










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkActa zoológica mexicana
On-line version ISSN 2448-8445Print version ISSN 0065-1737
Acta Zool. Mex vol.30 n.1 Xalapa Apr. 2014
Ensayo
Validación de la familia Melolonthidae Leach, 1819 (Coleoptera: Scarabaeoidea)
Validity of the family Melolonthidae Leach, 1819 (Coleoptera: Scarabaeoidea)
Mariana Alejandra Cherman1 & Miguel Ángel Morón2*
1 Departamento de Zoologia, Universidade Federal do Paraná (UFPR), 81531-980, Curitiba, PR, Brasil.
2 Instituto de Ecología, A. C. Apdo. Postal 63, Xalapa, Veracruz 91000. México. *autor corresponsal: <miguel.moron@inecol.mx>
Recibido: 17/06/2013;
Aceptado: 21/11/2013.
RESUMEN
La familia Melolonthidae Leach, 1819 fue definida por Endrödi en 1966 como un grupo cosmopolita compuesto principalmente por las subfamilias Melolonthinae, Dynastinae, Rutelinae y Cetoniinae, formadas por taxones con hábitos fitófagos o fitosaprófagos, a diferencia de otros grupos de Scarabaeoidea. A pesar de ser ampliamente utilizada en América Latina desde 1976, esta clasificación es cuestionada por autores de estudios morfológicos y filogenéticos que apoyan la clasificación utilizada en América del Norte, la cual agrupa las subfamilias citadas junto con los Scarabaeinae y Aphodiinae coprófagos dentro de la familia Scarabaeidae Latreille, 1802. La presente revisión tiene como objetivo apoyar la validez de la familia Melolonthidae con modificaciones. Para ello se compararon diferentes clasificaciones utilizadas a lo largo de la historia de la coleopterología, así como los análisis para relacionar los diferentes grupos de Scarabaeoidea que se han publicado recientemente. Con dicha información y a reserva de efectuar nuevos estudios de morfología comparada y de obtener mejores datos con técnicas moleculares para apoyar un análisis filogenético más preciso, en el presente trabajo se propone aceptar como válido el nombre del agrupamiento denominado Melolonthidae constituído por el clado Dynastinae-Rutelinae-Melolonthinae. También se sugiere retirar a los Cetoniinae de aquel grupo y elevarlos a la categoría de familia, así como comprobar las relaciones de la subfamilia Orphninae con respecto a los miembros de la familia Melolonthidae. Finalmente, se indica que para lograr un consenso global sobre la clasificación supragenérica de los Melolonthidae es necesario efectuar análisis filogenéticos con taxones de todas las regiones del mundo, que efectivamente representen la riqueza específica de cada subfamilia y tribu, utilizando caracteres de larvas y adultos para confirmar su monofilia y las relaciones con otros grupos de Scarabaeoidea.
Palabras clave: Clasificación supragenérica, Cetoniinae, Melolonthinae, Rutelinae, Dynastinae, Orphninae, Scarabaeidae.
ABSTRACT
Melolonthidae Leach, 1819 was defined by Endrödi in 1966 as a cosmopolitan group mainly composed by the subfamilies Melolonthinae, Dynastinae, Rutelinae and Cetoniinae. They are formed by phytophagous or phytosaprophagous taxa, unlike other groups of Scarabaeoidea. Despite being commonly used in Latin America since 1976, it is questioned by many authors of morphological and phylogenetic studies that support the North American classification, formed by the above cited subfamilies and the coprophagous Scarabaeinae and Aphodiinae within Scarabaeidae Latreille, 1802. This review aimed to assess the validity of Melolonthidae with modifications. Thus, different classifications used throughout history were gathered as well as analyzes of relationships between different groups of Scarabaeoidea. Taking into account the latest information and with reserve for future studies on comparative morphology and the support of new accurate molecular data, the present work intends to validate the name of the taxon so called Melolonthidae, formed by the clade Dynastinae-Rutelinae-Melolonthinae. It is also suggested to remove Cetoniinae from Melolonthidae and raise it to family category and, by other hand, to test the relationships of Orphninae with the other members of Melolonthidae. Because of the high abundant and diversity taxa, in order to achieve a worldwide taxonomic consensus, phylogenetic analyzes of Melolonthidae with representative samples of all subfamilies and tribus from all over the world are needed using characters of larvae and adults, to confirm their monophyly, and the relationships with other groups of Scarabaeoidea.
Key words: Suprageneric classification, Cetoniinae, Melolonthinae, Rutelinae, Dynastinae, Orphninae, Scarabaeidae.
INTRODUCCIÓN
Los miembros de la familia Melolonthidae Leach, 1819 sensu Endrödi (1966) como parte de los Scarabaeoidea (Coleoptera) se caracterizan porque su maza antenal esta formada por tres a siete lamelas brillantes con sedas dispersas, cuerpo oval de colores diversos y los orificios respiratorios de los últimos tres segmentos abdominales están situados cerca del borde superior de los esternitos, entre otras características. El grupo surgió a partir de una combinación de propuestas de Bates (1887), Erichson (1848) y Janssens (1949). Endrödi (1966) analizó la clasificación de Scarabaeidae en series (Pleurosticti y Laparosticti) y propuso que los grupos que tuvieran las características de la serie Pleurosticti deberían ser elevados a una familia independiente denominada Melolonthidae. Así, la familia quedó compuesta por las subfamilias Melolonthinae, Dynastinae, Rutelinae, Cetoniinae, Glaphyrinae y Phaenomerinae, o sea, los taxones de Scarabaeoidea con hábitos fitófagos o fitosaprófagos.
Entre muchos otros autores citados por Kohlmann & Morón (2003) mencionaremos los trabajos de Erichson (1848) y Janssens (1949) quienes consideraron a los Scarabaeoidea formados por tres familias, Passalidae, Lucanidae y Scarabaeidae, la última dividida en dos series: Laparosticti y Pleurosticti. Actualmente, existen tres propuestas para agrupar los taxones dentro de Scarabaeoidea: 1) Principalmente en Eurasia se utiliza la propuesta de Balthasar (1963) (modificada por Paulian 1988) ha sido apoyada por los autores "splitters" (sensu Simpson 1945), en la cual los grupos de la serie "Pleurosticti" de Janssens (1949) fueron elevados a la categoría de familia, quedando formada por 18 familias, entre las cuales Melolonthidae (sensu Balthasar 1963) es equivalente a la subfamilia Melolonthinae de las otras clasificaciones. 2) La clasificación de Endrödi (1966) es utilizada ampliamente por especialistas de América Latina (Morón 1997) en especial los dedicados al control de escarabajos plaga. 3) En Norteamérica se desarrolló la clasificación de Lawrence & Newton (1995) (detallada por Ratcliffe et al. 2002), propuesta apoyada por autores "lumpers" (sensu Simpson 1945), que consideran válidas la mayoría de las familias de "Laparosticti" (sensu Balthasar 1963), pero las subfamilias de Melolonthidae (sensu Endrödi 1966) las incluyen en la familia Scarabaeidae como el conjunto de grupos de especies fitófagas (Pleurosticti) diferente a los grupos coprófagos (Scarabaeinae y Aphodiinae).
