio VE-S1 marca Velab® provisto de una escala milimétrica adaptada a la platina del estereomicroscopio. Se obtuvieron imágenes digitales mediante una cámara Olympus®, un fotomicroscopio Carl Zeiss Tessovar® y una cámara digital Pixera® profesional.
RESULTADOS
El sistema reproductor de la hembra de C. redtenbacheri (Fig. 1A) presenta un par de ovarios, cada uno con cuatro ovariolas de tipo politrófico. Como en hembras de otras especies de insectos, cada ovariola de C. redtenbacheri presenta el germario, el vitelario y el pedicelo (Figs. 1A y 1B). El germario es pequeño y de forma ligeramente ovalada; el vitelario por su parte, es muy largo y contuvo en promedio 13 ovocitos con corion (que han concluido la vitelogénesis) (Fig. 1A) incluso en las pupas próximas a emerger. Considerando las ocho ovariolas contenidas en ambos ovarios, cada una en promedio con 13 ovocitos maduros, se tiene que la fecundidad potencial de C. redtenbacheri fue en promedio de 104 ovocitos por hembra.
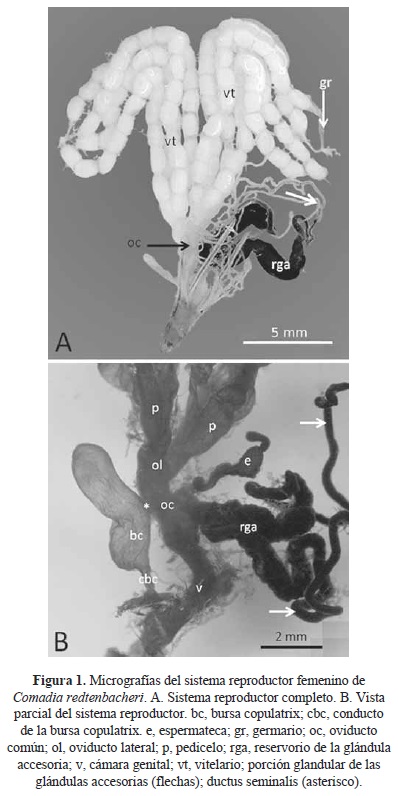
El pedicelo de cada ovariola es membranoso y tiene una forma sacular; los cuatro pedicelos de cada ovario desembocan finalmente en el oviducto lateral (Fig. 1B). Los dos oviductos laterales son cortos y se comunican con el oviducto común, también corto. El oviducto común finalmente conecta a la cámara genital (Fig. 1B). Hacia el oviducto común desemboca la bursa copulatrix, mediante un ductus seminalis corto (Fig. 1B), la cual es membranosa, de forma cilíndrica y basalmente presenta un pequeño divertículo o lóbulo, también membranoso (Fig. 2A).
En el interior de la bursa copulatrix de sólo un ejemplar disecado, se pudo encontrar un solo espermatóforo, cuyo cuerpo fue de color blanquecino y ligeramente piriforme con una longitud aproximada de 1.5 mm y con collum de alrededor de 1 mm (Fig. 2D). La bursa copulatrix finalmente se conecta hacia el exterior del cuerpo mediante su conducto que termina en la ostia como abertura secundaria y que sirve para la cópula (Fig. 2A).
En C. redtenbacheri cerca del ductus seminalis de la bursa copulatrix, se conecta la espermateca al oviducto común por medio de un conducto relativamente corto (Fig. 1B), misma que se conforma de dos lóbulos, el principal o utrículo y otro pequeño y sacular o lagena. El utrículo finalmente se une por su parte apical a la glándula de la espermateca, corta y tubular (Fig. 2C).
