










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkActa zoológica mexicana
versão On-line ISSN 2448-8445versão impressa ISSN 0065-1737
Acta Zool. Mex vol.29 no.3 Xalapa Dez. 2013
Artículos originales
Suppression of Drosophila ananassae flies owing to interspecific competition with D. melanogaster under artificial conditions
Supresión de las moscas Drosophila ananassae debido a la competencia interespecífica con D. melanogaster bajo condiciones artificiales
Arvind K. SINGH1 & Sanjay KUMAR
1 Genetics Laboratory, Department of Zoology, Banaras Hindu University, Varanasi-221005 INDIA. <aksbhu23@rediffmail.com>
Recibido: 18/09/2012;
aceptado: 22/05/2013.
ABSTRACT
Interspecific competition between two species of Drosophila: D. ananassae and D. melanogaster was studied at the larval and adult stages. It was found that when D. ananassae and D. melanogaster adult flies were co-cultured, very few D. ananassae offspring could be recovered in the first generation. To investigate the reasons of D. ananassae apparent inhibition, mating behavior of D. ananassae in the presence of D. melanogaster was observed and it was found that the number of matings deviated significantly from those recorded when it was kept alone. To determine larval development of D. ananassae after being initially exposed to D. melanogaster, the females of the two species were separated in different food bottles after 3 days of being kept together. Good D. ananassae cultures could be recovered indicating that initial exposure of D. ananassae to D. melanogaster did not hamper its egg laying capacity or eclosion. However, if they remained together, no D. ananassae could be recovered from larval diet, suggesting that either D. melanogaster adults interfered with fertilization or egg-laying, or their larvae eliminated competitors. To see whether there is larval competition, polytene chromosomes of 54 third instar larvae were analyzed out of which only 5.56 percent were found to be D. ananassae. Thus, if a few eggs are laid by D. ananassae and they develop, all the while facing competition from D. melanogaster and till the third instar larval stage is reached, there is almost complete elimination of D. ananassae. Thus, interspecific competition exists at all stages of life cycle and few if any D. ananassae flies emerge.
Key words: Interspecific competition, mating propensity, co-culture, fecundity, larval competition, D. ananassae.
RESUMEN
Se estudió la competencia interespecífica entre dos especies de Drosophila: D. ananassae y D. melanogaster en estado larvario y adulto. Se encontró que al criar simultáneamente adultos de D. ananassae y D. melanogaster, muy poca progenie de primera generación de D. ananassae podia recuperarse. Para investigar las causas de la aparente inhibición de D. ananassae, se observó el comportamiento de apareamiento de D. ananassae en presencia de D. melanogaster y se observó que el número de apareamientos se desvió significatívamente de aquel registrado cuando esta especie se mantuvo sola. Para observar el desarrollo larvario de D. ananassae después de la exposición a D. melanogaster, hembras de ambas especies fueron separadas en botes de alimento después de tres días de confinamiento conjunto. Se obtuvieron buenas crías de D. ananassae, indicando que la exposición de D. ananassae a D. melanogaster no afectó su capacidad de oviposición ni su fertilidad. Sin embargo, cuando ambas especies permanecieron juntas, no se recuperó D. ananassae de la dieta larvaria lo que sugiere que los adultos de D.melanogaster interfirieron con la fertilización u oviposición, o bien sus larvas eliminaron la competencia. Para establecer la existencia de competencia larvaria se examinaron los cromosomas polténicos de 54 larvas de tercer estadio entre las cuáles solo el 5.56 por ciento resultaron ser de D. ananassae. Por lo tanto, si algunos huevos son depositados por D. ananassae y se desarrollan, éstos enfrentan la competencia de D. melanogaster durante todo el desarrollo y hasta el tercer estadio, llegando a la casi total eliminación de D. ananassae. Existe competencia interespecifica en todas las etapas del ciclo de vida resultando en una casi nula emergencia de D. ananassae.
Palabras clave: Competencia interespecífica, co-cultivo, fecundidad, competencia larval, D. ananassae.
