










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkActa zoológica mexicana
versión On-line ISSN 2448-8445versión impresa ISSN 0065-1737
Acta Zool. Mex vol.26 no.3 Xalapa dic. 2010
Artículos originales
Cambios en las poblaciones de aves rapaces migratorias en Veracruz, México, 1995–2005
Changes in migratory raptor populations in Veracruz, México, 1995–2005
Ruelas Inzunza E. 1*, L. J. Goodrich2 y S. W. Hoffman3
1 Pronatura Veracruz, Xalapa, Veracruz, 91000 México y Dartmouth College, Biology Department, Gilman Hall, Hanover, New Hampshire, EUA 03755.
2 Acopian Center for Conservation Learning, Hawk Mountain Sanctuary, Orwigsburg, Pennsylvania, EUA 17961. E–mail: goodrich@hawkmtn.org
3 HawkWatch Internacional, Salt Lake City, Utah, EUA 84115. Dirección actual: Montana Audubon, Helena, Montana, EUA 59624. E–mail: shoffman@mtaudubon.org
*Autor para correspondencia:
E–mail: ernesto.ruelas@dartmouth.edu
Recibido: 26/06/2009
Aceptado: 05/08/2010
RESUMEN
Los censos durante la migración han sido utilizados como una herramienta válida para determinar estatus poblacionales de aves rapaces. En Veracruz, México, éstos tienen la posibilidad de generar valiosa información sobre poblaciones a escala regional y continental, y de proveer cobertura para aquellas que no son estudiadas en otros sitios. Los objetivos de este trabajo son evaluar el uso de los censos durante la migración en Veracruz para seguimiento de poblaciones, documentar los ajustes hechos a un método de análisis existente para hacerlo apropiado a las propiedades específicas de nuestros datos y presentar estimaciones de cambio poblacional de las especies a lo largo de una década. Esta metodología utiliza censos de la migración de rapaces a largo plazo ajustados a esfuerzo de muestreo para generar índices anuales de abundancia. La serie de índices anuales es ajustada a una regresión polinomial para producir una estimación de la tasa de cambio anual. El uso de esta metodología de análisis para los censos en Veracruz demostró su utilidad para determinar tendencias poblacionales en muchas especies, así como limitaciones en aquellas con gran variación interanual en los censos y en especies registradas raramente. Se encontraron cambios poblacionales significativos en siete especies. Seis especies, Elanoides forflcatus, Ictinia mississippiensis, Accipiter cooperü, Buteo swainsoni, B. albonotatus y Falco peregrinus muestran incrementos a una tasa anual entre 1.9–15.7% por año–1 en el periodo 1995–2005. Una especie más, Circus cyaneus, registró descensos a una tasa de –8.4% por año–1 en el mismo periodo. Las tendencias poblacionales que se reportan aquí confirman en su mayoría incrementos poblacionales y coinciden con los datos de fuentes como los Censos de Aves Reproductoras o los Censos Navideños de Aves.
Palabras clave: Censos durante la migración, índices de población, rapaces diurnas, tendencias poblacionales.
ABSTRACT
Migration counts of raptors have been used as a valid technique to assess population levéis. Counts from Veracruz, México, have the potential of generating valuable information for regional– to continental–scale populations of some species and to provide coverage for others that are not monitored elsewhere. This paper evaluates the use of Veracruz migration counts to monitor populations, documents the adjustments made to a currently existing method to analyze migration count data and fit it to the specific properties of our dataset, and presents estimates of population change over a decade. We used long–term, effort–adjusted migration count data to genérate annual indexes of abundance. The series of annual indexes is fitted to a polynomial regression to produce an estimate of the annual rate of change. The use of this methodology with Veracruz data shows its usefulness to dataseis of many species, as well as limitations to dataseis from species with high interannual variation in migration counts, and those recorded rarely. In seven species, the authors found significant population change. Six of them, Swallow–tailed Kite, Mississippi Kite, Cooper's Hawk, Swainson's Hawk, Zone–tailedHawk, and Peregrine Falcon show increases at an annual rate between 1.9–15.7% per year–1 inthe period 1995–2005. One species, the Northern Harrier, is decreasing at a rate of –8.4% year–1 during the same period. Trends reported here reveal population increases in most cases, and concur with the pattern found in migration counts from surveys such as the Breeding Bird Survey and the Christmas Bird Count.
Key words: Diurnal raptors, migration counts, population index, trend analysis.
INTRODUCCIÓN
En Norteamérica, el estado poblacional de las aves ha sido determinado a través de varios programas de censos que varían en escala geográfica de local hasta subcontinental y se basan en diferentes técnicas de muestreo (Bart 2005). Debido a que las rapaces diurnas (Familias Cathartidae, Accipitridae y Falconidae) presentan generalmente bajas densidades y tienen hábitos esquivos, la mayoría de las especies no han sido muestreadas adecuadamente, por ejemplo, a través del Censo de Aves Reproductoras (Breeding Bird Survey, en adelante BBS, Sauer & Droege 1992; Rich et al. 2004). Como consecuencia de este problema de "detectabilidad", regiones enteras carecen de cobertura adecuada (Zalles & Bildstein 2000; Faaborg 2002).
La conspicua migración de las rapaces ofrece una oportunidad para monitorear poblaciones, si se asume que una proporción importante de la población pasa de manera consistente por un área o punto en particular. Los censos durante la migración han sido utilizados para determinar tendencias poblacionales en muchos casos (Spofford 1969; Dunne & Sutton 1986; Bednarz et al. 1990; Titus & Fuller 1990; Hoffman & Smith 2003; Bildstein et al. 2008); sin embargo, los resultados de programas de seguimiento poblacional fuera de Estados Unidos de América, Canadá y países de la Unión Europea son muy escasos, en gran medida porque los trabajos fuera de estas regiones no tienen la continuidad a largo plazo necesaria para obtener series de datos adecuadas (Kjellén & Ross 2000; Bart 2003) pero también porque las metodologías de análisis existentes no se a ajustan las particularidades de los datos de otras regiones (e.g. el uso simultáneo de múltiples sitios para conteos; para detalles ver Métodos y Apéndice 1).
Veracruz, México, puede proporcionar estimaciones valiosas sobre poblaciones de rapaces diurnas que no son monitoreadas en otras localidades (Bussjaeger et al. 1967; Thiollay 1979, 1980; Palmer 1988; Tilly et al. 1990; Ruelas et al. 2000). Para algunas especies como Cathartes aura, Ictinia mississippiensis, Buteo platypterus y Buteo swainsoni, la alta proporción de la población total registrada en esta localidad indica que un programa de seguimiento puede proporcionar información sobre su estatus a gran escala (Ruelas et al. 2010).
La principal motivación de este trabajo es desarrollar el potencial de seguimiento poblacional de aves rapaces a escala continental. Los objetivos de este artículo son (1) evaluar el uso de los censos de rapaces migratorias en Veracruz para determinar tendencias poblacionales; (2) presentar un modelo para el análisis de datos de los censos durante la migración y los ajustes necesarios para aplicar este modelo a las particularidades de los datos de esta localidad y (3) determinar las tendencias poblacionales de las especies cuyas series de datos se ajustan al modelo en uso.
MATERIAL Y MÉTODOS
Supuestos, condiciones y fundamento de los análisis. Los censos durante la migración reflejan la variación poblacional a través del tiempo. Esta señal demográfica es difícil de detectar y estimar libre de interferencia y este hecho se manifiesta en la distribución sesgada del número de aves registradas en estos censos. En estas series de datos hay pocos días en los que se registran muchos individuos y muchos días en los que se registran pocos y estos sesgos de distribución de datos dificultan el uso de modelos simples de análisis (Hussell & Brown 1992).
El uso de censos en migración para estimar cambios poblacionales está condicionado a la aceptación de dos supuestos. El primero es que estos censos representan una muestra "adecuada" de la población que migra a través de Veracruz. En este caso, se asume que: (a) no hay pseudoreplicación (los individuos registrados en un sitio son diferentes a los registrados en el segundo sitio) y (b) que los individuos censados en un día son diferentes a los muestreados en cualquier otro día (el flujo de migrantes es constante).
En Veracruz, la población migratoria de las especies es muestreada a través de dos réplicas independientes (Fig. 1, localidades números 6 y 7) y la combinación de ambas muestras puede ser utilizada en una sola cuantificación de la migración (Francis & Hussell 1998; Farmer & Hussell 2008, Apéndice 1).

En segundo lugar, se asume que esta muestra representa una proporción constante o casi constante de la población a largo plazo (Alerstam 1978). Varios estudios han criticado este supuesto, porque además del cambio demográfico los conteos tienen muchas fuentes adicionales de variación y ésta necesita ser cuantificada e incorporada en los análisis. Entre éstas, se pueden citar: (a) variables del estado del tiempo local y regional (Kerlinger & Moore 1989); (b) cambios de habitat en la periferia del lugar del censo (Dunn 2005); (c) diferencias en la temporalidad de la migración; (d) problemas de detección, cuantificación e identificación (Kerlinger 1989) y (e) diferencias entre los observadores (Satler & Bart 1984). Las tres primeras variables están asociadas al número real de individuos migrando sobre un sitio de muestreo y son de carácter extrínseco, mientras que las restantes dos variables son de carácter intrínseco y están relacionadas al número de individuos que pueden ser detectados y la metodología utilizada.