A nivel mundial ha predominado el uso de la clasificación de Lawrence & Newton (1995), debido a que se ha empleado como referencia en varios trabajos sobre morfología y filogenía que incluyen representantes de la mayor parte de las subfamilias de Scarabaeoidea, con los que ha sido posible evaluar las relaciones entre los grupos de la superfamilia (d'Hotman & Scholtz 1990, Scholtz 1990, Browne & Scholtz 1995, 1998, 1999). En forma paralela se ha mencionado que Melolonthidae es un nombre inválido de acuerdo con el Código Internacional de Nomenclatura Zoológica (1999) (Ratcliffe & Jameson 2004, Smith 2006). Sin embargo, recientemente la clasificación de Lawrence & Newton (1995) ha generado controversia porque en algunos estudios filogenéticos, como los de Smith et al. (2006), Hunt et al. (2007) y Lawrence et al. (2011), que incluyen caracteres morfológicos de larvas y adultos, así como caracteres moleculares de adultos de una amplia muestra de especies, se han obtenido resultados que no apoyan la monofilia de Scarabaeidae. En parte refutan las hipótesis de agrupamiento de los taxones obtenidas por Scholtz (1990) y Browne & Scholtz (1999), y no coinciden con los caracteres escogidos para probar la monofilia de algunos grupos (Smith et al. 2006, Micó et al. 2008). Los estudios recientes también apoyan el abandono definitivo de la división de los Scarabaeidae en las series Pleurosticti-Laparosticti, que durante más de un siglo ha sido empleada conscientemente por conveniencia (Arrow 1909, Ritcher 1969, Woodruff 1973) y en cambio adoptar ampliamente el agrupamiento de subfamilias propuesto por Endrödi (1966) revisado por Morón (2010).
Con la intención de apoyar la consolidación de un sistema de clasificación de los Scarabaeoidea mundialmente aceptado, el objetivo del presente trabajo consiste en evaluar la validez de la familia Melolonthidae sensu Endrödi. Tal revisión permitirá proveer argumentos para seguir utilizando esa clasificación, para modificarla, o para abandonarla y substituirla por otra más adecuada. Para ello es preciso reunir todas las evidencias existentes en cuanto a la historia de las clasificaciones y los análisis filogenéticos disponibles sobre los grupos de Scarabaeoidea, muchos de los cuales fueron mencionados por Kohlmann & Morón (2003).
Melolonthidae sensu Endrödi 1966: Diagnosis
Los miembros de la superfamilia Scarabaeoidea Latreille, 1802 agrupados dentro de la familia Melolonthidae Leach, 1819 poseen las siguientes características que los diferencian de otros grupos: presentan colores diversos y vestidura variada; el tamaño corporal varía entre 3 y 170 mm; mandíbulas bien desarrolladas, esclerosadas, con dientes en el borde interno, más o menos ocultas bajo el clípeo. Escapo antenal mucho más corto que el flagelo, maza antenal formada por tres a siete artejos alargados y delgados con forma de lamelas capaces de extenderse y plegarse como un abanico, con superficie brillante y setas dispersas; usualmente las lamelas son más largas en los machos que en las hembras. Cabeza pequeña, cuerpo ovalado y robusto, ocasionalmente deprimido o esbelto. Abdomen formado por seis segmentos visibles, los segmentos cuarto y quinto tienen los espiráculos situados en los extremos laterales de los esternitos, el penúltimo terguito fuertemente esclerosado y fusionado lateralmente con el esternito respectivo cerca del último par de espiráculos. Patas anteriores usualmente fuertes, a veces esbeltas y más alargadas en los machos. Los tarsos son pentámeros en los tres pares de patas, uñas tarsales bien desarrolladas, con formas diversas: simples, dentadas, bífidas, usualmente móviles y en pares, solo los Hopliinae tienen una uña metatarsal agrandada. Los élitros siempre están desarrollados, en ocasiones fusionados entre sí, y las alas metatorácicas usualmente bien desarrolladas, en pocos casos reducidas o ausentes. Exhiben con frecuencia dimorfismo sexual, los machos pueden presentar proyecciones en la cabeza y el pronoto con forma de cuernos o protuberancias con diseños muy diversos (Endrödi 1966, Morón 1997).
Las larvas son de tipo escarabaeiforme, presentan la galea y la lacínia maxilares completa o parcialmente fusionadas entre sí formando una mala. Palpos maxilares con cuatro artejos, mandíbulas con processo ventral, epifaringe sin epitorma, antenas con cuatro artejos, el último con forma diferente y provisto con áreas sensoriales de forma variable. Los tres pares de patas bien desarrolladas, cada pata con cuatro artejos diferenciados y el último tiene una uña (tarsungulus). Tienen un par de espiráculos protorácicos y ocho pares de espiráculos abdominales de tipo cribriforme. El abdomen consta de diez segmentos y la abertura anal usualmente es horizontal. Las pupas son exaradas, con abdomen formado por nueve segmentos bien delimitados, frecuentemente con órganos dioneiformes en sus bordes dorsales. Los espiráculos de los segmentos segundo a cuarto tienen un peritrema esclerosado, mientras que los correspondientes a los segmentos quinto a octavo están atrofiados, y carecen de peritrema (Morón 1993, 1997).
Los adultos se alimentan con tejidos, secreciones o restos vegetales y las larvas comunmente son fitosaprófagas, rizófagas o xilófagas (Endrödi 1966, Morón 1997). Este agrupamiento, caracterizado por los hábitos alimentarios fitófagos o fitosaprófagos ha sido aceptado históricamente y confirmado en varios estudios recientes (Scholtz & Chown 1995, Jameson & Ratcliffe 2002, Smith et al. 2006, Hunt et al. 2007, Lawrence et al. 2011).
La familia Melolonthidae está representada en todas las regiones biogeográficas y las subfamilias que la componen son taxones estables entre los diferentes sistemas de clasificación, que han cambiado de categoría según las opiniones de algunos autores (Endrödi 1966).
Creación de la familia Melolonthidae y resultados del trabajo de Endrödi (1966)
Una de las clasificaciones más antiguas de los Scarabaeoidea que incluye la diferencia en la posición de los espiráculos abdominales (Pleurosticti y Laparosticti), fue propuesta por Erichson (1848) y adoptada por especialistas connotados, como Lacordaire (1856), Paulian (1945), Janssens (1949), Crowson (1955), Britton (1957), Balthasar (1963), Medvedev (1976), Iablokoff-Khnzorian (1977) y Howden (1982).
Bates (1887) propuso la división del grupo Lamellicornia ( = Scarabaeoidea) en subtribus, con base en la estructura de la lígula y el mentón. La subtribu con la lígula y el mentón fusionados, es equivalente a la línea Pleurosticti de Erichson (1848) con los espiráculos abdominales ubicados en la parte lateral superior de los esternitos. Este grupo dio lugar a la serie Scarabaeidae-Pleurosticti de Janssens (1949) (Fig. 1).
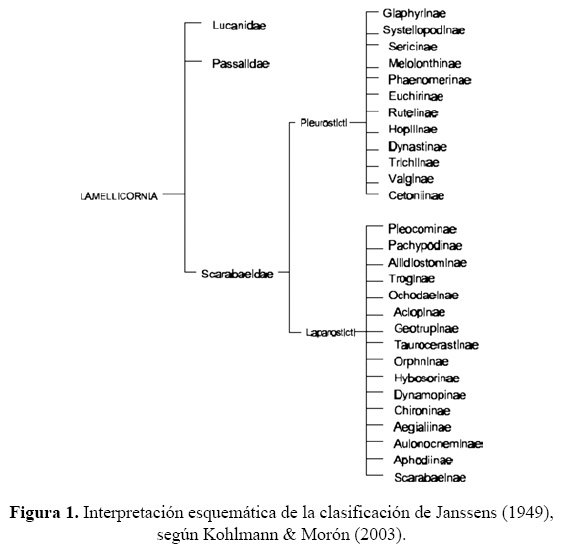
Endrödi (1966) reconoció este conjunto de taxones como una familia diferente, hermana de Scarabaeidae-Laparosticti, y eligió como género tipo a Melolontha Fabricius, 1775 que dio nombre a la familia Melolonthidae.