INTRODUCTION
Natural selection often operates through competition, allowing various forms to survive and establish under certain environmental conditions. It tends to eliminate, gradually or rapidly, forms exploiting identical niches. Charles Darwin (1859) in his book "Origin of species" stated that Intraspecific and interspecific competition among organisms are a part of natural selection. This competition can be the result of high reproductive potential of some species when compared to the environmental carrying capacity. The most severe form of interspecific competition exists when organisms of two species have the same requirements. On the basis of theoretical equations, Lotka (1925) and Volterra (1926) independently predicted that two species with identical needs and habits cannot survive in the same place if they compete for limited resources. If both types are found together in nature, they must differ in their ecology, or else there are fluctuations in the environment, favoring first one and then the other. Experiments on interspecific competition between similar species have also demonstrated the validity of this theory (Connell 1981; Crombie 1947; Denno et al. 1995; Gause 1934; Hochkirch & Groening 2012; Luan et al. 2012; Zhang et al. 2011; Zimmering 1948). Careful analysis of the instances in which both species survive has shown that they occupy slightly different niches (Gause 1934; Crombie 1947). According to Kohn and Orians (1962) competition between species reveals ecological differences permitting coexistence of stable populations of closely related species of various kinds of animals.
Drosophila turns out to be the apt model system for studying interspecific competition, because in the genus Drosophila one finds a large number of species, some showing high degrees of morphological and ecological similarities and also close genetical relationships (Tantawy 1964). No wonder, exhaustive studies have been done, especially involving sibling species of this genus to study interspecific competition (Nunney 1990). A practical way of studying interspecific competition in the laboratory is by co- culturing two species together in large population cages or in culture bottles. One may simply study the effect of co-culturing on the fecundity of the species involved or one can also check the effect of altering various ecological conditions such as larval density, temperature etc. on the fecundity of co-cultured populations of two species. Merrell (1951) cultured D. melanogaster and D. funebris together in food bottles and found that the proportion of D. funebris eggs reaching pupation decreased significantly in mixed larval populations. Interspecific competition between D. melanogaster and D. simulans, was studied in much detail by Barker & Podger (1970). They examined the effects of larval density and short-term adult starvation on fecundity, egg hatchability and adult viability. It was found that significant effect of larval density on fecundity was probably mediated through effects on adult body weight. D. melanogaster females raised in mixed species cultures were less fecund than those from pure cultures, while D. simulans showed the reverse effect. Increasing the larval density of the two species at 15 °C and 25 °C temperatures, causes a progressive reduction in per cent hatching (Tantawy & Soli-man 1967; Miller 1964; Chiang & Hodson 1950; Birch 1955). Boggild & Keiding (1958) in their studies on house fly larvae found that harmful effects of crowding on adult emergence result from an increased incidence of collisions between larvae. This causes the metabolic rate to be increased and thus reduces the fraction of ingested food available for tissue formation. Tantawy & Soliman (1967) found that there was a gradual elimination of D. simulans by D. melanogaster in the two cages, irrespective of their initial percentage at 25 °C. These results also supported the results of Moore (1952) and Barker (1963). The results from cages kept at 15 °C indicate that the outcome of competition at 15 °C is completely different from that at 25 °C. At 15 °C there is an increase in D. simulans at the expense of D. melanogaster. A similar experiment was done by Montchamp-Moreau (1983) involving the same species. The optimum temperature for D. simulans was found to be 20 °C and for D. melanogaster it was 25 °C as while at 25 °C D. melanogaster eliminated D.simulans and at 20 °C the reverse occurred in mix culture cage populations. D. pseudoobscura and D. subobscura, two species sympatric in the west coast of North America exhibited competition in the laboratory to such an extent that D. subobscura was completely eliminated in just a few generations of their being co-cultured. The outcome was no different when they were reared at different temperatures (Pascual et al. 1998).
In nature two competing species do not face a straight battle and there may be other species too which have their own impact on the competition. For example it was found that competition between two closely related species D. melanogaster and D. hydei was affected by a third distant species of an entirely different kingdom Aspergilus niger ( Hodge et al. 1999). Not only other species but components of interspcific competition are also known to be affected under different environmental conditions, creating a geographic mosaic of outcomes (Joshi 2004; Joshi & Thompson 1995). Price et al. (2012) found that presence of mating rivals cause males of D. pseudoobscura to modulate sperm transfer through an increase in sperm transfer, alteration in ejaculate composition etc. In D. melanogaster Bretman et al. (2009) reported that males kept with rivals prior to mating, mated for a longer duration.