Las variables de carácter extrínseco (principalmente asociadas con el estado del tiempo), afectan incluso los conteos más sistemáticos y sólo es posible cuantificar y controlar sus efectos. En cambio, las de carácter intrínseco están relacionadas con la consistencia con la que se aplica el protocolo de toma de datos a lo largo de los años (Dunn et al. 2008), siendo posible reducir y distribuir homogéneamente sus efectos. Varios autores (Dunn & Hussell 1995; Bart et al. 2004; Dunn 2005; Hussell & Ruelas 2008) argumentan que las cuantificaciones precisas de ambos tipos de variables pueden ser incorporadas a los modelos de análisis y que un diseño de muestreo adecuado puede distribuir este error homogéneamente y reducir los sesgos. En el Apéndice 1 se muestra el método a detalle.
Hay diferentes formas de determinar las tendencias poblacionales a través del tiempo utilizando censos de la migración (Hussell 1981; Fuller & Mosher 1981, 1987; Bednarz & Kerlinger 1989; Hill & Hagan 1991; Hoffman & Smith 2003; Ballard et al. 2004). Todas son similares en su fundamento: utilizan índices anuales basados en conteos ajustados al esfuerzo de muestreo y estiman tendencias a largo plazo mediante algún modelo de regresión.
Algunas utilizan modelos lineales, aunque en series de tiempo de mayor duración las poblaciones siguen trayectorias no–lineales que oscilan por encima y por debajo de la media, e.g., en un periodo de 25 años, una población puede tener un índice de población que decrece seguido por un incremento y termina con un segundo descenso (Link & Sauer 1997).
Para mejorar la precisión de las estimaciones a largo plazo, Hussell (1981, 1985), Francis & Hussell (1998) y Farmer et al. (2007) crearon métodos que calculan índices anuales de población, reducen la magnitud de la variación a través de una serie de ajustes y transformaciones y hacen estos índices comparables a lo largo de los años. Para estimar la trayectoria de las poblaciones, utilizan un modelo de regresión polinomial que genera un mejor ajuste para cuantificar fluctuaciones en series de tiempo prolongadas. Las estimaciones de tendencias poblacionales siguiendo este procedimiento generan un resultado más realista. En el Apéndice 1 se presenta una descripción detallada del método que se utiliza en este trabajo para combinar los censos en dos localidades, generar índices anuales y finalmente calcular tendencias poblacionales.
El poder estadístico (la capacidad de detectar un cambio en parámetros de interés, si este cambio existe) ingresó a la discusión de los análisis de tendencias poblacionales cuando los investigadores del tema se dieron cuenta de las relaciones entre la variación interanual en los conteos y el esfuerzo de muestreo en la detección de tendencias y decidieron incluirlas en "metas de estimación".
Butcher et al. (1990, 1993) propusieron una "meta razonable de precisión" para detectar tendencias poblacionales en conteos navideños de aves ("Christmas Bird Counts", en adelante CBC). Esta meta se ha utilizado como un punto de referencia en discusiones subsiguientes y ha sido aplicada a otros métodos de muestreo, incluyendo datos de censos de migración (Bart et al. 2004). Sin embargo, la aplicación de esta meta de precisión a priori (la capacidad de detectar un cambio poblacional de 50% en un periodo de 25 años, con α=0.10 y β=0.20, Lewis & Gould 2000) ha sido cuestionada por varios autores (Hoenig & Heisey 2001; Farmer & Hussell 2008) para series de datos con coeficientes de variación elevados. En el caso del presente análisis, decidimos presentar los resultados junto con los valores de intervalos de confianza para evitar una determinación post hoc inútil del poder estadístico de los análisis.
Localidades, temporadas y fechas de estudio. La región central de Veracruz, México, se encuentra en la intersección de dos grandes sistemas montañosos, la Sierra Madre Oriental y el Eje Neolvolcánico, que reducen la amplitud de la planicie costera cerca de los 19° N, ca. 30 km al norte de la ciudad de Veracruz. Las faldas del volcán Cofre de Perote (a 4,250 msnm) se extienden hacia el este formando la Sierra de Manuel Díaz y llegan al Golfo de México a la altura del poblado de Villa Rica. Las rapaces migratorias prefieren migrar sobre las tierras bajas de la planicie costera donde la convección térmica facilita su migración (Ruelas et al. 2000) y este estrecho de la planicie costera forma un embudo geográfico que concentra las migraciones en primavera y otoño (Ruelas 1992; Ruelas et al. 2005) (Fig. 1).
En esta región se hicieron censos durante la migración en primavera y otoño a lo largo de varios años. Estas observaciones permitieron conocer y cuantificar la dinámica de la migración de primavera en los años 1991, 1994, 1995 y 2004, pero debido a que los patrones espacio–temporales encontrados son difusos, la colecta de información a largo plazo durante esta temporada fue descontinuada.
Las observaciones en otoño se realizaron anualmente de 1991 a 2005, del 20 de agosto al 20 de noviembre (92 días consecutivos por temporada). Para hacer las estimaciones de tendencias poblacionales presentadas en este trabajo, sólo se utilizaron los datos de los censos de otoño de 1995 a 2005 (N=10 temporadas de campo), combinando los datos colectados en dos localidades fijas a lo largo de una línea de muestreo, orientada de oeste a este y perpendicular al frente de la migración.
Las dos localidades, Cardel (parte alta del edificio de cinco pisos del Hotel Bienvenido en el centro de la ciudad, 19°22'N/–96°22'W, a 29 msnm) y Chichicaxtle (torre de observación ubicada en el campo ejidal de fútbol en el margen este del poblado, 19°21'N/–96°28'W, a 120 msnm) se encuentran a 6.5 y 17 km de distancia lineal de la costa, respectivamente (Fig. 1). Los datos de 1991–1994 y 1997 fueron excluidos de este análisis debido a que las fechas y sitios de colecta de datos difieren de los empleados en los demás años. Las observaciones son coordinadas por Pronatura Veracruz, con asistencia técnica y de capacitación por parte de Hawk Mountain Sanctuary (Kempton, Pennsylvania, EUA) y HawkWatch Internacional (Salt Lake City, Utah, EUA) y se espera continuarlas a largo plazo.
Conteos de la migración. Para obtener datos en los dos sitios, se organizaron equipos de trabajo formados por dos biólogos de campo y un asistente con entrenamiento en este trabajo, quienes hicieron 9–10 hrs diarias de observaciones de las 0800 a las 1800 h (hora estándar del centro del país, sin ajuste al horario de verano). Los observadores fueron asignados aleatoriamente a dos puntos fijos de observación; en cada sitio, estos equipos de observadores identificaron y cuantificaron los individuos migratorios de 34 especies de aves rapaces (familias Cathartidae, Accipitridae y Falconidae) y acuáticas (familias Pelecanidae, Phalacrocoracidae, Anhingidae, Ardeidae, Threskiomithidae y Ciconiidae, excluidas del presente análisis) que se registraron volando sobre el sitio, habitualmente siguiendo una trayectoria de norte a sur.
Los censos se hicieron siguiendo el protocolo estándar de la Asociación para la Migración de Rapaces de Norteamérica (HMANA por sus siglas en inglés) (Kerlinger 1989; HMANA 2009), ligeramente modificado para incluir especies que no se registran en los Estados Unidos de América y Canadá y aplicado en específico como se describe a continuación.
Se utilizaron binoculares de 8–10X para buscar e identificar migrantes y telescopios de Kowa TSN 20X montados sobre tripiés sólo para identificar aves distantes. En la identificación de aves en el campo se emplean las guías de Howell & Webb (1995), Wheeler & Clark (1995) y Sibley (2000).
En los puntos de observación, los biólogos de campo trazan una línea divisoria imaginaria utilizando una referencia (e.g. un árbol, un edificio) mediante la cual parten el cielo en dos mitades y cada uno hace observaciones de distancia lateral ilimitada con búsquedas activas en la porción que le corresponde (este u oeste). Cada hora, los observadores alternan su mitad para distribuir homogéneamente los sesgos por observador.
La función principal de los biólogos de campo es la identificación y cuantificación de las aves migratorias. Para ello, sólo se emplean observadores con al menos dos años de experiencia en este trabajo y al inicio de la temporada participan en un taller de capacitación. Los asistentes de campo tienen la función de registrar en libretas las observaciones que les son dictadas por los biólogos de campo y, en ocasiones, de participar en la identificación de individuos cercanos al sitio de observación y/o de coordinar a los biólogos de campo en el conteo y estimación. Los asistentes también están encargados de hacer la colecta de datos del clima y condiciones de observación cada hora y de mantener la comunicación por radio con el otro sitio de observación.
Para mantener las observaciones en dos sitios simultáneamente, el equipo de campo completo consiste en nueve observadores (seis biólogos de campo y tres asistentes) que son rotados entre sitios y asignados en equipos de manera aleatoria. Cada observador trabaja dos días en observaciones y tiene un día de descanso, para reducir la fatiga a lo largo de la temporada.
Los datos de ambas localidades son muestras independientes de la migración. Ambos sitios disponen de un radio de dos vías para comunicarse. La distancia lineal entre ambos sitios es de 11 km, que se estima es una distancia razonable para evitar conteos simultáneos de los mismos individuos. Aunque se han tratado de documentar conteos dobles a través de la comunicación continua entre sitios por medio de los radios, en muy pocas ocasiones un grupo de aves ha sido visible en ambos sitios y en estos casos uno de los sitios no registra la observación.