Actualmente, los artículos que hacen referencia a los Scarabaeoidea con hábitos fitófagos (Melolonthidae sensu Endrödi), utilizan el término tradicional "Scarabaeidae: Pleurosticti" (Jameson & Ratcliffe 2002). Sin embargo, esa clasificación se ha continuado utilizando por conveniencia (Woodruff 1973) a pesar de que se ha recomendado abandonarla (Arrow 1909). Ritcher (1969) realizó un trabajo de morfología comparada con los espiráculos abdominales para fundamentar las relaciones evolutivas entre los grupos de Scarabaeoidea y, aunque no propuso otra clasificación, demostró que tal agrupamiento no es válido (ver detalles más adelante).
Endrödi (1966) afirmó que las subfamilias que Janssens agrupó en la serie Pleurosticti tienen una relación filogenética mucho mayor entre sí que con las subfamilias agrupadas en la serie Laparosticti, a las que clasificó como Scarabaeidae sensu stricto (Cuadro 1). Con base en las propuestas de Bates (1887), Erichson (1848) y Janssens (1949), Endrödi propuso la familia Melolonthidae, compuesta por Melolonthinae Samouelle, 1819, Rutelinae MacLeay, 1819, Dynastinae MacLeay, 1819, Cetoniinae Leach, 1815, Euchirinae Hope, 1840, Systellopodinae Sharp, 1877, Glaphyrinae MacLeay, 1819 y Phaenomeridinae Erichson, 1847 (Cuadro 2). Actualmente Euchirinae se considera una subfamilia válida (Huchet 2003, Sipek et al. 2011) y los Systellopodinae se agrupan provisionalmente como parte de Melolonthinae. Los Glaphyrinae presentan una mezcla de características entre los Pleurosticti y los Laparosticti (Ritcher 1969), además de otros caracteres que han permitido separarlos como una familia independiente, más relacionada con los Ochodaeidae e Hybosoridae (Smith et al. 2006). Los Phaenomeridinae son un taxon africano cuyas larvas y biología son desconocidas (Scholtz & Grebennikov 2005); y en el trabajo de d'Hotman & Scholtz (1990) se menciona una posible relación de los Phaenomeridinae con los Melolonthinae y Rutelinae, tomando en cuenta el tipo único de su genitalia masculina.
La clasificación interna de los Melolonthidae inicialmente fue modificada y adaptada por Morón (1984) en forma similar a la propuesta de Erichson (1848) (Cuadro 1). Tales subfamilias tradicionalmente fueron incluidas por varios autores dentro de la familia Scarabaeidae Latreille, 1802 (Janssens 1949), y en la actualidad algunos autores insisten en mantenerlas como parte de los Scarabaeidae (Ratcliffe et al. 2002) (Cuadro 2).
Endrödi (1966) utilizó la taxonomía numérica como método para determinar las relaciones entre los Melolonthidae y otras familias de Scarabaeoidea (Cuadro 3), así como entre las propias subfamilias de Melolonthidae. Al analizar las relaciones entre estas familias el mismo autor propuso que el linaje Melolonthidae-Scarabaeidae se separó antes o después del Eoceno y dio origen a los dos grupos hermanos actuales. Con referencia en el registro fósil Scholtz & Chown (1995) hipotetizaron que el linaje de los Scarabaeoidea se originó en el Cenozoico, pero en los trabajos de Krell (2000) y Philips et al. (2004) se comprobó que el origen del grupo se encuentra en el Cretácico, de acuerdo con las evidencias paleontológicas y biogeográficas disponibles.

El sistema de clasificación de Endrödi (1966) fue adoptado por Morón (1976, 1984, 1997) y promovido entre los estudiantes, técnicos e investigadores de América Latina, sobre todo entre los interesados en el estudio de los escarabajos asociados con los cultivos agrícolas (Gassen 1993, Nájera-Rincón 1998, Oliveira & García 2003, Silva & Salvadori 2004, Boito et al. 2006, Pardo-Locarno et al. 2006, Neita-Moreno & Gaigl 2008). Siguiendo el criterio básico, en años recientes se han propuesto nuevos ajustes en las definiciones o diagnosis de las familias de Scarabaeoidea (Morón 2003, 2010, 2013).
Diferentes sistemas de clasificación y su influencia en la validez de Melolonthidae
La clasificación más utilizada para los Scarabaeoidea en el siglo XX incluía tres familias: Passalidae, Lucanidae y Scarabaeidae (Fig. 1) (Janssens 1949, Arnett 1963), pero se le ha modificado con diferentes criterios, entre los que destacan tres enfoques.
Balthasar (1963) originó la clasificación más utilizada en Europa, caracterizada por la jerarquización de familia para los grupos que otros autores consideran como subfamilias, y reúne 18 familias. En tal sentido, la subfamilia Melolonthinae (sensu Endrödi) corresponde a Melolonthidae sensu Balthasar (Cuadro 1). En parte de América Latina se emplea la clasificación de Endrödi (1966) adaptada por Morón (1984). Los especialistas de Norteamérica han adoptado la propuesta de Lawrence & Newton (1995) que fue apoyada por la escuela sudafricana (Browne & Scholtz 1995, 1999). Esta clasificación está compuesta por 12 familias (solo 6 menos que la de Balthasar), que resultaron del desmembramiento de las antiguas series Pleurosticti y Laparosticti (Jameson & Ratcliffe 2002). En dicho sistema las subfamilias del grupo de fitófagos (Melolonthidae sensu Endrödi) se encuentran dentro de la familia Scarabaeidae, junto con los Scarabaeinae y Aphodiinae, cuyos taxones son conocidos comúnmente como "escarabeidos coprófagos" (Cuadro 2).
Los defensores de la clasificación de Lawrence & Newton (1995) dudan de la monofilia de Scarabaeidae, pero debido a los escasos análisis filogenéticos de los grupos más derivados de Scarabaeoidea, y como no han encontrado suficientes caracteres sinapomórficos para establecer familias y subfamilias, dichos especialistas "lumpers" han optado por no dividir más el grupo (Jameson & Ratcliffe 2002). Los mismos autores que continúan aceptando la ubicación de los taxones fitófagos dentro de Scarabaeidae también argumentan sobre la invalidez del nombre Melolonthidae, dado que "Cetoniidae" tendría prioridad (Ratcliffe & Jameson 2004).
Para discutir sobre la validez de los nombres de grupos del nivel familia, en especial el nombre Melolonthidae, es necesario citar el Código Internacional de Nomenclatura Zoológica (ICZN) en su cuarta edición (1999). Inciso 35.5. Precedencia de nombres en uso en una categoría superior dice: "If after 1999 a name in use for a family-group taxon (e.g. for a subfamily) is found to be older than a name in prevailing usage for a taxon at higher rank in the same family-group taxon (e.g. for the family within which the older name is the name of a subfamily) the older name is not to displace the younger name". En el artículo 23.1, alusivo al Principio de Prioridad, se hace referencia a esa excepción en el inciso 23.1.1 (IZCN 1999). Ese inciso menciona que para el grupo-género el nombre más nuevo es sustituido por el más antiguo.