The present study aimed at comparing the viability of two species of Drosophila,i.e. D. ananassae Doleschall and D. melanogaster Meigen when they are cultured together in food bottles. Taxonomically both species belong to the melanogaster species group of the subgenus Sophophora. Drosophila ananassae falls in the ananassae subgroup and the ananassae complex and D. melanogaster belongs to the melanogaster subgroup (Bock & Wheeler 1972). Distribution wise both species are cosmopolitan. D. melanogaster being semi-domestic, occurring in orchards and gardens and D. ananassae, a domestic species is found at a closer proximity to human habitation, such as fruit markets, kitchens etc. It occupies a unique status among the Drosophila species because of certain unusual genetical features (Singh 2010). The morphological features of the two species enable easy distinction of the two species and their sexes as males of D. melanogaster possess black abdominal tip and a characteristic pattern of sex comb. The females of D. melanogaster are larger in size with more swollen abdomen and black thin stripes at abdominal segment junctions. Drosophila ananassae males do not possess black coloration at the abdominal tip and they have a number of thick hairs on the first and second tarsal segments forming a diffused sex comb pattern. The females of D. ananassae are light in color as they lack black abdominal stripes. Earlier experiments on interspecific competition involving D. melanogaster have been done with its sibling species. However, D. ananassae and D. melanogaster are quite distinct. Therefore, it would be interesting to see the degree of competition that exists between the two, and whether they are able to thrive in presence of each other. Experiments were conducted to determine at which stage(s) of the life cycle interspecific competition occurs between the two species.
MATERIAL AND METHODS
This study was done by taking two wild type species of Drosophila, D. ananassae and D. melanogaster. These two stocks have been reared in our laboratory for the last three years. D. ananassae was originally collected from Ranchi (Jharkhand state) and D. melanogaster was collected from Varanasi (Uttar Pradesh), as adult forms by net sweeping. The stocks of both species were maintained in the laboratory on simple yeast-agar culture medium at 24oC with a 12 hours cycle of light and darkness.
Virgin male and female flies from both species were collected and aged. Twenty pairs (ten pairs from each species) of seven day-old flies were then kept together in 250 ml cylindrical culture bottles (height 15 cm, base diameter 5 cm, mouth diameter 2.5 cm) containing 50 ml of food. After three days, all flies were discarded. Flies emerging from the bottles were separated according to species and sex and counted. Pure cultures were established to serve as controls. The number of progeny emerging from such control cultures were also counted. The experiment was repeated five times.
Mating behavior of D. ananassae and D. melanogaster flies was observed in the presence of each other. Ten pairs of seven day-old virgin male and female flies of both species were kept in an Elen-Wattiaux mating chamber and observed for one hour. When a pair commenced mating it was aspirated out. As a control, 10 pairs of D. ananassae and 10 pairs of D. melanogaster were observed separately in the mating chamber. The total number of matings (formed couples) and the time interval in minutes from release to pair formation was recorded. Six replicates of each mating combination were carried out. All mating experiments were done between 6:00 am to 11:00 am, as Drosophila exhibits peak mating activity during the morning hours. One way ANOVA was done to test whether there was any difference in the mean number of matings among the four groups, (D. ananassae alone, D. ananassae in presence of D. melanogaster, D. melanogaster alone and D. melanogaster in presence of D. ananassae). Bonferroni t- test were used for pair wise comparison between the groups.
Ten pairs of seven day-old virgin male and female flies of both the species were kept together in culture bottles (as described above). After three days males were discarded and females of each species were kept separately in fresh culture bottles, and allowed to lay eggs. They were not exposed to males of their own species so that the progeny produced was only the outcome of matings that occurred when both the species were housed together. Also the effect of initial exposure of the two species to each other, on their progeny recovery could be determined.
The two species were also co-cultured in the food bottles to study larval competition. Ten pairs of seven day-old virgin male and female flies of both species were kept in fresh food bottles for three days, after which the flies were discarded. Polytene chromosome preparations of the third instar larvae were made by lacto-aceto-orcein method, to identify whether the larvae were D. ananassae or D. melanogaster.