El tamaño de los grupos de migrantes no permite hacer conteos directos la mayoría de las veces, de manera que el número total de aves registrado por hora es una estimación. Los conteos directos se hacen cuando los grupos tienen menos de 50–60 aves y los observadores hacen las estimaciones cuando las aves se desplazan entre corrientes térmicas, no cuando están ascendiendo en círculos (véase a Pennycuick 1998, para una descripción del vuelo a campo traviesa). Las estimaciones se hacen utilizando un contador manual, contando directamente un grupo pequeño de aves, e.g. 10 aves, y extrapolando el área que ocupan al resto de la parvada. Los múltiplos más comúnmente utilizados es estas estimaciones son 3, 5, 10, 20 y 50 individuos. Debido a que estas parvadas frecuentemente contienen más de una especie, cada observador dispone de 6–8 contadores manuales para hacer estimaciones de igual número de especies y al final de la hora el asistente recopila los datos de cada contador. Los individuos de especies que se registran en cantidades menores son comunicados verbalmente al asistente. Al final del día, las observaciones son vaciadas en hojas de datos y capturadas en hojas de cálculo electrónicas para ser enviadas a una base de datos en línea (http://www.hawkcount.org/). Este protocolo se sigue anualmente y está disponible por escrito en un manual de campo (Pronatura Veracruz 1999).
Análisis de datos. Aunque los censos durante la migración han sido utilizados anteriormente en la estimación de tendencias poblacionales (Bednarz & Keriinger 1989; Hussell & Ruelas 2008), hay varias consideraciones especiales para interpretar los resultados de nuestro estudio de caso: (1) las aves no siguen "líneas guía" durante la migración ("leading Unes" como las definen Mueller & Berger 1967) o "líneas de divergencia" ("diversión lines" sensu Bildstein 2006), sino senderos térmicos (Schüz et al. 1971), que son espacialmente más dispersos y menos predecibles, y (2) la migración registrada es de gran magnitud (>5.2 millones de aves anuales en promedio, Cuadro 1), y consecuentemente las estimaciones del número real de individuos registrados son menos precisas que las hechas mediante conteos directos en otros sitios con migraciones de menor magnitud.

Cálculo de índices anuales de población, tendencias a largo plazo y ajustes a métodos existentes. Para el análisis de los datos, se utiliza en lo fundamental el método descrito por Farmer et al. (2007), el cual hemos modificado para poder combinar los datos de dos sitios operados simultáneamente (Apéndice 1). Este método sigue dos pasos: el primero es la estimación de un índice anual de abundancia y el segundo es la estimación de una tasa de cambio anual derivado del ajuste de los índices anuales a una regresión polinomial. Los índices de población se calcularon de manera similar a otros análisis y las tendencias de largo plazo estimadas con ligeros ajustes. Para una descripción detallada de los modelos de análisis estadísticos utilizados, véase el Apéndice 1. En todos los análisis presentados se utilizó el programa SAS, versión 9.1 (SAS Instituto 2004).
RESULTADOS
Cambios poblacionales 1995–2005. Se encontraron cambios poblacionales estadísticamente significativos en siete especies, seis de ellas con incrementos y una en descenso. En las ocho especies restantes, los censos de cuatro muestran aumentos poblacionales, dos muestran descensos y dos no registran cambios aunque ninguno de estos fue estadísticamente significativo (Cuadro 1).
A continuación se presenta el detalle de estos resultados por especie (Fig. 2) y su significado estadístico. Los nombres comunes citados son los utilizados localmente.
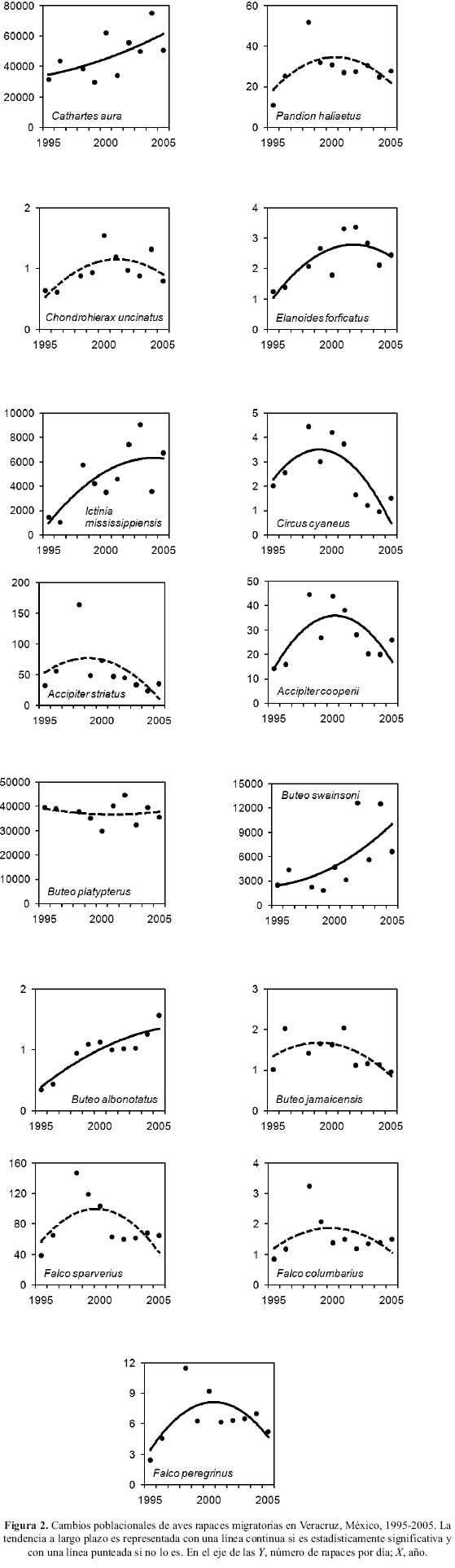
Aura Cathartes aura. Una de las especies más abundantes durante los censos en migración en Veracruz. Si bien los índices anuales registran una serie de altibajos, la variación interanual es baja (Cuadro 1, Fig. 2) y la tendencia poblacional a largo plazo es de un incremento anual elevado (r2=0.30, P=0.06).
Águila Pescadora Pandion haliaetus. Los censos muestran un incremento abrupto en los primeros tres años de la serie de datos, que se estabiliza en adelante y muestra al final un ligero decline (Fig. 2). La tasa de incremento anual no es significativa (r2=0.43, P=0.09).
Milano Pico–ganchudo Chondrohierax uncinatus. Esta especie es registrada regularmente en migración en Veracruz. Los censos anuales muestran altibajos a lo largo de la serie de datos con una tendencia creciente que no es significativa (Fig. 2; r2=0.05, P=0.25).
Milano Tijereta Elanoides forficatus. Las fechas del trabajo de campo en esta localidad dejan fuera una parte de la migración de esta especie. Los primeros milanos de la migración otoñal son registrados en Veracruz a partir de mediados de julio de cada año, por lo que una proporción desconocida queda fuera del cálculo de estos índices de población. Aun así, las estimaciones de cambio poblacional que se obtuvieron son altamente significativas (r2=0.66, P<0.01) y la trayectoria de la población muestra una tasa de incremento anual mayor al 7% que parece haberse estabilizado en años recientes (Fig. 2).
Milano Migratorio Ictinia mississippiensis. Esta es otra de las especies más abundantes en la región, que muestra una de las tasas de incremento significativo más altas de este análisis, mayor al 15% anual (Fig. 2; r2=0.52, P=0.02).
Gavilán Rastrojera Circus cyaneus. Esta especie es registrada en cantidades relativamente bajas (Cuadro 1) y es la única especie en este análisis en la que encontramos una tasa de descenso estadísticamente significativa (Fig. 2; r2=0.03, P=0.05).
Gavilán Pajarero Accipiter striatus. Este gavilán registró un aumento en los primeros tres años de la serie de datos y a partir de esa fecha los censos registraron una cantidad anual menor, que resultó en una tendencia negativa en el largo plazo. Este descenso es de dimensiones considerables, si bien ésta es sólo marginalmente significativa (Fig. 2; r2=0.35, P=0.07).
Gavilán Pollero A. cooperii. Los resultados anuales encontrados para esta especie muestran un aumento poblacional hasta la mitad de la serie de datos, que posteriormente decrece para una tasa de incremento moderada que es significativa (Fig. 2; r2=0.46, P=0.03).
Gavilán Alas–anchas Buteo platypterus. Esta es otra de las especies registradas en mayores cantidades en la región. El total anual registrado oscila considerablemente, aunque la tendencia a largo plazo es de un incremento moderado, si bien no es significativo (Fig. 2; r2=0.23, P=0.16).
Aguililla Cuaresmera B. swainsoni. La tercer especie de mayor abundancia en la región, con censos anuales que a largo plazo muestran una tasa de incremento anual considerable y estadísticamente significativa (Fig. 2; r2=0.36, P=0.03).
Aguililla Aura B. albonotatus. Esta aguililla muestra la mayor tasa de incremento poblacional anual en la serie de datos analizada y la cual es altamente significativa (Fig. 2; r2=0.75, P<0.01). Es interesante destacar que si bien el censo anual de esta especie es el menor entre todas, también es el que menos oscila interanualmente y la trayectoria poblacional muestra un patrón muy claro de incremento a largo plazo.
Aguililla Cola–roja B. jamaicensis. Otra de las especies registradas en menores cantidades en esta región, cercana al límite sur de su área de distribución durante la época no–reproductiva, que muestra una tendencia de decline moderado a largo plazo, aunque ésta no es significativa (Fig. 2; r2=0.07, P=0.21).