El nombre Melolonthidae fue utilizado por primera vez por Leach (1819). Según Endrödi (1966) Melolonthidae se compone de las subfamilias Melolonthinae Samouelle, 1819, Rutelinae MacLeay, 1819, Dynastinae MacLeay, 1819 y Cetoniinae Leach, 1815. Así, de acuerdo con el principio de prioridad, el nombre de la familia debería ser Cetoniidae. Sin embargo según el artículo 35.5 el uso de Melolonthidae sobre Cetoniidae se mantendría ya que el nombre Melolonthidae ha sido más utilizado, siempre que Melolonthinae y Cetoniinae sean aplicados como nombres para subfamilias dentro de Melolonthidae.
Ratcliffe & Jameson (2004) refutaron el nombre Melolonthidae tomando en cuenta solo el Principio de Prioridad (artículo 23.1) y sin considerar la excepción expuesta en el artículo 35.5 afirman que ese nombre es incorrecto y que, en caso de aceptarse la separación de los grupos fitófagos como un conjunto de nivel familia, ese debería denominarse Cetoniidae. Smith (2006: 146) citó el artículo 35.5 del código y precisamente utilizó Melolonthidae como ejemplo de la excepción del Principio de Prioridad. En esta forma, la excepción expuesta en el ICZN indica que el nombre Melolonthidae (sensu Endrödi 1966, Morón 1984, 1997) sería nomenclaturalmente válido. Sin embargo, para saber si la familia puede ser considerada válida, también deben ser revisados otros aspectos, como la presencia de sinapomorfías y las relaciones de éste taxón con otras familias.
Análisis filogenéticos y monofilia de Melolonthidae sensu Endrödi
Browne & Scholtz (1995, 1998, 1999) fueron los primeros en realizar estudios de la filogenia de Scarabaeoidea, que apoyaron la clasificación de Lawrence & Newton (1995). Para el análisis cladístico utilizaron 30 taxones de 13 familias de Scarabaeoidea (Cuadro 3) y 134 caracteres morfológicos de larvas y adultos, además de recodificar los 39 caracteres utilizados por Howden (1982). Según los cladogramas obtenidos en esos trabajos, Scarabaeidae sensu Lawrence & Newton (1995) es monofilético, sustentado por 23 estados derivados. Ese clado se mostró constituido por dos agrupamientos monofiléticos: Scarabaeinae-Aphodiinae y como grupo hermano el correspondiente a los taxones fitófagos (Melolonthinae, Rutelinae, Dynastinae, Cetoniinae). Este resultado apoyó durante más de una década la clasificación aceptada en América del Norte (Ratcliffe et al. 2002) y también es consistente con la ancestría común de esos taxones según Endrödi (1966).
Browne & Scholtz (1999) utilizaron pocos caracteres larvarios y obtuvieron índices de consistencia menores comparados con los caracteres de adultos. En un trabajo sobre filogenia de Cetoniidae, Micó et al. (2008) destacaron la importancia de los caracteres larvarios, que correspondieron a la mitad de los caracteres evaluados, con los que obtuvieron en su mayoría índices de consistencia mayores a 0.5. Grebennikov & Scholtz (2004) utilizaron sólo caracteres larvarios para evaluar la monofilia de Scarabaeoidea, y obtuvieron que Scarabaeidae sensu Lawrence & Newton (1995) no es monofilético.
La monofilia de los clados de taxones coprófagos (Scarabaeinae-Aphodiinae) y fitófagos (Melolonthinae-Rutelinae-Dynastinae-Cetoniinae) es confirmada en un análisis con caracteres moleculares de adultos (Smith et al. 2006) y en otro análisis con caracteres morfológicos de larvas y adultos (Lawrence et al. 2011). Sin embargo el estudio de filogenia molecular de Hunt et al. (2007) no apoya la monofilia del clado de los coprófagos. La aplicación estricta de los resultados obtenidos por Smith et al. (2006) con respecto a la monofilia de los Scarabaeidae sensu Lawrence & Newton (1995) cambiaría completamente esa clasificación (Fig. 2), ya que los Scarabaeinae y Aphodiinae se relacionan más estrechamente con los Glaphyridae, Hybosoridae y Ochodaeidae, que con los Dynastinae, Rutelinae, Cetoniinae y Melolonthinae, de lo cual podría determinarse que: 1) los Scarabaeinae y Aphodiinae forman parte de una familia (Scarabaeidae), 2) que sería necesario considerar a los Scarabaeidae y Aphodiidae como familias hermanas, o 3) que los Glaphyridae, Hybosoridae y Ochodaeidae son subfamilias de Scarabaeidae junto con Aphodiinae y Scarabaeinae. En forma paralela las subfamilias Cetoniinae, Dynastinae, Rutelinae y Melolonthinae formarían otra familia, hermana de Scarabaeidae.
Mediante el análisis molecular de 150 taxones de 13 familias de Scarabaeoidea Smith et al. (2006) obtuvieron poco soporte para la monofilia del clado coprófagos-fitófagos, como para continuar reconociéndolos como Scarabaeidae sensu Lawrence & Newton (1995). Un resultado parecido fue obtenido con el análisis bayesiano llevado a cabo por Hunt et al. (2007) y el cladograma más parsimonioso de Lawrence et al. (2011). Entre los comentarios incluidos en el artículo de Smith et al. (2006: 42) se propone reconocer al clado de los fitófagos como otra familia separada de Scarabaeidae (sensu lato), basándose en la filogenia de la superfamilia y en los hábitos fitófagos compartidos por los miembros de dicho clado. Esto es, dividir la familia definida por Scholtz (1990), Lawrence & Newton (1995) y Browne & Scholtz (1995, 1999) en dos familias: Scarabaeidae (constituída por Scarabaeinae y Aphodiinae) y Melolonthidae (incluyendo Melolonthinae, Rutelinae, Dynastinae y Cetoniinae) (Fig. 2).
La conclusión de Smith et al. (2006) es consistente con las propuestas de Endrödi (1966) y Morón (1984, 1997). Así, el agrupamiento constituido por los grupos fitófagos de Scarabaeoidea se confirma después de ser utilizado históricamente por más de 150 años como "Scarabaeidae: Pleurosticti". Melolonthidae debe substituir esa clasificación tradicional basada en la posición de los orificios respiratorios de los adultos, lo cual parece corresponder a caracteres sinapomórficos, ya que según Ritcher (1969) el tipo de orificios, su número y su ubicación en cada segmento abdominal pudieron ocurrir en diferentes linajes en distintos tiempos. En opinión de Morón (datos inéditos) el tema de los orificios respiratorios aún requiere ser estudiado comparativamente entre los distintos grupos de Scarabaeoidea, sobre todo en relación con los estados inmaduros, para entender mejor su evolución y el valor como caracteres en la sistemática del grupo.
Las relaciones internas en las subfamilias de Melolonthidae no han sido estudiadas empleando métodos filogenéticos modernos (Smith et al. 2006). Las subfamilias Rutelinae, Dynastinae y Melolonthinae parecen polifiléticas, y se dispone de escasos estudios sobre las relaciones en algunos de sus grupos (Sanmartín & Martin-Piera 2003; Ahrens 2006; Carrillo-Ruiz & Morón 2006; Smith et al. 2006; Ahrens et al. 2007; Coca-Abia 2007; Sípek & Ahrens 2011). Smith et al. (2006) confirmaron la monofilia de Cetoniinae (incluyendo Trichiini y Valgini), lo cual apoya los resultados de Browne & Scholtz (1998). Micó et al. (2008) evaluaron la monofilia de Cetoniidae sensu Krikken (1984), mediante un estudio filogenético biogeográficamente representativo, empleando caracteres morfológicos de larvas y adultos, y concluyeron que es un grupo monofilético que podría ser considerado en el nivel de familia. Este resultado refuerza la validez del nombre Melolonthidae para el clado compuesto por Dynastinae-Rutelinae-Melolonthinae.