RESULTS
Drosophila ananassae flies were almost eliminated in number during the first generation itself, when reared in culture bottles along with D. melanogaster adults as very few D. ananassae, males could be recovered. The numbers of D. melanogaster (both males and females) counted in the bottles are given in Table 1. Also, the numbers of flies counted in the control sets are shown.

In the second set of experiments, where males and females of both the species were observed in Elen-Wattiaux mating chamber, it was found that males of both species largely courted females of their own species. However, occasionally, D. melanogaster males would unsuccessfully court a D. ananassae female. It was also observed that D. melanogaster males would interfere with the courtship rituals of D. ananassae males and chased away a courting D. ananassae male. The results of one way ANOVA (Table 2.) show that there is a significant difference between the mean number of matings in D. ananassae and D. melanogaster when they are confined alone in the mating chamber and when they are kept in presence of each other. Bonferonni t-tests (Table 3.) proved that there are significant reductions in the number of D. ananassae matings, when it is kept with D. melanogaster. Therefore the presence of D. melanogaster affects the mating propensity of D. ananassae. Figure 1 shows that as the number of D. melanogaster pairs decreases in the mating chamber after being aspirated out when a pair commenced mating, the number of matings of D. ananassae increases. The average data of the six sets of mating experiments with ten pairs of D. ananassae in the presence of 10 pairs of D. melanogaster was used. Figure 2 shows the effect of time on mating success in D. ananassae and D. melanogaster in different conditions. The graph was constructed with the pooled data of the six sets of mating experiments. It depicts that the number of matings in D. ananassae in first 10 minutes is very high, when they are alone in the mating chamber, compared to when they are confined with D. melanogaster. In the next 50 minutes, however, the number of matings remains almost the same under the two conditions. For D. melanogaster, the number of matings was found to be less when alone than when in the presence of D. ananassae.


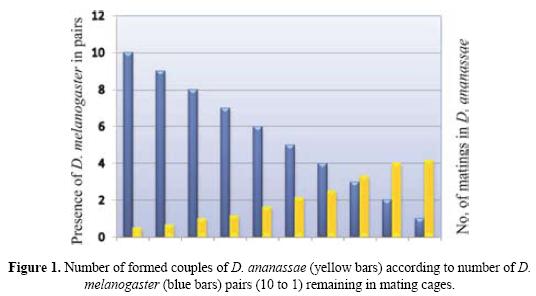
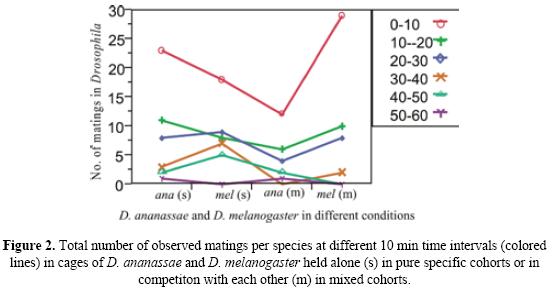
Initial exposure of the two species to each other does not affect fecundity or larval development, as good D. ananassae and D. melanogaster cultures could be recovered from culturing females which had been exposed to males of the other species earlier.
From the mixed culture of the two species, larvae were also analyzed to identify which species they belonged to by observing their salivary gland chromosomes.
Out of fifty four larvae examined only three were found to be D. ananassae and the remaining larvae were all identified as D. melanogaster. Thus, bulk mortality of D. ananassae occurs in or before the third instar larval stage, with only a handful remaining alive to reach the pupal stage and even less if any emerging as adults.
DISCUSSION
Our experiments demonstrate that D. ananassae flies cannot thrive when confined in the small space of a food bottle with D. melanogaster. A finding consistent with observation of their habits in their natural habitat (Bock & Wheeler 1972), where in the presence of one, the other prefers another niche. It has also been observed by the authors that in nature when there is abundance of D. melanogaster in a certain place, D. ananassae is found fewer in number.
To go deeper into understanding the nature of interspecific competition that exists between the two species, we planned different experiments and found that competition starts right at the level of courtship rituals and matings and lasts until the late larval stages and perhaps beyond until D. ananassae is eliminated completely or its development severely hindered. Though, it is not sure what the outcome would have been if the two species were co-cultured in larger enclosures, one thing is certain, if D. ananassae is almost eliminated in the first generation in food bottles, in larger enclosures too it would not take a number of generations for it to disappear.