Cernícalo o Lili Falco sparverius. La tendencia poblacional de largo plazo del cernícalo, muy parecida a la de las siguientes dos especies de halcones, muestra un incremento durante los primeros cuatro años de la serie de datos, que sigue después una etapa de descenso y estabilización (Fig. 2). La tasa porcentual de cambio poblacional es muy cercana a cero, aunque esta no es significativa (r2=0.18, P=0.18).
Esmerejón F. columbarius. Las poblaciones de esta especie no muestran una tasa de cambio significativa (Fig. 2; r2=0.05, P=0.34).
Halcón Peregrino F. peregrinus. La tendencia de largo plazo de esta especie es de un incremento moderado, estadísticamente significativo (Fig. 2; r2=0.43, P=0.04).
Evaluación de los censos durante la migración en Veracruz para estimar tendencias poblacionales. Nueve especies raras o registradas accidentalmente fueron excluidas de los análisis por estar presentes en menos de 75% de los días de muestreo (Nij), con lo que violan los supuestos del modelo de análisis (ver Apéndice 1).
Estas especies son el Milano Cola–blanca o Chililique (Elanus leucurus, Nij=0.7, CV=150%), Milano Plomizo (Ictinia plumbea, Nij=4.1, CV=148%), Águila Calva (Haliaetus leucocephalus, Nij=0.1, CV=0.33), Aguililla Cangrejera o Guincho Prieto (Buteogallus anthracinus, Nij=2.1, CV=177%), Aguililla Conejera (Parabuteo unicinctus, Nij=6.9, CV=54%), Gavilán Gris (Buteo nitidus, Nij=180, CV=142%), Gavilán Ranero (B. lineatus, Nij=l0.l, CV=73%), Aguililla Real (B. regalis, Nij=0.6, CV=130) y Águila Real (Aquila chrysaetos, Nij=0.8, CV=167%).
La variación interanual en los censos (CV), oscila entre el 13 y el 95% (Cuadro 1), lo que indica que la precisión de las estimaciones de cambio poblacional tiene un margen amplio entre diferentes especies.
DISCUSIÓN
Censos durante la migración y tendencias poblacionales. El gran valor de los censos durante la migración de Veracruz para el seguimiento de poblaciones a escala continental se sustenta en dos argumentos principales. Por un lado, la posición geográfica de este estudio, al sur del rango reproductivo y al norte del rango no–reproductivo de la mayoría de las especies, maximiza la extensión del área de origen que puede ser muestreada en una sola localidad. Por otro lado, el volumen de la migración en Veracruz, que incluye una gran proporción de la población global total de algunas especies, permite hacer aseveraciones sobre cambios demográficos a gran escala (Ruelas et al. 2010).
Sin embargo, el modelo de análisis que se presenta aquí es limitado y se ajusta a pocas especies. Las nueve especies sin resultados fueron excluidas por dos motivos diferentes: (1) sus conteos presentan una alta variación interanual y por este motivo requieren de series de tiempo de mayor longitud para obtener trayectorias estadísticamente significativas, y (2) aquellas registradas en pocas cantidades, o distribuidas en menos de 20 días durante la temporada de migración, violan los supuestos del modelo de análisis.
Esta aparente desventaja metodológica tiene valor biológico, pues permite suprimir de los análisis a las especies que se registran marginal o raramente en la región y a las especies que por su alta variación interanual muy posiblemente se ven afectadas en mayor medida por variables extrínsecas, ajenas al cambio "real" en la señal demográfica.
Estas fuentes de variación han sido discutidas en la descripción del modelo y consideradas en los supuestos y condiciones. Sin embargo, hay tres fuentes de error intrínsecas a los censos durante la migración que conviene señalar aquí por su repercusión en la forma en que cuantifican o incluyen en los análisis: (a) errores de detección (e.g. aves pequeñas son más difíciles de detectar que las aves grandes y las especies que migran en solitario son más difíciles de detectar que las que migran en parvadas grandes), (b) errores de identificación (cerca de 2% de las aves registradas no son identificadas y este problema, que no es significativo en otras localidades, aquí significa > 100,000 aves por temporada), y (c) errores de cuantificación de grandes cantidades de migrantes (que tienen una correlación positiva con el tamaño de la parvada).
La forma de ajustar el error de estas fuentes ha sido indicada por Bart et al. (2003, 2004). Estos autores recomiendan cuantificar cada una de estas fuentes de error, e.g. el sesgo en la cuantificación por observador, e incorporar esta fuente de variación a los cálculos de los índices anuales. Por ejemplo, si el sesgo por observador (s) es estimado con un valor de ±1% del conteo, los limites superior e inferior pueden ser sustraídos/agregados a cero (el sesgo se asume con valor cero en estimaciones de poder). Con esto, es posible obtener un límite superior e inferior del sesgo, ss=–0.01 y si=0.01. Si la tasa anual de cambio es 1, sólo se puede concluir que ha ocurrido un cambio si el valor estimado es 1+ ss ó 1– si. Es posible incluir varias fuentes de error en el modelo base que presentamos aquí, y Bart et al. (2004) han desarrollado una ecuación para incorporar estos sesgos. En nuestro caso, no hemos cuantificado las fuentes de error por no disponer de los datos necesarios, pero en una futura etapa será valioso obtener los datos que permitan ajustar los datos al sesgo estimado (e.g. cuantificaciones de la variación entre observadores). Esto quizá permitirá incluir a algunas de las especies excluidas en los análisis, como algunas de las especies raras (aquellas con <100 individuos por temporada, si están apropiadamente distribuidas en tiempo).
Poder estadístico y la capacidad de alcanzar las metas de estimación. Las especies con más alta variación interanual requieren series de datos de mayor duración para detectar tendencias significativas. Si bien se reconoce que existe una relación inversa entre el poder estadístico y el valor del coeficiente de variación (Butcher 1990,1993; Lewis & Gould 2000), el análisis de poder es el que permite determinar la longitud de la serie de tiempo necesaria para detectar tendencias en especies con alta variación interanual (Bart et al. 2004). Varios autores han encontrado que estos cálculos se desempeñan mejor reportando tendencias positivas que negativas (Lewis & Gould 2000) y autores como Hoenig & Heisley (2001) han determinado que su uso no es necesario para evaluar los resultados. Por lo anterior, nuestro estudio muestra la tasa de variación anual y los intervalos de confianza para asistir la interpretación de los resultados (Cuadro 1).
Validez de las estimaciones de cambio poblacional de las especies. Para interpretar biológicamente esta información es necesario contrastar la correspondencia entre los resultados de este estudio y la información de otras fuentes. Además de las consideraciones estadísticas descritas arriba, la validación de los resultados se asiste de comparaciones cruzadas con datos de otras fuentes. James & McCulloch (1995) sugieren que para hacerlo de manera adecuada es necesario (a) contar con información sobre el origen geográfico de la población que se observa en nuestra localidad de estudio, (b) una estimación de la proporción de esta población que se muestrea en nuestros sitios de censo, (c) datos del mismo periodo de tiempo que correspondan a la misma población (en este caso, 1995–2005) y/o (d) datos de la región donde se estima que la población muestreada en Veracruz permanece durante la temporada no–reproductiva.
La información de que disponen los autores es insuficiente para realizar una comparación cuantitativa, por lo que la discusión que sigue a continuación debe ser interpretada con precaución. La mayoría de los datos vienen del BBS, cuyas limitaciones para detectar rapaces han sido discutidas anteriormente (Farmer et al. 2007; Sauer et al. 2008) y éstos son del periodo 1980–2006 (Sauer et al. 2008). Los datos del BBS que se utilizan en estas comparaciones son la tasa de cambio anual y el número de rutas de las cuales hay datos. Sauer et al. (2008) clasifican la "credibilidad" de los datos del BBS para cada especie en tres categorías, que aquí simplificamos como "baja", "media" y "alta". También se utilizan datos de los Conteos Navideños de Aves ("Christmas Bird Counts", en adelante CBC, National Audubon Society 2009), de los cuales se toman el número de individuos registrados por persona/hora y el número de rutas donde se registró la especie en el periodo 1995–2005. La pertinencia geográfica de los datos es variable, pues muchas de las especies no están presentes en el área de cobertura del CBC (en su mayoría los Estados Unidos de América y el sur de Canadá) y es clasificada en categorías similares.
Para hacer los resultados de nuestro estudio comparables con la información de las fuentes citadas arriba, dimos a los resultados de cada especie una calificación cualitativa de la confiabilidad de la información que reportamos aquí (Cuadro 2). Para hacerlo tomamos en cuenta dos criterios: el tamaño de la muestra y su variación. Dado que la abundancia de las especies tiene una distribución log–normal, el tamaño de muestra (número de individuos registrados por temporada) fue dividido en tres categorías (de 100–1,000 individuos por temporada; de 1,001 a 10,000 y más de 10,000), dándole un valor menor a las especies que se registran en menor abundancia. Por otro lado, el coeficiente de variación interanual de los censos varia de 13–95% y creamos tres categorías simples (de 1–33%, de 34–66% y >67%) en las que la categoría de mayor valor lo tienen las especies con menor variación. Así mediante la suma de estos dos criterios estimamos que los datos de especies que se registran en mayores cantidades y con variación interanual baja son más robustos que los de aquellas especies que tienen las características opuestas y las categorías de confiabilidad utilizadas son alta, media y baja; las especies con confiabilidad baja, como se explicó arriba, fueron excluidas de estos análisis. Finalmente, se hace uso de información anecdótica disponible para complementar estas comparaciones.