Otro grupo tradicionalmente situado como subfamilia cuya posición amerita ser evaluada es Orphninae Erichson, 1847. En los trabajos de Browne & Scholtz (1998) y Hunt et al. (2007) dicho grupo fue relacionado con las subfamilias Dynastinae, Rutelinae, Melolonthinae y Cetoniinae (Fig. 3). Esa posición fue soportada por caracteres morfológicos en la región articular de las alas (Browne & Scholtz 1998), caracteres larvales (Paulian & Lumaret 1982; Morón 1991; Grebennikov & Scholtz 2004) y caracteres moleculares (Hunt et al. 2007). Scholtz & Chown (1995) hipotetizaron que Orphninae debería situarse en una posición basal con respecto al clado que incluye las subfamilias Melolonthinae, Dynastinae, Rutelinae y Cetoniinae (y por lo tanto podría ser miembro de Melolonthidae).
Algunas críticas a las clasificaciones de Scarabaeoidea
A pesar de la confirmación de la monofilia de Melolonthidae sensu Endrödi principalmente por el trabajo de Smith et al. (2006), sus relaciones con otros miembros de la superfamilia no se han precisado. En la clasificación de Endrödi (1966), Scarabaeoidea está formada por cinco familias: Lucanidae, Passalidae, Trogidae, Scarabaeidae (sensu stricto) y Melolonthidae. La familia Scarabaeidae sensu Endrödi, está compuesta por varios grupos que posteriormente fueron reubicados como familias en otras clasificaciones (Browne & Scholtz 1999; Carlson 2002a, 2002b; Ratcliffe et al. 2002), entre ellas se encuentran los Hybosoridae, Ochodaeidae y Glaphyridae (Cuadro 3).
En el cladograma de Smith et al. (2006) el clado de los fitófagos está dentro de un clado mayor monofilético constituído por dos subgrupos: Hybosoridae-Ochodaeidae-Glaphyridae (básicamente saprófagos y fitófagos) y Aphodiinae-Scarabaeinae (co-prófagos), cuya monofilia está soportada por la sinapomorfia de la genitalia bilobada (explicada en d'Hotman & Sholtz 1990). Para que la clasificación de Scarabaeidae sensu Lawrence & Newton (1995) tenga validez, una alternativa seria reubicar a los Hybosoridae, Ochodaeidae y Glaphyridae con categorías de subfamilias. Smith et al. (2006) opinaron que esa solución no sería la más correcta, ya que son taxones molecularmente muy divergentes, que además tienen características morfológicas, hábitos de alimentación e historias evolutivas diferentes, como mencionamos a continuación.
Hybosoridae Erichson, 1847 es considerada como familia por Lawrence & Britton (1994) y Jameson (2002) porque sus larvas poseen estructuras estriduladoras en las patas protorácicas y mesotorácicas y tres lóbulos truncados en el borde apical del labro, carácter único en Scarabaeoidea (Browne & Scholtz 1995). En Ochodaeidae Mulsant & Rey, 1871 la presencia del espolón pectinado en la mesotibia es un caracter único entre los grupos de la superfamilia y ayuda a establecer la monofilia de la familia (d'Hotman & Scholtz 1990, Scholtz 1990, Scholtz & Chown 1995, Carlson 2002a). La posición taxonómica de los Glaphyridae MacLeay 1819, ha sido tema de discusión durante mucho tiempo (Crowson 1955, Machatschke 1959, Ritcher 1969, Medvedev 1976, Iablokoff- Khnzorian 1977, Scholtz 1990, Carlson 2002b, Hawkins 2006, Smith et al. 2006), pero en el trabajo de filogenia molecular de Scarabaeoidea (Smith et al. 2006) se le confirmó como una familia independiente, concordando con los trabajos de genitalia masculina de Scarabaeoidea realizados por d'Hotman & Scholtz (1990). Atendiendo a la estructura de los orificios respiratorios larvarios Hinton (1967) también propuso que Glaphyrinae fuera elevado de categoria. Browne & Scholtz (1995) consideraron a Glaphyridae como un grupo monofilético, hermano de un clado grande (Trogidae, Bolboceratinae [Geotrupidae] y Pleocomidae) con base en los caracteres de la articulación alar. Esto último difiere de la posición de Glaphyridae en la filogenia de Smith et al. (2006), donde el grupo está relacionado con Ochodaeidae e Hybosoridae.
Hemos visto que de acuerdo con varios de los trabajos citados son evidentes algunas fallas en la clasificación de Scarabaeoidea sensu Endrödi (1966) y de Scarabaeidae sensu Lawrence & Newton (1995), pero también se han destacado evidencias que confirman la integración y validez de Melolonthidae como una familia natural.
CONSIDERACIONES FINALES
Con base en las evidencias expuestas se propone:
1. Reconocer con el nombre de Melolonthidae al clado Dynastinae-Rutelinae-Melolonthinae, delimitado por Endrödi (1966) y confirmado por Smith et al. (2006), considerando:
1.1. Aceptar Melolonthidae sensu Endrödi (1966) modificado por Morón (1997, 2010) como una familia válida, ya que los estudios morfológicos y moleculares preliminares recientes de la superfamilia Scarabaeoidea confirman su monofilia e independencia del clado de los escarabaeoídeos co-prófagos (Aphodiinae-Scarabaeinae). La siguiente diagnosis distingue a Melolonthidae: adultos con el labro y las mandíbulas parcialmente expuestas u ocultas por completo bajo el clípeo. Labro parcialmente esclerosado, membranoso o vestigial. Mandíbulas esclerosadas. Antenas formadas por 8-10 artejos, tres a siete de los cuales forman la maza antenal brillante, plegadiza. Con canthus ocular, sin escotadura anteocular acentuada. Quilla proepisternal ausente, incompleta o difusa. Proepímero plano o poco cóncavo. Mesepímero usualmente oculto bajo el élitro. Élitros sin emarginación posthumeral. Abdomen con seis esternitos visibles y siete pares de orificios respiratorios, tres o cuatro en las regiones pleurales, dos o tres en los extremos esternales y uno en la región tergal. Placa pigidial completamente expuesta o cubierta en parte por los élitros. Genitales masculinos bilobulados o fusionados. Larvas con antenas formadas por cuatro artejos alargados. Epifaringe asimétrica con tormae ampliamente separadas, y rara vez con clithra. Maxila con galea y lacinia fusionadas, unci de lacinia normalmente grandes. Placas respiratorias cribiformes. Patas posteriores largas, tarsungulos usualmente alargados, afilados.
1.2. Validar el nombre Melolonthidae de acuerdo con la excepción del Principio de Prioridad expuesto en el artículo 35.5 del ICZN (1999) Precedencia de los nombres en uso en una categoría superior.
1.3. Abandonar el término tradicional "Scarabaeidae:Pleurosticti" para referirse a los grupos que componen la familia Melolonthidae, pues se basa en caracteres heterogéneos que requieren mayor estudio comparativo para aplicarse en niveles supragenéricos.