Fitness of D. ananassae is reduced in presence of D. melanogaster as D. melanogaster males interfere with the courtship rituals of D. ananassae males and prevent fertilization. Therefore in the presence of D. melanogaster, D. ananassae mates more infrequently than normal.
D. ananassae females which have succeeded in mating, may face competition from D. melanogaster females for egg laying sites. Given that when they were exposed initially to D. melanogaster and separated from them latter, their egg laying was normal and resulted in good larval recovery. Therefore, fitness of D. ananassae is affected only as long as D. melanogaster is present.
It was not possible for us to utilize the first and second instar larval stages for species identification, as they are small, present deep inside the food and their polytene chromosome preparations are not very good. However, the fact that a few D. ananassae third instar larvae could be identified through polytene chromosomes, indicates that some D.ananassae larvae are able to fight and withhold the battle until the third larval stage, facing a tough competition from D. melanogaster.
We observed that out of few progeny of D. ananassae reaching the third instar larval stage owing to larval competition, fewer pupate and lesser hatch as flies. Similar result was found by Merrel (1951) on D. funebris and D. melanogaster. This may indicate that individuals surviving to stage display lower survival probabilities than those stemming from pure cultures.
A very interesting observation was that while in D. ananassae the number of mating is reduced in presence of D. melanogaster, with D. melanogaster just the opposite occurs, that is D. melanogaster mates more rapidly in the presence of D. ananassae than when held alone at similar densities.
Both these behaviors are the outcomes of competition, a proof that both the species compete with each other. In the face of competition while D. ananassae may be conserving the energy required in courtship and matings (an important cost of reproduction) and invest it in survival waiting for less restrictive conditions, D. melanogaster increases its mating rate, perhaps to serve the purpose of ousting the competitor with the weapon of numbers. Under our experimental conditions the strategy displayed by D. melanogaster is perhaps better, as clearly it wins the battle. Yet D. ananassae might be more successful under different environmental conditions.
ACKNOWLEDGEMENT
We are highly grateful to Ms. Parul Banerjee for her help in the preparation of this manuscript. We are thankful to University Grants Commission, New Delhi for giving financial assistance to S K in the form of Project fellow. We thank anonymous reviewers for their very appropriate corrections and suggestions in the manuscript.
LITERATURE CITED
Barker, J. S. F. & Podger, R. N. 1970. Interspecific competition between Drosophila melanogaster and Drosophila simulans. Effects of larval density and short-term adult starvation on fecundity, egg hatchability and adult viability. Ecology, 51: 855-864. [ Links ]
Barker, J. S. F. 1963. The estimation of relative fitness of Drosophila populations. II. Experimental evaluation of factors affecting fitness. Evolution, 17: 56-71. [ Links ]
Birch, L. C. 1955. Selection in Drosophila pseudoobscura in relation to crowding. Evolution, 9: 389-399. [ Links ]
Bock, I. R. & Wheeler, M. R. 1972. The Drosophila melanogaster species subgroup. University of Texas Publications, 7213: 1-102. [ Links ]
Boggild, O. & Keiding, J. 1958. Competition in house fly larvae. Experiments involving a DDT-resistance and susceptible strains. Oikos, 9: 1-25. [ Links ]
Bretman, A., Fricke, C. & Chapman, T. 2009. Plastic response of male Drosophila melanogaster to the level of sperm competition increase male reproductive fitness. Proceedings of the Royal Society of London, Biological Sciences, 276: 1705-1711. [ Links ]
Chiang, H. C. & Hodson, A. C. 1950. An analytical study of population growth in Drosophila melanogaster. Ecological Monographs, 20: 173-206. [ Links ]
Connell, A. D. 1981. The taxonomy and distribution of some calanoid Copepods in South African east coast estuaries. Annals Natal Museum, 24: 489-500. [ Links ]