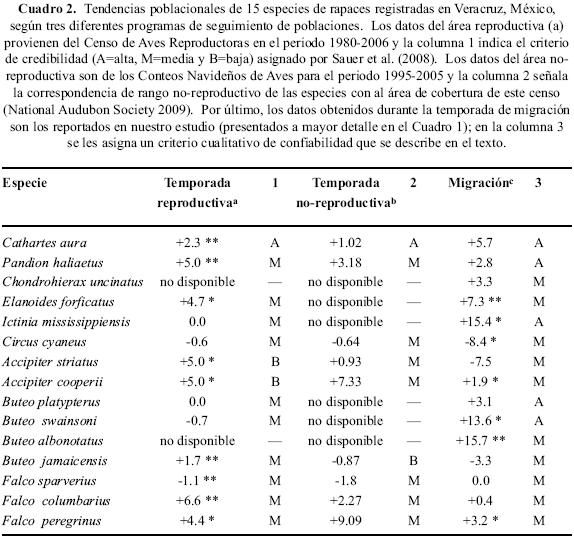
Cathartes aura. Una gran proporción de la población total de Norteamérica es observada en los dos sitios de censo en Veracruz, pero la cantidad aproximada y el origen de esa población no puede ser estimada utilizando la información existente. Es posible que esta población se origine en el centro y el sureste de Canadá, las regiones centro y este de los Estados Unidos de América y el noreste de México (Ruelas et al. 2010). La tasa anual de incremento poblacional encontrada (Cuadro 2) es mayor al doble de la estimada en rutas del BBS (P<0.001, N=2,195 rutas, Sauer et al. 2008) y es probable que sea similar en el periodo que comprende este estudio. Kirk & Mossman (1998) reportan que esta especie está expandiendo su rango rápidamente e incrementando en número, aunque no hacen ninguna estimación cuantitativa. Los CBC reportan un incremento anual de 0.53–0.59 individuos por persona/hora (National Audubon Society 2009), que indica que el número de Auras que permanece en Norteamérica en la temporada no–reproductiva también se ha incrementado, si bien de una manera moderada.
Pandion haliaetus. La población de esta especie que migra por Veracruz es marginal (Cuadro 1) y estas aves parecen originarse en localidades a todo lo ancho de Norteamérica (Martell et al. 2001; Elliot et al. 2007), por lo que es difícil encontrar la correspondencia específica entre las áreas de origen de estas aves y los datos de abundancia de su rango reproductivo. Aunque los resultados de este análisis no son estadísticamente significativos, estos registran un incremento. El Águila Pescadora está aumentando significativamente en toda su área de distribución (Poole et al. 2002; Hoffman & Smith 2003; Farmer et al. 2008a, 2008b) y el BBS reporta incrementos para el periodo 1980–2006 (P<0.00l, N=453, Sauer et al. 2008), mientras que los CBC registran incrementos moderados, de 0.020–0.027 individuos por persona/hora de observación (National Audubon Society 2009), que indican que el numero de aves que permanecen en Norteamérica durante la temporada no–reproductiva se ha mantenido relativamente constante.
Chondrohierax uncinatus. Antes de nuestras observaciones en Veracruz, esta especie era considerada como sedentaria y se conocían movimientos altitudinales y otros desplazamientos irregulares (AOU 1998). El trabajo previo de los autores (Ruelas et al. 2002) ha documentado su estatus como una especie que migra en un eje norte a sur a lo largo de la planicie del Golfo y cuyas poblaciones se originan en el sur de Texas y/o en el noreste de México. No existen datos poblacionales de BBS o CBC para hacer comparaciones poblacionales.
Elanoides forficatus. El origen geográfico de las poblaciones que migran por Veracruz es posiblemente la porción este de Texas, Louisiana, e incluso Alabama o el oeste de Florida (Meyer 1995). Otros autores han documentado su migración a través del Golfo de México, de Florida a Cuba y es posible que la mayoría de la subespecie norteña utilice esta ruta migratoria.
Los conteos de migración en Veracruz inician el 20 de agosto de cada año y una buena parte de la migración de esta especie no es cuantificada en esta localidad. Aun así, los incrementos significativos detectados (Cuadro 1), coinciden con datos de otras fuentes. El BBS reporta incrementos (P=0.02, N=53, Sauer et al. 2008). Los CBC no registran datos de valor para esta especie (National Audubon Society 2009).
Ictinia mississippiensis. Aunque es probable que más del 95% de la población total de esta especie migre a través de Veracruz (Ruelas et al. 2010) y en ésta se encontró uno de los incrementos anuales más significativos de este estudio (Cuadro 1), estos datos sólo pueden comprarse con valores del BBS, que curiosamente no reportan ninguna tendencia poblacional (P=0.97, N=192, Sauer et al. 2008). Dado que estos milanos están ausentes de su rango reproductivo en la temporada invernal casi por completo, el CBC no tiene datos para comparaciones (National Audubon Society 2009). Parker (1999) coincide en que las poblaciones y el rango de esta especie se están expandiendo, pero aporta poca información cuantitativa para comparar con los datos que se obtuvieron en este estudio.
Circus cyaneus. La proporción de la población de Norteamérica que es muestreada en Veracruz es mínima. Debido al amplio rango de distribución y a la ausencia de información detallada sobre la migración de esta especie, no es posible hacer comparaciones precisas sobre el origen de los migrantes que se registran en Veracruz.
Esta especie registra el declive más significativo de todas las especies (Cuadro 1). Sin embargo, datos de los censos durante la migración en Veracruz no parecen estar relacionados con lo que ocurre con sus poblaciones (Cuadro 2), porque en una comparación con datos del BBS para todo el rango de distribución reproductiva, estos no reportan descensos significativos (P=0.13, N=979, Sauer et al. 2008). De igual manera, el CBC arroja una disminución en el número de aves reportadas por ruta (0.14–013 individuos por persona/hora) (National Audubon Society 2009). Otros trabajos muestran descensos regionales de manera anecdótica (MacWhirter & Bildstein 1996).
Accipiter striatus. Con poblaciones marginales migrando por la región, esta especie muestra descensos de gran magnitud, aunque estos no son significativos (Cuadro 1). No existe información sobre la región de origen de estas poblaciones, pero en base a recapturas de individuos anillados y en base a datos morfométricos, estos se originan en la región central y el este de los Estados Unidos de América (Grandes Lagos) (Ruelas 2007). Es posible que los descensos detectados en nuestro trabajo obedezcan a cambios en la distribución durante la temporada no–reproductiva más que a cambios demográficos, pues esta especie se registra regularmente en rutas del BBS y en ellos se observan incrementos, aunque estos censos aportan información limitada por su dificultad de detección (P=0.03, N=304, Sauer et al. 2008). Hay evidencia de que esta especie está cambiando sus patrones de migración en años recientes (Viverette et al. 1996; Duncan 1996), permaneciendo en localidades más norteñas. Esto puede verse reflejado en datos de dos fuentes: en la tasa de encuentros en CBC, los cuales han incrementado ligeramente (0.039–0.043 individuos por persona/hora, National Audubon Society 2009) y en la frecuencia de visitas de esta especie en comederos de aves en Estados Unidos de América y Canadá, que también se ha incrementado (D.N. Bonter, Project FeederWatch, com. pers.).
A. cooperii. A diferencia de su congénere, esta especie registró una tasa de incremento significativa (Cuadro 1). No existe información detallada sobre el origen geográfico de las poblaciones registradas en Veracruz, pero en base a recapturas de algunos individuos en la estación de anillamiento local, es muy probable que sean similares a las de A. striatus. Varios autores reportan incrementos de manera anecdótica y las rutas del BBS registran incrementos (P=0.01, N=592). Es posible que sus poblaciones también se encuentren cambiando sus patrones de migración y que estén invernando más al norte (La Sorte & Thompson 2007). Los datos del CBC (0.031–0.056 individuos por persona/hora, National Audubon Society 2009) y registros en comederos de aves muestran incrementos (D.N. Bonter, Project FeederWatch, com. pers.).
Buteo platypterus. Es probable que la mayoría de la población global de esta especie (<95%) migre por Veracruz (Ruelas et al. 2010). La tasa de incremento anual registrada para esta especie no es significativa (Cuadro 1). No hay información disponible para contrastar este valor, pues en rutas del BBS no hay cambio registrado (P=n.s., N=696) y debido a que es una migratoria total, no se registra en CBC (<25 conteos, National Audubon Society 2009).
B. swainsoni. Al igual que la especie anterior, la mayoría de la población global de esta especie migra por Veracruz. Ésta registra la tercera tasa de crecimiento anual más alta (Cuadro 1), que contrasta con un ligero descenso, no significativo, reportado por el BBS (P=0.08, N=683, Sauer 2008). Debido a que es una migratoria total, se registra escasamente en CBC (<25 rutas, National Audubon Society 2009).
B. albonotatus. Las poblaciones que se registran en Veracruz posiblemente se originen en el noreste de México (Johnson et al. 2000). Esta especie muestra la tasa de crecimiento más alta de este estudio (Cuadro 1), aunque es difícil encontrar la correspondencia entre este valor con otras fuentes. El BBS y el CBC no muestran datos (Cuadro 2). Anecdóticamente aparece expandiendo su rango y aumentando sus poblaciones (Snyder & Glinski 1988; Snyder 1998).