1.4. Retirar la subfamilia Cetoniinae de la familia Melolonthidae, debido a su reciente confirmación como clado monofilético hermano de ese agrupamiento, con caracteres propios que lo delimitan. La siguiente diagnosis distingue a Cetoniidae como familia: adultos con el labro y las mandíbulas ocultas bajo el clípeo. Labro membranoso. Mandíbulas generalmente membranosas. Antenas formadas por 10 artejos, tres de los cuales corresponden a la maza antenal brillante, plegadiza. Con canthus ocular y escotadura anteocular acentuada. Quilla proepisternal bien marcada. Proepímero excavado. Mesepímero usualmente expuesto dorsalmente. Élitros frecuentemente con emarginación posthumeral amplia y acentuada. Abdomen con seis esternitos visibles y siete pares de orificios respiratorios, tres en las regiones pleurales, tres en los extremos esternales y uno en la región tergal. Placa pigidial usualmente no cubierta por los élitros. Genitales masculinos bilobulados. Larvas con antenas formadas por cuatro artejos cortos y anchos. Epifaringe simétrica con tormae muy separadas y frecuentemente con clithra. Maxila con galea y lacinia fusionadas, unci de lacinia reducidos. Placas respiratorias cribiformes. Patas posteriores cortas. Tarsúngulos usualmente reducidos. 1.5. Evaluar la posición de la subfamilia Orphninae, para confirmar sus relaciones con las subfamilias de Melolonthidae y ubicarla en una posición adecuada dentro del esquema de clasificación.
2. Abandonar los esquemas de clasificación regionales y desarrollar un sistema universal, basado en las relaciones evolutivas entre sus miembros. Esto es necesario teniendo en cuenta que:
2.1. Para proponer una nueva clasificación para el agrupamiento de taxones fitófagos de Scarabaeoidea se requiere desarrollar un estudio filogenético extensivo del clado Melolonthidae con un número representativo de los géneros del mundo, para confirmar la monofilia de cada subfamilia y desarrollar la clasificación natural de todos sus integrantes. Esto permitiría la discusión de conclusiones taxonómicas fundamentadas para que dicha clasificación pueda ser ampliamente aceptada.
2.2. Una vez que se ha validado la familia Melolonthidae es preciso revisar la clasificación de la superfamilia Scarabaeoidea, especialmente la monofilia de la familia Scarabaeidae (sensu stricto), puesto que existen evidencias de que no es un grupo natural.
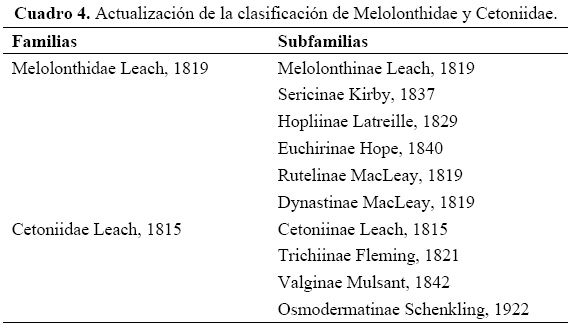
El esquema de clasificación resultado de las actualizaciones propuestas se muestra en el Cuadro 4, y se espera fundamentarlo y afinarlo con los análisis filogenéticos que estamos llevando a cabo.
AGRADECIMIENTOS
Se reconocen los comentarios de Estefanía Micó (Universidad de Alicante, España) y Fernando Vaz de Mello (Universidade Federal de Mato Grosso, Brasil) para aclarar algunas ideas sobre el tema de este ensayo, que constituye una contribución a la línea de investigación "Coleópteros Lamelicornios de América Latina", apoyada por el Instituto de Ecología, A. C. (cuenta 2003010011)
LITERATURA CITADA
Ahrens, D. 2006. The phylogeny of Sericini and their position within the Scarabaeidae based on morphological characters (Coleoptera: Scarabaeidae). Systematic Entomology, 31: 113-144. [ Links ]
Ahrens, D., Monaghan, M. & Vogler, A. 2007. DNA-based taxonomy for associating adults and larvae in multi-species assemblages of chafers (Coleoptera: Scarabaeidae). Molecular Phylogenetics and Evolution, 44: 436-449. [ Links ]
Arnett, R. H Jr. 1963. The beetles of the United States (A manual for identification). Catholic University of America Press, Washington, D.C. 1112 p. [ Links ]
Arrow, G. J. 1909. XV. On the characters and relationships of the less-known groups of lamellicorn Coleoptera, with descriptions of new species of Hybosorinae, etc. Transactions of the Royal Entomological Society of London, 57: 479 - 507. [ Links ]
Balthasar, V. 1963. Monographie der Scarabaeidae und Aphodiidae der palaearktischen und orientalischen Region (Coleoptera: Lamellicornia). Tschechoslowak Akademie der Wissenschaften, Praga. 1 - 3, 287 pp. [ Links ]
Bates, H. W. 1887. Biologia Centrali Americana, Insecta, Coleoptera: Lamellicornia. 2: 25-160. [ Links ]
Boito, G. T., Ornaghi, J. A., Giuggia, J. A. & Giovanini, D. 2006. Primera cita de dos especies de insectos sobre el cultivo de maní (Arachis hypogaea L.) en Córdoba, Argentina. Agriscientia, 23: 99-103. [ Links ]
Britton, E. B. 1957. A revision of the Australian Chafers (Coleoptera: Scarabaeidae: Melolonthinae). British Museum (Natural History) Journal, 1: 1- 185. [ Links ]
Browne, D. J. & Scholtz, C. H. 1995. Phylogeny of the families of Scarabaeoidea (Coleoptera) based on the characters of the hindwing articulation, hindwing base and wing venation. Systematic Entomology, 20: 145-173. [ Links ]
Browne, J. & Scholtz, C. H. 1998. Evolution of the scarab hindwing articulation and wing base: a contribution toward the phylogenys of the Scarabaeidae (Scarabaeoidea: Coleoptera). Systematic Entomology, 23: 307-326. [ Links ]
Browne, J. & Scholtz, C. H. 1999. A phylogeny of the families of Scarabaeoidea (Coleoptera). Systematic Entomology, 24: 51-84. [ Links ]
Carlson, D. C. 2002a. Ochodaeidae Mulsant and Rey 1870, pp. 28-31 In: Arnett, R. H. Jr., M. C. Thomas, P. S. Skelley & J. H. Frank (Eds.). American Beetles Volume 2. Polyphaga: Scarabaeoidea through Curculionoidea. CRC Press, Boca Raton, 2: 881 p. [ Links ]
Carlson, D. C. 2002b. Glaphyridae MacLeay 1819, pp. 37-38 In: Arnett, R. H. Jr., M. C. Thomas, P. S. Skelley & J. H. Frank (Eds.). American Beetles Volume 2. Polyphaga: Scarabaeoidea through Curculionoidea. CRC Press, Boca Raton, 2: 881 p. [ Links ]
Carrillo-Ruiz, H. & Morón M. A. 2006. Study on the phylogenetic relationships of the Hopliids (Coleoptera: Scarabaeoidea). Proceedings of the Entomological Society of Washington, 108: 619-638. [ Links ]
Coca-Abia, M. M. 2007. Phylogenetic relationships of the subfamily Melolonthinae (Coleoptera: Melolonthidae). Insect Systematics and Evolution, 38: 447-472. [ Links ]