Hacia el oviducto común también confluyen un par de glándulas accesorias (o colaterales) por medio de un conducto impar relativamente largo. Las glándulas accesorias son muy prominentes y están formadas de un reservorio glandular tubular de gran anchura, donde se almacena la secreción (Figs. 1A, 1B y 2B). En este reservorio desemboca la porción glandular, que se distingue del reservorio por estar formada de un tubo muy delgado de gran longitud, donde se produce la secreción, el cual llega a medir más de 55 mm (Fig. 2B). La secreción de las glándulas accesorias es de color negro intenso, dando a toda la glándula y sus reservorios esa coloración, incluso en las pupas próximas a eclosionar.
DISCUSIÓN
Comadia redtenbacheri presentó ocho ovariolas de tipo politrófico, cantidad que se presenta en la mayor parte de Lepidoptera, mientras que el ovario politrófico es una característica básica en todos los miembros de este Orden (Kristensen 2003).
La fecundidad potencial de C. redtenbacheri fue en promedio de 104 ovocitos; esta cantidad es cercana a la fecundidad potencial de 118 huevos reportada por Llanderal-Cázares et al. (2007) para esta especie. El hecho de que las hembras de C. redtenbacheri presentaran ovocitos con corion desde que emergen, ubica a la especie como proovigénica, característica de gran importancia, ya que su etapa adulta es de muy corta duración, la cual según Llanderal-Cázares et al. (2007) es de sólo tres a cinco días, y además durante este tiempo no se alimenta por tener atrofiado su aparato bucal. De esta forma, la proovigenia capacita a la hembra a emerger con ovocitos maduros listos para ser fertilizados y ovipositados en el transcurso de horas. Llanderal-Cázares et al. (2007) indican en este sentido, que en C. redtenbacheri el apareamiento puede tener lugar el mismo día de la emergencia y la oviposición presentarse al siguiente.
En general en los insectos la proovigenia puede no ser del todo favorable, ya que como en las hembras proovigénicas todos los nutrientes para los huevos son adquiridos durante el estado larvario y éstas emergen con una cantidad significativa de ovocitos maduros, tienen limitaciones para dispersarse antes de la oviposición, ya que la cantidad de energía disponible para su vuelo es limitada (Riddick 2005; Pélisson et al. 2012).
Comadia redtenbacheri presentó en la bursa copulatrix un pequeño divertículo o lóbulo basal. Con relación a lo anterior, Kristensen (2003) señala que cuando en Lepidoptera la bursa copulatrix presenta un lóbulo extra situado en el extremo posterior (adyacente al conducto de ésta) algunas veces se le denomina como pseudobursa o appendix bursae. Sin embargo, en este trabajo con C. redtenbacheri, no se puede asegurar que el divertículo presente en su bursa copulatrix, pudiera estar relacionado con la estructura mencionada, por lo que se requiere profundizar en ello. El sistema reproductor de la hembra de C. redtenbacheri es del tipo Ditrysia, ya que la bursa copulatrix y el oviducto común están conectados entre sí por medio del ductus seminalis, característica que según Dugdale (1974) y Kristensen (2003) comparten la mayor parte de los Lepidoptera.
El hecho de haber encontrado un solo espermatóforo en la bursa copulatrix de C. redtenbacheri, sustentaría lo señalado por Miranda-Perkins & Llanderal-Cázares (2013), quienes sugirieron que por su corta longevidad y su incapacidad de alimentarse, los adultos de C. redtenbacheri copularían una sola vez. Lo anterior es parecido a lo que sucede en la palomilla Prionoxystus robiniae Peck (Cossidae), donde las hembras son monogámicas y los machos copulan sólo una vez en el 90% de los casos (Solomon & Neel 1973). Con relación a esto, Drummond (1984) indica que en Lepidoptera, el macho normalmente sólo transfiere un espermatóforo durante la cópula. Aunque en general en Lepidoptera la bursa copulatrix tiene la función principal de recibir el espermatóforo (Kristensen 2003), en el caso de la palomilla Illiberis rotundata Jordan (Zygaenidae) puede funcionar además como un depósito de huevos provenientes del oviducto común, que al reabsorberse sirven como fuente de nutrientes para la supervivencia de las hembras (Koshio et al. 2002).