B.jamaicensis. El origen geográfico de la población que se registra en Veracruz no está determinado, pero posiblemente se trate de poblaciones del noreste de México y el sur de los Estados Unidos de América, pues es una "migratoria en cadena" (Bildstein 2006). La tasa de descenso encontrada en este estudio no es significativa. La información anecdótica y cuantitativa muestra incrementos en esta especie. En base a datos del BBS (P<0.00l, N=2,996, Sauer et al. 2008) y en CBC (0.52–0.47 individuos por persona/hora, National Audubon Society 2009), varios autores especulan que esta especie está cambiando sus hábitos migratorios, permaneciendo en regiones más norteñas durante la época no reproductiva y consecuentemente descendiendo sus valores en los censos migratorios en localidades meridionales (Farmer et al. 2008b).
Falco sparverius. Sin información detallada sobre el origen geográfico de las poblaciones registradas en Veracruz, los censos de esta localidad no reportan cambios. Esta especie experimenta un descenso en todo su rango de distribución (Farmer & Smith 2010), en rutas del BBS (P=<0.001, N=2,349, Sauer et al. 2008) y en CBC (0.29–0.23 individuos por persona/hora, National Audubon Society 2009).
F. columbarius. No se tiene conocimiento del origen geográfico de la modesta población de esta especie que se registra en Veracruz. Los resultados de estos censos señalan un incremento mínimo y éste no es estadísticamente significativo (Cuadro 1). Varios autores (Farmer et al. 2008a, 2008b) reportan anecdótica y cuantitativamente incrementos notables en sus poblaciones. El BBS registra incrementos (P<0.001, N=156, Sauer et al. 2008). Estos incrementos se extienden a conteos durante la temporada no–reproductiva, el CBC reporta aumentos notables (0.008–0.01 individuos por persona/hora, National Audubon Society 2009).
F. peregrinus. Las poblaciones de este halcón registradas en la región tienen origen geográfico desconocido. En estas se registra un incremento anual mediano y estadísticamente significativo (Cuadro 1), que corresponde a lo mostrado en otros censos como las rutas del BBS (P=0.04, N=26, Sauer et al. 2008), en CBC (0.003–0.006 individuos por persona/hora, National Audubon Society 2009) y en numerosas referencias tanto anecdóticas como cuantitativas (White et al. 2002).
En síntesis, este trabajo demuestra que los censos migratorios en Veracruz pueden ser utilizados para determinar tendencias poblacionales en las especies que se ajustan a los supuestos y condiciones de la metodología. Para mejorar el alcance de este modelo de análisis, los autores identifican varios puntos para mejorarlo y ajustarlo a las cualidades de los datos de las especies raras y de aquellas que son muy abundantes.
Nuestros resultados ilustran una visión de la dinámica demográfica de varias especies de rapaces migratorias que muestra aumentos significativos en la gran mayoría de las especies. La información obtenida de diferentes fuentes es, en su mayoría, coherente entre sí (Cuadro 2). Aunque las tasas de cambio poblacional reportadas son variables, la tendencias de incremento o decline son virtualmente las mismas (con dos excepciones, discutidas en el texto). Individualmente, los BBS muestran ocho especies con incrementos, una sin cambios, dos con declines y dos sin datos disponibles. Por su parte, los CBC reportan seis especies con incrementos, cero sin cambios, tres con declines y seis sin datos disponibles. Finalmente, los datos de nuestro estudio reportan 11 especies con incrementos, una sin cambios y dos con declines.
Esto contrasta con los hallazgos de reportes generales para otros grupos de aves que muestran más declines que incrementos (e.g. Rich et al. 2004; NABCI et al. 2010). ¿Cuáles pueden ser las razones que expliquen mayores incrementos en las rapaces que en otros grupos taxonómicos? Es posible que los factores que limitaban selectivamente a las poblaciones de rapaces en el pasado, como la contaminación y la caza directa (Sergio et al. 2005) hayan sido reducidas como amenazas significativas y que las rapaces estén regresando a niveles poblacionales "normales". Además de cambio demográfico, los resultados de los conteos pueden reflejar otros procesos de gran escala como el cambio climático reciente, que altera de manera significativa los patrones de distribución geográfica y temporal de la migración de las aves (Møller et al. 2006; Both et al. 2006; La Sorte & Thompson 2007; Miller–Rushing et al. 2008). La determinación precisa de los procesos que regulan los cambios poblacionales presentados en este artículo es sin duda una de las mayores prioridades de futuras investigaciones.
AGRADECIMIENTOS
Queremos agradecer a Ricardo Rodríguez Estrella y a dos revisores anónimos sus comentarios al manuscrito. Christopher J. Farmer, David J.T. Hussell y Alfonso Godino contribuyeron también a desarrollar este trabajo y a preparar esta publicación. También agradecemos a más de 130 biólogos de campo y otros colaboradores del proyecto Veracruz Río de Rapaces. A través de los años, este proyecto ha sido financiado por National Fish and Wildlife Foundation, John D. y Catherine T. MacArthur Foundation, el Acta de Aves Migratorias Neotropicales del U.S. Fish and Wildlife Service, Center for the Study of Tropical Birds, American Bird Conservancy, American Birding Association, Lannan Foundation (Patrick y Andy Lannan, Bill Johnston y Mary Pat Day), The Nature Conservancy, Fondo Mexicano para la Conservación de la Naturaleza, The International Osprey Foundation, National Audubon Society, Rare Conservation y University of Missouri (John Faaborg y Josh Millspaugh, Conservation Biology Summer Fellowship 2003). ERI recibió una beca postdoctoral del Howard Hughes Medical Institute en Dartmouth College y completó este manuscrito mientras trabajaba en el laboratorio de Matthew P. Ayres. Esta publicación es la contribución de ciencias de la conservación No. 193 de Hawk Mountain Sanctuary.
LITERATURA CITADA
Alerstam, T. 1978. Analysis and theory of visible migration. Oikos, 30:273–349. [ Links ]
Alien, P.E., L.J. Goodrich, & K.L. Bildstein 1996. Within– and among–year effects of cold fronts on migrating raptors at Hawk Mountain, Pennsylvania, 1934–1991. The Auk, 113:329-338. [ Links ]
American Ornitholologists' Unión (AOU). 1998. Checklist of North American birds. 6a. Edición.American Ornithologists' Unión, Washington, D.C. [ Links ]
Ballard, G., G.R. Geupel, N. Nur, & T. Gardali. 2004. Long–term declines and decadal patterns in population trends of songbirds in western North América, 1979–1999. The Cóndor, 105:737–755. [ Links ]
Bart, J. 2003. Raptors and the North American Coordinated Bird Monitoring Plan. Hawk Migration Studies, 29:11–13. [ Links ]
Bart, J. 2005. Monitoring the abundance of bird populations. The Auk, 122:15–25. [ Links ]
Bart, J., B. Collins, & R.I.G. Morrison. 2003. Estimating population trends with a linear model. The Cóndor, 105:367–372. [ Links ]
Bart, J., K.P. Burnham, E.H. Dunn, C.M. Francis, & C.J. Ralph. 2004. Goals and strategies for estimating trends in landbird abundance. Journal of Wildlife Management, 68:611–623. [ Links ]
Bednarz, J.C. & P. Kerlinger. 1989. Monitoring hawk populations by counting migrants. Pp. 328–342. In B.G. Pendleton, M.N. LeFranc, Jr., M.B. Moss, CE. Ruibal, M.A. Knighton, & D.L. Krahe (eds). Proceedings ofthe Northeast Raptor Management Symposium and Workshop. Institute for Wildlife Research and National Wildlife Federation, Washington, D.C. [ Links ]
Bednarz, J.C, D. Klem, Jr., L.J. Goodrich, & S.E. Senner. 1990. Migration counts of raptors at Hawk Mountain, Pennsylvania, as indicators of population trends, 1934–1986. The Auk, 107:96–109. [ Links ]
Bildstein, K.L. 2006. Migrating raptors ofthe world, their ecology and conservation. Cornell University Press, Ithaca, Nueva York. [ Links ]
Bildstein, K.L., J.P. Smith, E. Ruelas I., & R.R. Veit (eds). 2008. State of North America's birds of prey. Nuttall Ornithological Club & American Ornithologists' Unión Series in Ornithology No. 3, Cambridge, Massachusetts. [ Links ]
Both, C, S. Bouwhuis, C.M. Lessells, & M.E. Visser. 2006. Climate change and population declines in a long–distance migratory bird. Nature, 44:81–83. [ Links ]
Burnham, K.P. & D.R. Anderson. 2002. Model selection and multimodel inference; A practical information–theoretic approach. Second Edition. Springer, Nueva York. [ Links ]