Crowson, R. A. 1955. The natural classification of the families of Coleoptera. Nathaniel Lloyd & Co., Ltd., London, 187 p. [ Links ]
d'Hotman, D. & Scholtz, C. H. 1990. Phylogenetic significance of the structure of the external male genitalia in the Scarabaeoidea (Coleoptera). Entomological Memoir of the Department of Agricultural Development, 77: 1-51. [ Links ]
Endrödi, S. 1966. Monographie der Dynastinae (Coleoptera, Lamellicornia). Tiel 1. Abhandlungen Staatliches Museum für Tierkunde, Dresden, Bd. 33: 1-460. [ Links ]
Erichson, W. F.1848. Naturgeschichte der Insecten Deutschlands. Abt. I, Coleoptera 3: 801-968. [ Links ]
Gassen, D. N. 1993. Diloboderus abderus (Coleoptera: Melolonthidae) in notillage farming in Southern Brazil, pp. 129-141. In: Morón, M. A. (Ed.). Diversidad y manejo de plagas subterráneas. Sociedad Mexicana de Entomología e Instituto de Ecología, A. C. México. [ Links ]
Grebennikov, V. V. & Scholtz, C. H. 2004. The basal phylogeny of Scarabaeoidea (Insecta: Coleoptera) inferred from larval morphology. Invertebrate Systematics, 18: 321-348. [ Links ]
Hawkins, S. J. 2006. A revision of the Chilean tribe Lichniini Burmeister, 1844 (Coleoptera: Scarabaeidae: Melolonthinae). Zootaxa, 1266: 1-66. [ Links ]
Hinton, H. E. 1967. Structure and ecdysial process of the larval spiracles of the Scarabaeoidea, with special reference to those of Lepidoderma. Australian Journal of Zoology, 15: 947-953. [ Links ]
Howden, H. F. 1982. Larval and adult characters of Frickius Germain, its relationships to the Geotrupini, and a phylogeny of some major taxa in the Scarabaeoidea (Insecta: Coleoptera). Canadian Journal of Zoology, 60: 2713-2724. [ Links ]
Huchet, J. B. 2003. Faune de Madagascar. Insecta Coleoptera Chironidae. IRD (Eds.),CIRAD, MNHN, Paris, 91 p. [ Links ]
Hunt, T., Bergsten J., Levkanicova, Z., Papadopoulou, A., St. John, O., Wild, R., Hammond, P. M., Ahrens, D., Balke, M., Caterino, M. S., Gómez-Zurita, J., Ribera, I., Barraclough, T. G., Bocakova, M., Bocak, L. & Vogler, A. P. 2007. A Comprehensive Phylogeny of Beetles Reveals the Evolutionary Origins of a Superradiation. Science, 318: 1913-1916. [ Links ]
International Commission on Zoological Nomenclature (ICZN). 1999. International Code of Zoological Nomenclature, Fourth Edition. International Commission on Zoological Nomenclature, The Natural History Museum, London, U.K. 156 p. [ Links ]
Iablokoff-Khnzorian, S. M 1977. Uber die Phylogenie der Lamellicornia. Entomologische Abhandlungen der Staatlichen Museum für Tierkunde, Dresden 41: 135-199. [ Links ]
Jameson, M L. 2002. Hybosoridae Erichson 1847, pp. 32-33. In: Arnett, R. H. Jr., M. C. Thomas, P. S. Skelley & J. H. Frank (Eds.). American Beetles Volume 2. Polyphaga: Scarabaeoidea through Curculionoidea. CRC Press, Boca Raton, 2. 881 p. [ Links ]
Jameson, M. L. & Ratcliffe, B. C. 2002. Series Scarabaeiformia Crowson 1960 ( = Lamellicornia) Superfamily Scarabaeoidea Latreille 1802. Introduction, pp. 1-5. In: Arnett, R. H. Jr., M. C. Thomas, P. S. Skelley & J. H. Frank (Eds.). American Beetles Volume 2. Polyphaga: Scarabaeoidea through Curculionoidea. CRC Press, Boca Raton, 2: 881 p. [ Links ]
Janssens, A. 1949. Table synoptique et essai de classification pratique des Coléoptères Scarabaeidae. Institut royal des sciences naturelles de Belgique, 25: 1-30. [ Links ]
Kohlmann, B. & Morón, M. A. 2003. Análisis histórico de la clasificación de los Coleoptera Scarabaeoidea o Lamellicornia. Acta Zoológica Mexicana (n.s.) 90: 175-280. [ Links ]
Krell, F. T. 2000. The fossil record of Mesozoic and Tertiary Scarabaeoidea (Coleoptera: Polyphaga). Invertebrate Taxonomy, 14: 871-905. [ Links ]
Krikken, J. 1984. A new key to the suprageneric taxa in the beetle family Cetoniidae, with annotated lists of the known genera. Zoologische Verhandelingen, 210: 1-75. [ Links ]
Lacordaire, J. T. 1869. Histoire Naturelle des Insectes. Genera des Coléoptères. Paris, Roret. 9: 1-409. [ Links ]
Lawrence, J. F. & Britton, E. B. 1994. Series Scarabaeiformia, Superfamily Scarabaeoidea, pp. 98-104. In: Australian Beetles. Carlton, Melbourne University Press. 192 p. [ Links ]
Lawrence, J. F. & Newton, A. F. 1995. Families and subfamilies of Coleoptera (with selected genera, notes and references, and data on family-group names), pp. 779-1006. In: Pakaluk, J. & Slipinski, S. A. (Eds.), Biology, Phylogeny, and Classification of Coleoptera: Papers Celebrating the 80th Birthday of Roy A. Crowson, Warsaw: Muzeum i Instytut Zoologii PAN. 277 p. [ Links ]
Lawrence, J. F., Slipinski, A., Seago, A. E., Thayer, M. K., Newton, A. F. & Marvaldi, A. E. 2011. Phylogeny of the Coleoptera Based on Morphological Characters of Adults and Larvae. Annales Zoologici, 61: 1-217. [ Links ]
Leach, W. E. 1819. Melolonthidae, p: 189. In: Samouelle, G. (Ed.) The Entomologist's useful compendium. Thomas Boys, London. 496 p. [ Links ]
Machatschke, J. W. 1959. Untersuchungen über die verwandtschaftlichen Beziehungen der Gattungen der bisherigen Glaphyrinae (Coleoptera: Lamellicornia). Beiträge zur Entomologie, 9: 528-545. [ Links ]
Medvedev, S. I. 1976. Sistematike i Filogenii Plastinchatouyx Zhukov Palearktiki. Entomologischeskoe Obozreine, 55: 400-409. [ Links ]
Micó, E., Morón, M. A., Sipek, P. & Galante, E. 2008. Larval morphology enhances phylogenetic reconstruction in Cetoniidae (Coleoptera: Scarabaeoidea) and allows the interpretation of the evolution of larval feeding habits. Systematic Entomology, 33: 128-144. [ Links ]
Morón, M. A. 1976. Descripción de las larvas de tres especies mexicanas de pelidnotinos (Coleoptera: Melolonthidae: Rutelinae) y algunas observaciones sobre su biología. Anales del Instituto de Biología, Universidad Nacional Autónoma de México. Serie zoología, 74: 7-18. [ Links ]
Morón, M. A. 1984. Escarabajos, 200 millones de años de evolución. Publicación 14. Instituto de Ecología, México. 131 p. [ Links ]
Morón, M. A. 1991. Larva and pupa of Aegidium cribratum Bates (Coleoptera: Scarabaeidae: Orphninae). Coleopterists Bulletin, 45: 360-367. [ Links ]