Crombie, A. C. 1947. Interspecific competition. Journal of Animal Ecology, 16: 44-73. [ Links ]
Darwin, C. 1859. The origin of species. Random House, New York. [ Links ]
Denno, R. F., McClure, M. S. & Ott, J. R. 1995. Interspecific interactions in phytophagous insects: competition reexamined and resurrected. Annual Review of Entomology, 40: 297–331. [ Links ]
Gause, G. F. 1934. The struggle for existence. Williams and Wilkins, Baltimore. [ Links ]
Hochkirch, A. & Groening, J. 2012. Niche overlap in allotopic and syntopic populations of sexually interacting ground-hopper species. Insect Science, 19: 391-402. [ Links ]
Hodge, S., Mitchell, P. & Arthur, W. 1999. Factors affecting the occurrence of facilitative effects in interspecific interactions: an experiment using two species of Drosophila and Aspergillus niger. Oikos, 87: 166-174. [ Links ]
Joshi, A. & Thompson, J. N. 1995. Alternative routes to the evolution of competitive ability in two competing species of Drosophila. Evolution, 49: 616-625. [ Links ]
Joshi, A. 2004. Variation in the relative magnitude of intraspecific and interspecific competitive effects in novel versus familiar environments in two Drosophila species. Journal of Genetics, 83: 179–188. [ Links ]
Kohn, A. J. & Orians, G. H. 1962. Ecological data in the classification of closely related species. Systematic Zoology, 11: 119-127. [ Links ]
Lotka, A. J. 1925. Elements of physical biology. Williams and Wilkins, Baltimore. [ Links ]
Luan, J., Xu, J., Lin, K. K., Myron, P. Z. & Liu, S. S. 2012. Species Exclusion Between an Invasive and an Indigenous Whitefly on Host Plants with Differential Levels of Suitability. Journal of Integrative Agriculture, 11: 215-224. [ Links ]
Merrell, D. J. 1951. Interspecific Competition between Drosophila funebris and Drosophila melanogaster. The American Naturalist, 85: 159-169. [ Links ]
Miller, R. C. 1964. Larval competition in Drosophila melanogaster and D. simulans. Ecology, 45: 132-148. [ Links ]
Monnchamp-Moreau, C. 1983. Interspecific competition between Drosophila melanogaster and D. simulans: Temperature effect on competitive ability and fitness component. Genetics Selection Evolution, 15: 367-378. [ Links ]
Moore, J. A. 1952. Competition between Drosophila melanogaster and D. simulans. Evolution, 6: 407–420. [ Links ]
Nunney, L. 1990. Drosophila on oranges: colonization, competition, and coexistence. Ecology, 71: 1904–1915. [ Links ]
Pascual, M., Serra, L. & Ayala, F. J. 1998. Interspecific laboratory competition of the recently sympatric species Drosophila subobscura and Drosophila pseudoobscura. Evolution, 52: 269-274. [ Links ]
Price, T. A. R., Lize, A., Marcello, M. & Bretman, A. 2012. Experience of mating rivals causes males to modulate sperm transfer in the fly Drosophila pseudoobscura. Journal of Insect Physiology, 58: 1669-1675. [ Links ]
Singh, B. N. 2010. Drosophila ananassae: A good model species for genetical, behavioural and evolutionary studies. Indian Journal of Experimental Biology, 48: 333-345. [ Links ]
Tantawy, A. O. & Soliman, M. H. 1967. Studies on natural populations of Drosophila VI. competition between Drosophila melanogaster and Drosophila simulans. Evolution, 21: 34-40. [ Links ]
Tantawy, A. O. 1964. Studies on natural populations of Drosophila. III. Morphological and genetical differences in wing length of Drosophila melanogaster and D. simulans in relation to seasonal fluctuations. Evolution, 18: 560-570. [ Links ]
Volterra, V. 1926. Variazioni e fluttuazioni del numero d'individui in specie animali conviventi. Mem. R. Accad. Nap. dei Lincei. Ser. VI, vol. 2 (Trans. in appendix of Chapman, R. N., 1931, Animal ecology. McGraw Hill, New York. [ Links ]
Zhang, G. F., Li, D. C., Liu, T. X., Wan, F. H. & Wang, J. J. 2011. Interspecific Interactions Between Bemisia tabaci Biotype B and Trialeurodes vaporariorum (Hemiptera: Aleyrodidae). Environmental Entomology. 40: 140-150. [ Links ]
Zimmering, S. 1948. Competition between Drosophila pseudoobscura and Drosophila smelanogaster in population cages. The American Naturalist, 82: 326-330. [ Links ]