La espermateca de C. redtenbacheri está conformada de un utrículo y de una lagena, pero se desconoce la posible función de estas estructuras en esta especie, ya que en Lepidoptera, tanto el utrículo como la lagena varían tanto en forma como en función, sobre todo con relación a la recepción de los espermatozoides eupirénicos y apirénicos, ya que, por ejemplo, en Diatraea saccharalis (F.) (Crambidae) ambos tipos de espermatozoides entran tanto en el utrículo como en la lagena (Miskimen et al. 1983), mientras que en Spodoptera litura (F.) (Noctuidae) ambos tipos se llegan a encontrar sólo en el utrículo pero nunca en la lagena (Etman & Hooper 1979).
En C. redtenbacheri la parte glandular de las glándulas accesorias es extremadamente larga, por lo que contrasta con la longitud en otras especies con glándulas accesorias con morfología semejante, ya que por ejemplo en los Noctuidae Sesamia nonagrioides (Lef.) dicha parte glandular llega a medir sólo 10 mm (De Santis et al. 2008), mientras que en Spodoptera exigua (Hubner) mide tan sólo 5 mm (Amaldoss & Hsue 1989) y en S. litura (F.) mide 28 mm (Etman & Hooper 1979). Llanderal-Cázares et al. (2007) indican que dicha secreción en C. redtenbacheri sirve tanto para unir los huevos unos a otros durante la oviposición, como para adherirlos a la base del maguey, su planta hospedera. Sin embargo, en S. nonagrioides, que presenta glándulas accesorias morfológicamente semejantes a las observadas en C. redtenbacheri, se sabe que la secreción de dichas glándulas puede funcionar además como una kairomona (De Santis et al. 2008).
Es necesario profundizar en el estudio sobre la estructura y el funcionamiento del sistema reproductor femenino de C. redtenbacheri, con la finalidad de conocer más acerca de la morfofisiología reproductiva de esta importante especie.
AGRADECIMIENTOS
Al Consejo Nacional de Ciencia y Tecnología, por el apoyo brindado a través del proyecto 166898, "Biología de Comadia redtenbacheri Hamm.". A los revisores anónimos por sus valiosos comentarios.
LITERATURA CITADA
Amaldoss, G. & Hsue, N. 1989. The biology and the reproductive morphology of beet army worm Spodoptera exigua (Hubner) (Lepidoptera: Noctuidae). Chinese Journal of Entomology, 9: 239-250. [ Links ]
Borth, R., Ivinskis, P., Saldaitis, A. & Yakovlev, R. 2011. Cossidae of the Socotra Archipelago (Yemen). Zookeys, 122: 45-69. [ Links ]
Camacho, A. D., Sánchez H., A., Jiménez L., J. E. & Nolasco M., A. 2003. Observaciones en condiciones de laboratorio de la biología del gusano rojo de maguey Comadia redtenbacheri H. (Lepidoptera: Cossidae). Entomología Mexicana, 3: 281-287. [ Links ]
De Santis, F., Conti, E., Romani, R., Salerno, G., Parillo, F. & Bin, F. 2008. Colleterial glands of Sesamia nonagrioides as a source of the host-recognition kairomone for the egg parasitoid Telenomus busseolae. Physiological Entomology, 33: 7-16. [ Links ]
Drummond III, B. A. 1984. Multiple mating and sperm competition in the Lepidoptera, pp. 291-370. In: Smith, R. L. (Ed.). Sperm Competition and Evolution of Animal Mating Systems. Academic Press, Inc. Orlando, Florida. [ Links ]
Dugdale, J. S. 1974. Female genital configuration in the classification of Lepidoptera. New Zealand Journal of Zoology, 1: 127-146. [ Links ]