Bussjaeger, L. J., C C Carpenter, H. L. Cleveland, & D. L. Marcellini. 1967. Turkey Vulture migration in Veracruz, México. The Cóndor, 69:425–426. [ Links ]
Butcher, G., M. Fuller, L.S. McAUister, & P.H. Geissler. 1990. An evaluation of Christmas Bird Count for monitoring population trends of selected species. Wildlife Society Bulletin, 18:129–134. [ Links ]
Butcher, G., B.G. Peterjohn, & CJ. Ralph. 1993. Overview of national bird population monitoring programs and databases. Pp. 192–203. In D.M. Finch & PW. Stangel (eds). Status and management of Neotropical migratory birds. U.S. Forest Service Rocky Mountain Forest and Range Experiment Station GTR–RM–229, Estes Park, Colorado. [ Links ]
Duncan, C.D. 1996. Changes in the winter abundance of Sharp–shinned Hawks in New England. Journal ofField Ornithology, 67:254–262. [ Links ]
Dunn, E.H. 2005. Counting migrants to monitor bird populations: state ofthe art. Pp. 712–717. In C.J. Ralph, & T.D. Rich (eds). Bird conservation implementation and integration in the Americas. Proceedings ofthe Third International Partners in Flight Conference. USDA Forest Service Gen. Tech. Rep. PSW–GTR–191, Albany, California. [ Links ]
Dunn, E.H. & D.J.T. Hussell. 1995. Using migration counts to monitor landbird populations: review and evaluation of current status. Current Ornithology, 12:43–88. [ Links ]
Dunn, E.H., D.J.T. Hussell, & E. Ruelas I. 2008. Recommended methods for population monitoring at raptor–migration watchsites. Pp. 447–460. In K.L. Bildstein, , J.P. Smith, E. Ruelas I., & R.R. Veit (eds). State ofNorth America's birds ofprey. Nuttall Ornithological Club & American Ornithologists' Unión Series in Ornithology No. 3, Cambridge, Massachusetts. [ Links ]
Dunne, P. & C. Sutton. 1986. Population trends of coastal raptor migrants over ten years of Cape May Point autumn counts. Records of New Jersey Birds, 12:39–43. [ Links ]
Elliot, J.E., C.J. Henny, C.A. Morrissey, E. Ruelas I., & P. Shaw. 2007. Satellite telemetry and prey sampling reveal contaminant sources to Pacific Northwest Ospreys. Ecológica! Applications, 17:1223–1233. [ Links ]
Faaborg, J. 2002. Saving migrant birds, developing strategies for thefuture. University of Texas Press, Austin, Texas. [ Links ]
Farmer, C.J., D.J.T. Hussell, & D. Mizrahi. 2007. Detecting population trends in migratory birds of prey. TheAuk, 124:1047–1062. [ Links ]
Farmer, C.J. & D.J.T. Hussell. 2008. The Raptor Population índex in practice. Pp. 165–178. In K.L. Bildstein, J.P. Smith, E. Ruelas I., & R.R. Veit (eds). State of North America's birds ofprey. Nuttall Ornithological Club & American Ornithologists' Unión Series in Ornithology No. 3, Cambridge, Massachusetts. [ Links ]
Farmer, C.J., L.J. Goodrich, D.J.T. Hussell, R. Bell, B. Drolet, E. Greenstone, D. Grove, D. Mizrahi, F.J. Nicoletti, & J. Sodergren. 2008a. Trends in autumn counts of migratory raptors in northeastern North América, 1974–2004. Pp. 179–216. In K.L. Bildstein, J.P. Smith, E. Ruelas I., & R.R. Veit (eds). State of North America's birds of prey. Nuttall Ornithological Club & American Ornithologists' Unión Series in Ornithology No. 3, Cambridge, Massachusetts. [ Links ]
Farmer, C.J., L.J. Goodrich, E. Ruelas I., & J.P. Smith. 2008b. Conservation status reports. Pp. 303– 420. In K.L. Bildstein, J.P. Smith, E. Ruelas I., & R.R. Veit (eds). State ofNorth America's birds of prey. Nuttall Ornithological Club & American Ornithologists' Unión Series in Ornithology No. 3, Cambridge, Massachusetts. [ Links ]
Farmer, C.J. & J.P. Smith. 2009. Migration monitoring indicates widespread declines of American Kestrels {Falco sparverius) in North América. Journal of Raptor Research, 43:263–273. [ Links ]
Francis, C.M. & D.J.T. Hussell. 1998. Changes in numbers of land birds counted in migration at Long Point Bird Observatory, 1961–1997. Bird Populations, 4:37–66. [ Links ]
Fuller, M.R. & J.A. Mosher. 1981. Methods for detecting and counting raptors: A review. Studies in Avian Biology, 6:235–246. [ Links ]
Fuller, M.R. & J.A. Mosher. 1987. Raptor survey techniques. Pp. 37–65. In B.G. Pendleton, M.N. LeFranc, Jr., M.B. Moss, CE. Ruibal, M.A. Rnighton, & D.L. Rrahe (eds). Proceedings of the Northeast Raptor Management Symposium and Workshop. Institute for Wildlife Research and National Wildlife Federation, Washington, D.C. [ Links ]
Haugh, J.R. 1972. A study of hawk migration in eastern North América. Search, 2:1–60. [ Links ]
Hill, N.P. &J.M. Hagan, III. 1991. Population trends of some northeastern North American landbirds: half–century of data. Wilson Bulletin, 103:165–338. [ Links ]
Hawk Migration Association of North América (HMANA). 2009. Data sheets and protoco Is for data collection (http://www.hmana.org/drfl.xls, 1 junio 2009). [ Links ]
Hoenig, J.M. & D.M. Heisley. 2001. The abuse of power: the pervasive fallacy of power calculations for data analysis. American Statistician, 55:19–24. [ Links ]
Hoffman, S.W. & J.P. Smith. 2003. Population trends of migratory raptors in western North América, 1997–2001. The Cóndor, 105:397–419. [ Links ]
Howell, S.N.G. & S. Webb. 1995. A guide to the birds of México and northern Central América. Oxford University Press, Oxford, Reino Unido. [ Links ]
Hussell, D.J.T. 1981. The use of migration counts for monitoring bird population levéis. Studies in Avian Biology, 6:92–102. [ Links ]
Hussell, D.J.T. 1985. Analysis of hawk migration counts for monitoring population levéis. Pp. 243–254. In M. Harwood (ed). Proceedings of Hawk Migration Conference IV. Hawk Migration Association of North América, North Wales, Pennsylvania. [ Links ]
Hussell, D.J.T. & L. Brown. 1992. Population changes in diurnally–migrating raptors at Duluth, Minnesota (1974–1989) and Grimsby, Ontario (1975–1990). Ontario Ministry of Natural Resources, Maple, Ontario, Canadá [ Links ].
Hussell, D.J.T. & E. Ruelas I. 2008. The Raptor Population índex in principie. Pp. 151–164. In K.L. Bildstein, J.P. Smith, E. Ruelas I., & R.R. Veit (eds). State of North America's birds ofprey. Nuttall Ornithological Club & American Ornithologists' Unión Series in Ornithology No. 3, Cambridge, Massachusetts. [ Links ]
James, F.C. &CE. McCulloch. 1995. The strength of inferences about causes of trends in populations. Pp. 40–84. In T.E. Martin, & D.M. Finch (eds). Ecology and management ofNeotropical migratory birds: synthesis and review of critica! issues. Oxford University Press, Nueva York & Londres. [ Links ]
Johnson, R.R., R.L. Glinski, & S.W. Matteson. 2000. Zone–tailed Hawk (Buteo albonotatus). In A. Poole & F. Gilí (eds). The Birds of North América No. 529. The Birds of North América, Inc, Philadelphia, Pennsylvania. [ Links ]
Kerlinger, P. 1989. Flight strategies of migrating hawks. University of Chicago Press, Chicago, & Londres. [ Links ]
Kerlinger, P. & F.R. Moore. 1989. Atmospheric structure and avian migration. Current Ornithology, 6:109–142. [ Links ]
Kirk, D.A. & M.J. Mossman. 1998. Turkey Vulture (Cathartes aura). In A. Poole & F. Gilí (eds). The Birds of North América No. 339. The Birds of North América, Inc, Philadelphia, Pennsylvania. [ Links ]
Kjellén, N. & G. Ross. 2000. Population trends in Swedish raptors demonstrated by migration counts at Falsterbo, Sweden 1942–1997. Bird Study, 47:195–211. [ Links ]
La Sorte, F. A. &Thompson, F. R. (2007). Poleward shifts in winter ranges of North American birds. Ecology, 88:1803–1812. [ Links ]
Lewis, S.A. & W.R. Gould. 2000. Survey effort effects on power to detect trends in raptor migration counts. Wildlife Society Bulletin, 28:317–329. [ Links ]
Link, W.A. & J.R. Sauer. 1997. Estimation of population trajectories from count data. Biometrics, 53:488–497. [ Links ]
Martell, M.S., C J. Henny, P.E. Nye, & M.J. Solensky. 2001. Fall migration routes, timing, and wintering sites of North American Ospreys as determinedby satellite telemetry. The Cóndor, 103:715–724. [ Links ]
MacWhirter, R.B. & K.L. Bildstein. 1996. Northern Harrier (Circus cyaneus). In A. Poole, & F. Gilí (eds). The Birds of North América No. 210. The Academy of Natural Sciences & The American Ornithologists' Unión, Philadelphia, Pennsylvania. [ Links ]
Meyer, K.D. 1995. Swallow–tailed Kite (Elanoides forficatus). In A. Poole & F. Gilí (eds). The Birds of North América No. 138. The Academy of Natural Sciences & The American Ornithologists' Unión, Philadelphia, Pennsylvania. [ Links ]