Morón, M A. 1993. Observaciones comparativas sobre la morfología pupal de los Coleoptera Melolonthidae neotropicales. Giornale Italiano di Entomologia, 6: 249-255. [ Links ]
Morón, M. A. 1997. Antecedentes, pp.1-8. In: Morón, M. A., B. C. Ratcliffe, & C. Deloya (Eds.). Atlas de los escarabajos de México. Coleoptera Lamellicornia. Vol. I. Familia Melolonthidae. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad y Sociedad Mexicana de Entomología, México. 280 p. [ Links ]
Morón, M. A. 2003. Atlas de los escarabajos de México. Coleoptera: Lamellicornia. Vol. II Familias Scarabaeidae, Trogidae, Passalidae y Lucanidae. Argania editio. Barcelona. 227 p. [ Links ]
Morón, M. A. 2010. Diversidad y distribución del complejo "gallina ciega" (Coleoptera: Scarabaeoidea), pp. 41-63. In: Rodríguez del Bosque, L. A. & M. A. Morón (Eds.). Plagas del suelo. Mundi-Prensa, México. 417 p. [ Links ]
Morón, M. A. 2013. Introducción al conocimiento de los escarabajos de Puebla, pp. 1-27. In: Morón, M. A., A. Aragón & H. Carrillo (Eds.). Fauna de escarabajos del estado de Puebla. Publicación de Escarabajos Mesoamericanos, A. C., Coatepec, Veracruz, México. 467 p. [ Links ]
Nájera-Rincón, M. B. 1998. Diversidad y abundancia del complejo "gallina ciega" (Coleoptera: Melolonthidae) en agroecosistemas de maíz de la región templada de Michoacán, México, pp. 99-106. In: Morón, M. A. & A. Aragón (Eds.). Avances en el estudio de la diversidad, importancia y manejo de los coleópteros edafícolas americanos. Benemérita Universidad Autónoma de Puebla y Sociedad Mexicana de Entomología. México. 184 p. [ Links ]
Neita, J. C. & Gaigl, A. 2008. Escarabajos de importancia agrícola en Colombia (Coleoptera: Scarabaeidae "Pleurosticti"). Universidad Nacional de Colombia, Facultad de Agronomía. Bogotá D. C., 160 p. [ Links ]
Oliveira, L. J. & Garcia, M. A. 2003. Flight, feeding and reproductive behavior of Phyllophaga cuyabana (Moser) (Coleoptera: Melolonthidae) adults. Pesquisa agropecuária brasileira, 38: 179-186. [ Links ]
Pardo-Locarno, L. C., Morón, M. A. & Gaigl, A. 2006. Los estados inmaduros de Coelosis biloba (Coleoptera: Melolonthidae: Dynastinae) y notas sobre su biología. Revista Mexicana de Biodiversidad, 77: 215-224. [ Links ]
Paulian, R. 1945. Coléoptères Scarabéides de l'Indochine (Première Partie). Faune de l'Empire Français III. Librairie Larose, Paris. 225 p. [ Links ]
Paulian, R. 1988. Biologie des Coléoptères. Editions Lechevalier, Paris. pp. 76-80. [ Links ]
Paulian, R. & Lumaret, J.-P. 1982. La larve des Orphnidae. Bulletin de la Société Entomologique de France, 87: 263-272. [ Links ]
Philips, T. K., Pretorius, E., & Scholtz, C. H. 2004. A phylogenetic analysis of dung beetles (Scarabaeinae: Scarabaeidae): unrolling an evolutionary history. Invertebrate Systematics, 18: 53-88. [ Links ]
Ratcliffe, B. C. & Jameson, M. L. 2004. The Revised Classification for Scarabaeoidea: What the Hell is Going On? Scarabs, 15: 3-10. [ Links ]
Ratcliffe, B. C., Jameson, M. L. & Smith, A. B. T. 2002. Scarabaeidae Latreille 1802, pp. 39-81. In: Arnett, R. H. Jr., M. C. Thomas, P. S. Skelley & J. H. Frank (Eds.). American Beetles Volume 2. Polyphaga: Scarabaeoidea through Curculionoidea. CRC Press, Boca Raton, 881 p. [ Links ]
Ritcher, P. O. 1969. Spiracles of adult Scarabaeoidea (Coleoptera) and their phylogenetic significance, I. The abdominal spiracles. Annals of the Entomological Society of America, 62: 869-880. [ Links ]
Sanmartin, I. & Martin-Piera, F. 2003. First phylogenetic analysis of the subfamily Pachydeminae (Coleoptera, Scarabaeoidea, Melolonthidae): the Palearctic Pachydeminae. Journal of Zoological Systematics and Evolutionary Research, 41: 2-46. [ Links ]
Scholtz, C. H 1990. Phylogenetic trends in the Scarabaeoidea (Coleoptera). Journal of Natural History, 24: 1027-1066. [ Links ]
Scholtz, C. H. & Chown, S. L. 1995. The evolution of habitat use and diet in the Scarabaeoidea: a phylogenetic approach, pp. 355-374. In: J. Pakaluk & S. A. Slipinski (Eds.). Biology, Phylogeny, and Classification of Coleoptera: Papers Celebrating the 80th Birthday of Roy A.Crowson. Museum I Institut Zoologii PAN, Warszawa. [ Links ]
Scholtz, C. H & Grebennikov, V. V. 2005. Scarabaeoidea Latreille, 1802, pp. 368-425. In: R. G. Beutel & R. A. B. Leschen (Eds.) Handbook of Zoology, Coleoptera, Beetles. Volume 1. Morphology and Systematics (Archostemata, Adephaga, Myxophaga, Polyphaga partim. Berlin, New York: Walter de Gruyter. 567 p. [ Links ]
Silva, M. T. B. & Salvadori, J. R. 2004. Coródas-pastagens, pp. 191-210. In: Salvadori, J. R., Ávila C. J., Silva, M. T. B. (Eds.), Pragas de solo no Brasil. Passo Fundo: Embrapa Trigo. Dourados: Embrapa Agropecuária Oeste. Cruz Alta: Fundacep Fecotrigo. 544 p. [ Links ]
Simpson, G. G. 1945. The principles of classification and a classification of mammals. Bulletin of the American Museum of Natural History, 85: 1-350. [ Links ]
Sipek, P. & Ahrens, D. 2011. Inferring larval taxonomy and morphology in Maladera species (Coleoptera: Scarabaeidae: Sericini) using DNA taxonomy tools. Systematic Entomology, 36: 628-643. [ Links ]
Sipek, P., Jansta, P. & Král, D. 2011. Immature stages of Euchirinae (Coleoptera : Scarabaeoidea): genera Cheirotonus and Propomacrus with comments on their phylogeny based on larval and adult characters. Invertebrate Systematics, 25: 282-302. [ Links ]
Smith, A. B. T. 2006. A Review of the Family-group Names for the Superfamily Scarabaeoidea (Coleoptera) with Corrections to Nomenclature and a Current Classification. Coleopterists Society Monograph, 5: 144-204. [ Links ]
Smith, A. B. T., Hawks, D. C. & Heraty, J. M. 2006. An overview of the classification and evolution of the major scarab beetle clades (Coleoptera: Scarabaeoidea) based on preliminary molecular analyses. Coleopterists Society Monograph, 5: 35-46. [ Links ]
Woodruff, R. E. 1973. The scarab beetles of Florida (Coleoptera: Scarabaeidae) Part I. Arthropods of Florida and neighboring land areas, vol. 8. Florida Department of Agriculture and Consumer Services, Gainesville. 220 p. [ Links ]