Etman, A. M. & Hooper, G. H. S. 1979. Developmental and reproductive biology of Spodoptera litura (F.) (Lepidoptera: Noctuidae). Journal of the Australian Entomological Society, 18: 363-372. [ Links ]
Gurr, E. 1956. A Practical Manual ofMedical and Biological Staining Techniques. 2nd. edition. Interscience Publishers. New York. [ Links ]
Ivinskis, P., Rimsaite, J., Saldaitis, A. & Yakovlev, R. 2012. Description of two new species of Cossidae (Lepidoptera) from China. Zookeys, 192: 35-49. [ Links ]
Koshio, C., Tanaka, Y. & Kudo, S. 2002. The collapsed eggs found in the bursa copulatrix of a plum moth, Illiberis rotundata Jordan (Zygaenidae: Procridinae): an unusual egg resorption system? Journal of the Lepidopterists' Society, 56: 62-65. [ Links ]
Kristensen, N. P. 2003. Reproductive organs, pp. 427- 447. In: Kristensen, N. P. (Ed.). Handbook of Zoology. Arthropoda: Insecta. Lepidoptera, Moths and Butterflies.Vol. 2. Morphology, Physiology and Development. Walter de Gruyter GmbH & Co. KG, Berlin. [ Links ]
Llanderal-Cázares, C., Nieto-Hernández, R., Almanza-Valenzuela, I. & Ortega-Álvarez, C. 2007. Biología y comportamiento de Comadia redtenbacheri (Hamm) (Lepidoptera: Cossidae). Entomología Mexicana, 6: 252-255. [ Links ]
Martínez, M. I. 2002. Técnicas básicas de anatomía microscópica y de morfometría para estudiar los insectos. Boletín de la Sociedad Entomológica Aragonesa, 30: 187-195. [ Links ]
Miranda-Perkins, K. & Llanderal-Cázares, C. 2013. Cruzas con diferente proporción de sexos en Comadia redtenbacheri Hamm. Entomología Mexicana, 12: 530-533. [ Links ]
Miranda-Perkins, K., Llanderal-Cázares, C., De Los Santos-Posadas, H. M., Portillo-Martínez, L. & Vigueras-Guzmán, A. L. 2013. Comadia redtenbacheri (Lepidoptera: Cossidae) pupal development in the laboratory. Florida Entomologist, 96: 1424-1433. [ Links ]
Miskimen, G. W., Rodríguez, N. L. & Nazario, M. 1983. Reproductive morphology and sperm transport facilitation and regulation in the female sugarcane borer, Diatraea saccharalis (F.) Lepidoptera: Crambidae). Annals of the Entomological Society of America, 76: 248-252. [ Links ]
Pélisson, P. F., Bel-Venner, M. C., Rey, B., Burgevin, L., Martineau, F., Fourel, F., Lecuyer, C., Menu, F. & Venner, S. 2012. Contrasted breeding strategies in four sympatric sibling insect species: when a proovigenic and capital breeder copes with a stochastic environment. Functional Ecology, 26: 198-206. [ Links ]
Riddick, E. W. 2005. Are labcultured Anaphes iole females strictly proovigenic? Biocontrol, 50: 911-919. [ Links ]
Solomon, J. D. 1976. Sex ratio of the carpenterworm moth (Prionoxystus robiniae) (Lepidoptera: Cossidae). The Canadian Entomology, 108: 317-318. [ Links ]
Solomon, J. D. & Neel, W. W. 1973. Mating behavior in the carpenterworm moth, Prionoxystus robiniae (Lepidoptera: Cossidae). Annals of the Entomological Society of America, 66: 312-314. [ Links ]
Yakovlev, R. V. 2006. New Cossidae (Lepidoptera) from Asia, Africa and Macronesia. Tinea, 19: 188-213. [ Links ]
Yakovlev, R. V. 2008. Zeuzerocossus Yakovlev, gen. n., a new genus of Cossidae from south-eastern Asia. Tinea, 20: 105-107. [ Links ]
Yen, A. L. 2009. Entomophagy and insect conservation: some thoughts for digestion. Journal of Insect Conservation, 13: 667-670. [ Links ]
Yen, A. L. 2012. Edible insects and management of country. Ecological Management & Restoration, 13: 97-99. [ Links ]