Miller–Rushing, A.J., T.L. Lloyd–Evans, R.B. Primack, & P. Satzinger. 2008. Bird migration times, climate change, and changing population sizes. Global Change Biology, 14:1959–1972. [ Links ]
Maller, A. P., W. Fiedler, & P. Berthold (eds). 2006. Birds and climate change. Academic Press, Amsterdam, Holanda. [ Links ]
Mueller, H.C. & D.D. Berger. 1961. Weather and fall migration of hawks at Cedar Grove, Wisconsin. Wilson Bulletin, 73:171–192. [ Links ]
Mueller, H.C. & D.D. Berger. 1967. Wind drift, leading lines, and diurnal migrations. Wilson Bulletin, 79:50–63. [ Links ]
National Audubon Society. 2009. Christmas Bird Count historical results (http://www.audubon.org/bird/cbc/, 1 junio 2009). [ Links ]
North American Bird Conservation Inititative (NABCI), American Bird Conservancy, Association of Fish and Wildlife Agencies, National Audubon Society, Cornell Laboratory of Ornithology, Klamath Bird Observatory, The Nature Conservancy, United States Geological Survey, Partners in Flight, United States Shorebird Conservation Plan, North American Waterfowl Conservation Plan, & Waterbird Conservation of the Americas. 2010. The state ofthe birds, 2010 report on climate change (http://www.stateofthebirds.org/, 15 junio 2010). [ Links ]
Newton, I. 1979. Population ecology of raptors. Buteo Books, Vermillion, South Dakota. [ Links ]
Palmer, R. S. 1988. Broad–winged Hawk. Pp. 3–133. In R.S. Palmer (ed). Handbookof North American Birds, Vol. 5. Yale University Press, New Haven, Connecticut. [ Links ]
Parker, J.W. 1999. Mississippi Kite (Ictinia mississippiensis). In A. Poole, & F. Gilí (eds). The Birds of North América No. 402. The Birds of North América, Inc, Philadelphia, Pennsylvania. [ Links ]
Pennycuick, C.J. 1998. Field observations of thermals and thermal streets, and the theory of cross–country soaring flight. Journal of Avian Biology, 29:33–43. [ Links ]
Poole, A.F., R.O. Bierregaard, & M.S. Martell, 2002. Osprey (Pandion haliaetus). In A. Poole & F. Gilí (eds). The Birds of North América No. 683. The Birds of North América, Inc. Philadelphia, Pennsylvania. [ Links ]
Pronatura Veracruz. 1999. Veracruz Río de Rapaces: protocolo para conteos de migración. Pronatura Veracruz & National Fish and Wildlife Foundation, Xalapa, Veracruz, México. [ Links ]
Rich, T.D., C.J. Beardmore, H. Berlanga, PJ. Blancher, M.S.W. Bradstreet, G.S. Butcher, D.W. Demarest, E.H. Dunn, W.C. Hunter, E.E. Iñigo–Elias, J.A. Kennedy, A.M. Martell, A.O. Panjabi, D.N. Pashley, K.V. Rosenberg, C.M. Rustay, J.S. Wendt, & T.S. Will. 2004. Partners in Flight North American Landbird Conservation Plan. Cornell Laboratory of Ornithology, Ithaca, Nueva York. [ Links ]
Richardson, W.J. 1978. Timing and amount of bird migration en relation to weather: a review. Oikos, 30:224–272. [ Links ]
Ruelas I., E. 1992. Mexico region; hawk migration regional report. Hawk Migration Studies, 17:43–45. [ Links ]
Ruelas I., E. 2007. Raptor and wading bird migration in Veracruz, México: Spatial and temporal dynamics, flight performance, and monitoring applications. Tesis doctoral. University of Missouri.Columbia, Missouri. [ Links ]
Ruelas I., E., L.J. Goodrich, S.W. Hoffman, & R.Tingay. 2000. Conservation strategies for the World's largest raptor migration flyway: Veracruz, theRiverof Raptors. Pp. 591–596. In R.D. Chancellor, & B.–U. Meyburg (eds). Raptors atRisk. Hancock House Publishers, Surrey, B.C., Canadá [ Links ].
Ruelas I., E., J.E. Montejo D., S.W. Hoffman, & L.J. Goodrich 2002. The migration ofthe Hookbilled Kite (Chondrohierax uncinatus). Memorias de la III North American Ornithological Conference. New Orleans, Louisiana. [ Links ]
Ruelas I., E., S.W. Hoffman, & L.J. Goodrich. 2005. Stopover ecology of Neotropical migrants Veracruz, México. Pp. 657–673. In C.J. Ralph, & T.D. Rich (eds). Bird conservation implementation and integration in the Americas: Proceedings ofthe Third International Partners in Flight Conference. Vol. 2. Gen. Tech. Rep. PSW–GTR–191. Pacific Southwest Research Station, Forest Service. U.S. Department of Agriculture, Albany, California. [ Links ]
Ruelas I., E., L.J. Goodrich, & S.W. Hoffman. 2010. North American population estimates of water–birds, vultures, and hawks from migration counts in Veracruz, México. Bird Conservation International, 20:124–133. [ Links ]
SAS Institute. 2004. SAS. Versión 9.1. The SAS Institute, Cary, North Carolina. [ Links ]
Satler, G. & J. Bart. 1984. Reliability of counts of migrating raptors: an experimental analysis. Journal of Field Ornithology, 55:415–537. [ Links ]
Sauer, J.R. & S. Droege. 1992. Geographical patterns in population trends of Neotropical migrants in North América. Pp. 26–42. In J.M. Hagan & D.W. Johnston (eds). Ecology and conservation of Neotropical migrant landbirds. Smithsonian Institution Press, Washington, D.C. [ Links ]
Sauer, J.R., J.E. Hiñes, & J. Fallón. 2008. The North American Breeding Bird Survey, Results and Analysis 1966 – 2004. Versión 2005.2. USGS Patuxent Wildlife Research Center, Laurel, Maryland (http://www.mbr–pwrc.usgs.gov/bbs/bbs.html/, 1 septiembre 2008). [ Links ]
Sergio, F., I. Newton, & L. Marchesi. 2005. Top predators and biodiversity. Nature, 436:192. [ Links ]
Sibley, D.A. 2000. The Sibley guide to the birds. Alfred A. Knopf, Nueva York. [ Links ]
Schüz, E., P. Berthold, E. Gwinner, & H. Oelke. 1971. Grundri, der Vogelzugskunde. Parey Books, Berlin & Hamburgo, Alemania. [ Links ]
Smith, J.P., C.J. Farmer, S.W. Hoffman, C.A. Lott, L.J. Goodrich, J. Simón, C.Riley, & E. Ruelas I. 2008a. Trends in autumn counts of migratory raptors around the Gulf of México. Pp. 217–252. In K.L. Bildstein, J.P. Smith, E. Ruelas I., & R.R. Veit (eds). State of North America's birds ofprey. Nuttall Ornithological Club & American Ornithologists' Unión Series in Ornithology No. 3, Cambridge, Massachusetts. [ Links ]
Smith, J.P., C J. Farmer, S.W. Hoffman, G.S. Kaltenecker, K.Z. Woodruff, & P.F. Sherrington. 2008b. Trends in counts of migratory raptors in western North América. Pp. 253–278. In K.L. Bildstein, J.P. Smith, E. Ruelas I., & R.R. Veit (eds). State of North América 's birds ofprey. Nuttall Ornithological Club & American Ornithologists' Unión Series in Ornithology No. 3, Cambridge, Massachusetts. [ Links ]
Snyder, H.A. 1998. Zone–tailed Hawk. Pp. 99–101. In R.L. Glinski (ed.) The raptors ofArizona. Univ. of Arizona Press, Tucson & Arizona Game and Fish Department, Phoenix, Arizona. [ Links ]
Snyder, H.A. & R.L. Glinski. 1988. Zone–tailed Hawk. Pp. 105–110. In R.L. Glinski, B.G. Pendleton, M.B. Moss, M.N. LeFranc, Jr., B.A. Millsap, & S.W. Hoffman (eds). Proceedings of the Southwest Raptor Management Symposium and Workshop. Scientific Technical Series No. 11. National Wildlife Federation, Washington, D.C. [ Links ]
Spofford, W.R. 1969. Hawk Mountain counts as population indices in northeastern North América. Pp. 323–332. In J.J. Hickey, (ed.) Peregrine Falcon populations: their biology and decline. University of Wisconsin Press, Madison, Wisconsin. [ Links ]
Tabachnick, B.G. & L.S. Fidell. 1989. Using multivariate statistics. Segunda edición. Harper Collins, Nueva York. [ Links ]
Thiollay, J.–M. 1979. Importance of an axis of migration along the east coast of México. Alauda, 47:235–246. [ Links ]
Thiollay, J.–M. 1980. Spring hawk migration in eastern México. Raptor Research, 14:13–19. [ Links ]
Tilly, F.C., S. W. Hoffman, & C. R. Tilly. 1990. Spring hawk migration in southern México. Hawk Migration Studies, 15:21–29. [ Links ]
Titus, K. & M.R. Fuller. 1990. Recent trends in counts of migrant hawks from north–eastern North América. Journal of Wildlife Management, 54:463–470. [ Links ]
Viverette, C.B., S. Struve, L.J. Goodrich, & K.L. Bildstein. 1996. Decreases in migrating Sharpshinned Hawks (Accipiter striatus) at traditional raptor–migration watch sites in eastern North América. TheAuk, 113:32–40. [ Links ]
Wheeler, B.K. & W.S. Clark. 1995. A photographic guide to North American raptors. Princeton University Press, Princeton, New Jersey. [ Links ]
White, C.M., N.J. Clum, T.J. Cade & W.G. Hunt. 2002. Peregrine Falcon (Falco peregrinus). In A. Poole & F. Gilí (eds). The Birds of North América No. 660. The Birds of North América, Inc, Philadelphia, Pennsylvania. [ Links ]
Zalles, J. & K.L. Bildstein. 2000. Raptor watch: a global directory of raptor migration sites. BirdLife International & Hawk Mountain Sanctuary Association, Londres, Reino Unido. [ Links ]

